PROF. DR WITOLD SYLWANOWICZ, PROF. DR HAB. MED. ALEKSANDER MICHAJLIK I DOC. DR HAB. MED. WITOLD RAMOTOWSKI
ANATOMIA I FIZJOLOGIA CZŁOWIEKA PODRĘCZNIK DLA ŚREDNICH SZKÓŁ MEDYCZNYCH
WARSZAWA 1980
PAŃSTWOWY ZAKŁAD WYDAWNICTW LEKARSKICH
Podręcznik zatwierdzony do użytku średnich szkół medycznych przez Departament Szkolnictwa i Nauki Min. Zdrowia i Opieki Społ. p i s m e m ND-51/M/77 z dnia 3.XII.1977 r.
© C o p y r i g h t by Państwowy Zakład W y d a w n i c t w Lekarskich, 1980 r.
Ryciny w y k o n a l i : Aleksander Dawidziuk i Zenon Projekt okładki: Maksymilian Katużny
Nowacki
Redaktor: m g r Elżbieta Kozłowska-Abgarowicz Redaktor techniczny: Tadeusz Wojdowski, Marian Krupicki Korektor: m g r Zofia Kalinowa
ISBN 83-200-0168-4 PAŃSTWOWY ZAKŁAD WYDAWNICTW LEKARSKICH - WARSZAWA 1980 Wydanie l/VII. Nakład 5000IR225 egz. Objętość ark. wyd. 38,8=34.0 ark druk. Papier offset kl. III. 90 g. 70x100cm. Oddano do składania w kwietniu 1978 r. Podpisano do druku i druk ukończono w styczniu 1980 r. Zam. nr. 2997/K 0-122 ZAKŁADY GRAFICZNE „DOM SŁOWA POLSKIEGO" - WARSZAWA Cena zł 90.-
SPIS TREŚCI
WSTĘP
1
WIADOMOŚCI OGÓLNE
5
Budowa i czynności komórki
5
Podział komórki Rozmnażanie Krótki zarys rozwoju zarodkowego człowieka Organogeneza . . Tkanki . . Tkanka nabłonkowa Tkanka łączna . . Tkanka mięśniowa Tkanka nerwowa Postać człowieka jako całość Okolice ciała Budowa i proporcje ciała w zależności od wieku Zmianyzachodząceworganizmiewciągużycia Symetria i asymetria „ Metameria Proporcje ciała ludzkiego i ich zmiany w ciągu życia Różnice konstytucyjne budowy ciała Przegląd układów narządów Układ narządów ruchu Mięśnie Układ trawienny Układ oddechowy Układ krążenia Serce Układ moczowo-płciowy Układ moczowy Układ płciowy Układ nerwowy Układ gruczołów dokrewnych
8 9 10 13 13 14 16, 19 21 23 24 27 29 31 32 33 34 35 35 38 43 44
45 46
49 49 49 50 51
UKŁAD NARZĄDÓW RUCHU .
53
Układ kostny-osteologia
53
Dane ogólne O kostna Czynność kości Rozwój i wzrost kości Osteologia szczegółowa Kręgosłup Szkielet klatki piersiowej Kości kończyny górnej Kości obręczy kończyny górnej Kości kończyny górnej wolnej Kości kończyny dolnej
i
.
53 55 56 57 59 59 64 65 66 67 72
V
Kości obręczy kończyny dolnej wolnej
73
Kości kończyny dolnej wolnej
75
Kości głowy Dane ogólne Dółczaszkowyprzedni Dół czaszkowy środkowy Dółczaszkowytylny Czaszka trzewna Czaszka noworodka Układwięzadłowy-syndesrrjologia Połączenia kręgów . Czynność kręgosłupa Połączenia żeber. Klatka piersiowa jako całość Połączenia kończyny górnej Staw mostkowo-obojczykowy Staw barkowo-obojczykowy Staw ramienny Staw łokciowy . . Stawpromieniowo-nadgarstkowy Stawy ręki Ręka jako całość Połączenia kończyny dolnej Połączenia obręczy kończyny dolnej Staw krzyżowo-biod rowy Spojenie łonowe Miednica jako całość Połączenia kończyny dolnej wolnej Staw biodrowy Staw kola nowy Połączenia kości goleni Stawskokowo-goleniowy Stawy stopy Stopa jako całość Połączenia kości czaszki StawskronioWo-żuchwowy Układ m i ę ś n i o w y - m i o l o g i a Wstęp. Budowa mięśni Podstawy miomechaniki Miologia szczegółowa Mięśnietułowia Mięśnie klatki piersiowej Przepona Mięśnie brzucha Pochewka mięśnia prostego brzucha Więzadło pachwinowe Czyn ność m ięśni brzucha Mięśnie grzbietu Powięź piersiowo-lędźwiowa Czynność mięśni grzbietu Mięśnie szyi i głowy Mięśnie głowy Mięśnie żwaczowe
VI
80 80 83 84 84 84 87 87 87 89 90 90 91 91 91 92 94 96 96 97 98 98 98
• •
~
98 99 99 99 101 103 104 105 106 107 107 107 107 108 109 110 110 110 112 114 114 115 115 115 119 119 120 122 122
Czynność mięśni żwaczowych
122
Mięśnie mimiczne . Czynność mięśni mimicznych Mięśnie kończyny górnej Mięśnie obręczy kończyny górnej
123 124 124 124
Dół pachowy Czynność mięśni obręczy kończyny górnej Mięśniekończynygórnejwolnej Mięśnie ramienia Mięśnie przedramienia Mięśnie ręki. Pochewki ścięgien ręki Czynnośćmięśnikończynygórnejwolnej Vlięśnie kończyny dolnej Mięśnieobręczykończynydolnej Grupa przednia mięśniobręczykończynydolnej Grupa tylna mięśni obręczy kończyny dolnej (mięśnie pośladkowe) Mięśnie głębokie miednicy Czynnośćmięśniobręczykończynydolnej . Mięśnie kończynydolnej wolnej
124 125 126 126 128 131 134 134 135 135 135 135 136 137 137
Mięśnie uda Grupa przednia mięśni uda Grupa tylna mięśni uda Grupa przyśródkowa mięśni uda Powięzieuda Czynność mięśni uda Mięśnie goleni Grupa przednia mięśni goleni Grupa tylna mięśni goleni Grupa boczna mięśni goleni Powięzie goleni Mięśnie stopy Czynność mięśni goleni i stopy Fizjologia mięśni Dane ogólne Przemiana materii i energii w mięśniach Skurczmięśnia Pobudzenie mięśnia Zmęczenie Zjawiska elektryczne w mięśniu .
UKŁAD TRAWIENNY
137 137 138 139 140 141 141 141 142 144 144 145 147 161 161 162 163 164 166 167
168
Budowa ścian przewodu pokarmowego
170
Jama ustna Podniebienie Zęby Język Błona śluzowa języka Mięśnie języka Gruczoły ślinowe. Ślinianka przyuszna Ślinianka podżuchwowa Ślinianka podjęzykowa Gardło . . .
170 171 172 176 176 177 178 178 178 178
•
179
VII
Część nosowa ga rdła Część ustna ga rdła Część krtaniowa gardła . Budowa ściany gardła Trawieniepokarrńuwjamieustnej Połykanie
179 180
180 180 181 182
Przełyk
183
Żołądek.... Budowa ściany żołądka Trawienie pokarmu wżołądku Sok żołądkowy .•••'• Metody otrzymywania soku żołądkowego Skład soku żołądkowego Regulacja wydzielania soku żołądkowego Wymioty Jelito. Jelito cienkie Budowa jelita cienkiego Gruczoły jelita cienkiego Dwunastnica Jelito cienkie krezkowe Jelitogrube Jelito ślepe i wyrostek robaczkowy Okrężnica Odbytnica Wielkie gruczoły trawienne Wątroba Budowa wątroby Budowa mikroskopowa wątroby Drogi żółciowe i pęcherzyk żółciowy Czynności wątroby Trzustka Budowa wewnętrzna trzustki Czynności trzustki Otrzewna Trawienie p o k a r m ó w w jelicie cienkim Ruchy jelita cienkiego Trawieniechemicznewjeliciecienkim Wchłanianie Trawienie pokarmów w jelicie grubym Ruchy jelita grubego Kał Defekacja Przemiana materii i energii. Odżywianie Przemiana białka Przemiana węglowodanów Regulacja poziomu cukru we krwi Przemiana tłuszczów Przemiana w o d y i soli mineralnych
183 185 186
Solewapnia
VIII
"
.
Solefosforu Sole sodu i potasu Innesole Witaminy
^
187 187 188 189 189 189 189 191 191 192 193 194 195 196 196 196 197 199 200 202 203 203 204 204 205 205 207 210 212 212 213 213 214 215 218 222 222 225 226
•
227 227 227 227
Przemiana energii Metody pomiaru wydatkowania energii Współczynnik oddechowy Oznaczanie wytwarzanej energii na podstawie przemiany gazowej Przemiana podstawowa Przyrost czynnościowy przemiany energii Swoistodynamiczne działanie pokarmu Żywienie Zapotrzebowanie kaloryczne Regulacja temperatury Bilans cieplny ustroju Produkcja ciepła . Straty ciepła Regulacja temperatury ciała
UKŁAD ODDECHOWY . .
230
230 231 232 232 233 234 234 234 238 238 239 239 240
242
Drogi oddechowe Nos i jama nosowa Krtań Chrząstki krtani Więzadła krtani Mięśnie krtani Jama krtani • • • ; Tchawica Oskrzela Płuca Topografia płuc Budowa oskrzeli i płuc Unaczynienie i unerwienie płuc Opłucna Śródpiersie Fizjologia układu oddechowego Oddychanie płucne Mechanika oddychania Czynniki wpływające na ciśnieniewjamieopłucnej Pojemność płuc Krążenie płucne Wymiana gazów Rola krwi w procesie oddychania Oddychanie tkankowe Rola dwutlenku węgla w regulacji równowagi kwasowo-zasadowej Regulacja oddychania
243 243 245 246 247 248
UKŁAD MOCZOWY
271
Budowa nerki Czynność nerek Przepływ krwi przez nerki Przesączanie kłębkowe Pomiar przesączania kłębkowego Reabsorpcja kanalikowa Regulacja reabsorpcji Wydzielanie Regulacja równowagi kwasowo-zasadowej Regulacja gospodarki wodnej
251 252 252 255 256 257 258 259 259 260 260 261 262 263 264 266 267 268 269
272 273 274 275 275 277 278 279 279 281
IX
Mocz . Moczowód Pęcherz moczowy Oddawanie moczu Cewka moczowa
282 283 284 285 285
UKŁAD PŁCIOWY
287
Narządy płciowe męskie.
287
Narządy płciowe męskie wewnętrzne Jądro Tkanka śródmiąższowa i wydzielanie wewnętrzne jądra Najądrze Nasieniowód Gruczoł krokowy Gruczoły opuszkowo-cewkowe . Powrózek nasienny Przewody wytryskowe Narządy płciowe męskie zewnętrzne Prącie. Wzwód prącia Moszna Narządy płciowe żeńskie Narządy płciowe żeńskie wewnętrzne Jajnik Hormony płciowe żeńskie Jajowód Macica Pochwa Zewnętrzne części płciowe żeńskie Wzgórek łonowy Wargi sromowe większe Wargi sromowe mniejsze Przedsionek p o c h w y . Błona dziewicza Łechtaczka Opuszki przedsionka Gruczoły przedsionkowe większe Sutek Akt płciowy, spółkowanie Zapłodnienie. Ciąża. Poród Implantacja Ciąża Poród Mechanizm porodowy. . Termin porodu
UKŁAD KRĄŻENIA
^
X
288 288 290 292 293 293 294 294 294 295 •
296 297
297 298
,
298 298 302 304 305 310
312 312 313 313 314 314 314
5 315 315
317 318 318 320
320 321 323
324
Serce
325
Kształt serca Przedsionek prawy Komora prawa Przedsionek lewy Komora lewa Budowa serca
325 327 327 328 328 328
Worek osierdziowy Położenie serca Unaczynienię i unerwienie serca . . Naczynia krwionośne Tętnice Żyły Naczynia włosowate Naczynia tętnicze krążenia wielkiego . . Tętnica główna (aorta) Gałęzie łuku aorty Tętnica szyjna wspólna. . . Tętnica szyjna zewnętrzna Gałęzie tętnicy szyjnej zewnętrznej Tętnica szyjna wewnętrzna Tętnica podobojczykowa Tętnica pachowa Tętnica ramienna Gałęzie aorty zstępującej. Odgałęzienia aorty piersiowej Gałęzie aorty brzusznej Gałęzie ścienne aorty brzusznej Gałęzietrzewnenieparzysteaortybrzusznej Gałęzie trzewne parzyste aorty brzusznej Gałęzie końcowe aorty brzusznej Tętnica biodrowa wewnętrzna Tętnica biodrowa zewnętrzna Tętnica udowa Tętnica podkolanowa Tętnice goleni i stopy Naczynia żylne krążenia wielkiego .Żyła główna górna Żyła nieparzysta krótka Żyła ramienno-głowowa Żyły głowy i szyi Żyły kończyny górnej Żyły powierzchowne kończyny górnej Żyły głębokie kończyny górnej Żyła główna dolna Dopływy ścienne żyły głównej dolnej Dopływy trzewne żyły głównej doi nej Żyły biodrowe wspólne Żyły kończyny dolnej Układ żyły wrotnej Połączenia żyływrotnej z układami żyły głównej górnej i dolnej Naczynia krążenia małego Krążenie płodowe Układ chłonny Naczynia i węzły chłonne kończyny dolnej Naczynia i węzłychłonne jamy brzusznej i miednicy małej Naczynia i węzły chłonne klatki piersiowej Naczynia i węzły chłonne głowy i szyi Naczynia i węzły chłonne kończyny górnej Śledziona Budowa śledziony
.• . . .
33U 330 331 332 332 332 333 333 333 335 335 336 336 337 338 339 340 342 342 343 343 343 345 345 345 346 347 348 348 350 350 350 350 351 352 352 352 354 354 355 355 356 357 358 358 359 359 361 362 363 364 364 364 365
XI
Czynność śledziony Fizjologia układu krążenia Wiadomości ogólne Czynność serca Uderzenie koniuszkowe i tony serca-. Praca serca Objętość m i n u t o w i Objętość minutowa a opór o b w o d o w y Objętość minutowa a powrótżylny Objętość minutowa a częstość skurczów Objętość minutowa a objętość wyrzutowa (rzutserca) Zjawiska elektryczne w sercu Automatyzm serca Czynność naczyń krwionośnych Krążenie krwi w tętnicach Ciśnienie krwi W p ł y w siły ciężkości na ciśnienie krwi Tętno ' Szybkość prądu krwi Krążenie krwi wtkankach Regulacja przepływu tkankowego krwi Baroreceptory Stężenie C0 2 Stężenie 0 2 Nerwy czuciowe Ośrodkowy układ nerwowy Inne czynniki regulujące objętość łożyska naczyniowego W p ł y w nerwów rozszerzających naczynia Płyn tkankowy i limfa Powstawanie i resorpcja płynu tkankowego Obrzęk Krążenie płucne
KREW
. ... .
366 366 366 367 368 369 372 372 372 374 375 375 377 378 378 378 381 382 382 383 383 383 384 385 385 385 385 386 387 387 388 389
391
Osocze krwi
392
Odczyn krwi Krzepnięcie krwi Substancje przeciwkrzepliwe Limfa Elementy upostaciowane krwi Krwinki czerwone (erytrocyty) Krwinki płytkowe (płytki krwi,trombocyty) Szpik kostny Krwinki białe Limfocyty Monocyty Układ siateczkowo-śródbłonkowy Mechanizmy obronne krwi
395 395 396 397 397 397 399 400 400 401 401 401 401
Antygeny Przeciwciała Mechanizm powstawania przeciwciał ^ Grasica . . Reakcja antygen-przeciwciało Grupy krwi i przetaczanie krwi
402 402 402 403 403 404
XII
UKŁAD NERWOWY
407
Neuron Rdzeń kręgowy Ośrodki rdzenia kręgowego Odruch Opony rdzenia kręgowego Opony mózgowia Opona twarda mózgowia / Pajęczynówkamózgowia . . . . . Opona miękka mózgowia Płyn mózgowo-rdzeniowy Mózgowie ..... Rdzeń przedłużony Tyłomózgowie wtórne Śródmózgowie Międzymózgowie i jądra podkorowe kresómózgowia Kresomózgowie Półkule mózgowe Kora Komory boczne półkul mózgowych Węchomózgowie Ośrodki korowe Ośrodki m o w y Elektroencefalografia Odruchywarunkowe Sen . . Twór siatkowaty Nerwy czaszkowe Nerwy węchowe (I). Nerw wzrokowy (II) Nerw okoruchowy (III) Nerw bloczkowy (IV) Nerw trójdziel ny (V) Nerw odwodzący (VI) Nerw twarzowy (VII) Nerw przedsionkowo-ślimakowy (VIII) Nerw językowo-gardłowy (IX) Nerw błędny (X) Nerw dodatkowy (XI) Nerw podjęzykowy (XII) Nerwy rdzeniowe Gałęzie grzbietowe nerwów rdzeniowych Gałęzie brzuszne nerwów rdzeniowych. Splot szyjny Splot ramienny Nerwy międzyżebrowe Splot lędźwiowy . Splot krzyżowy Splot s r o m o w y . Splot guziczny Układ nerwowy autonomiczny Zwoje autonomiczne Część współczulna układu autonomicznego
407
412 415 415 417 418 — • • ... ... • -
. • •
418 419 419 420 420 424 425 427 427 430 430 430 432 433 433 435 435 436 437 438 439 439 441 442 443 443 444 444 445 446 446 447 448 449 450 450
.
• •
451 451 455 455 456 458 458 458 461 462
XIII
Część przywspółczulna układu autonomicznego Autonomiczne unerwienie narządów i tkanek
463 464
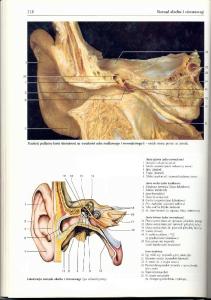
UKŁAD NARZĄDÓW ZMYSŁÓW Narządy czucia powierzchniowego i głębokiego Narząd smaku . . . Narząd powonienia Narząd przedsionkowo-ślimakowy Narząd ślimakowy Ucho zewnętrzne Błona bębenkowa Ucho środkowe Jama bębenkowa Kosteczki słuchowe Trąbka słuchowa Ucho wewnętrzne Ślimak Kanały półkoliste Łagiewka i woreczek Narząd wzroku Budowa ogólna Gałka oczna Błona włóknista gałki ocznej Twardówka Rogówka . . . . Błona naczyniowa gałki ocznej
466 467 469
- •
470 471 471 472 473 473 473 474 475 475 477 478 479 479 481 481 481 482 483
Naczyniówka Ciało rzęskowe . Tęczówka Błona wewnętrzna gałki ocznej Nerwwzrokowy Zawartość gałki ocznej Komora przednia gałki ocznej Komora tylna gałki ocznej Soczewka Ciało szkliste Czynności optyczne gałki ocznej Akomodacja Adaptacja Naczynia i nerwy gałki ocznej Narządy dodatkowe oka Aparat ruchowy gałki ocznej Aparat ochronny gałki ocznej
483 483 484 484 486 486 487 488 488 489 489 491 492 492 493 493 494
Spojówka. . Narząd łzowy Gruczoł łzowy Drogi łzowe. .
495 495 495 496
POWŁOKA WSPÓLNA
497
Budowa ogólna skóry Budowa szczegółowa skóry Twory nabłonkowe skóry ' Gruczoły skóry Gruczoły potowe
497 499 501 501 501
XIV
Gruczoły łojowe Gruczoły sutkowe Włosy Paznokcie
> 503 503
• •
GRUCZOŁY DOKREWNE
504
506
Przysadka Gruczołtarczowy Gruczoły przytarczyczne
512 514
Grasica '•.. Gruczołynadnerczowe . Aparat wysepkowy trzustki Gruczoły płciowe jako gruczoły dokrewne Jajnik Jądro Hormony lokalne
515 516 519 523 523 525 525
507
•
WSTĘP
Już w najwcześniejszych okresach rozwoju ludzkości człowiek stykał się z chorobą. Musiał doznawać skaleczeń czy zranień w życiu codziennym, szczególnie na łowach lub przy ciężkiej pracy fizycznej i szukać pomocy u zajmujących się sztuką leczenia, którzy byli jednocześnie czarodziejami i kapłanami. Ślady wykonywanych przez nich zabiegów, nieraz dość skomplikowanych, znajdują archeolodzy na odkopywanych szczątkach kości ludzkich. Zajmujący się leczeniem musieli przez długi szereg pokoleń zdobywać i pogłębiać znajomość budowy i czynności ciała ludzkiego. Pierwsze uporządkowane wiadomości anatomiczne pochodzą z piątego wieku przed naszą erą, dając początek anatomii, nauce o budowie ciała ludzkiego, najstarszej z podstawowych nauk lekarskich. Fizjologia, nauka o czynnościach organizmu, zaczęła się rozwijać znacznie później i dopiero od wieku XVII rozpoczyna się szybki postęp w tej dziedzinie. Obie wymienione nauki są częściami biologii - nauki o życiu. Należy pamiętać, że budowa i czynności organizmu są ze sobą ściśle powiązane i oddziałują wzajemnie na siebie. Jako przykład można podać budowę ręki człowieka, dzięki której może on wykonywać zarówno ciężką pracą fizyczną, jak i precyzyjne czynności. Natomiast praca znacznie obciążająca stopy, jak dźwiganie ciężarów przez robotników portowych czy taniec w balecie klasycznym, prowadzi do wyraźnego pogrubienia kości śródstopia. Z tego względu omawiamy równolegle budowę i czynności ciała ludzkiego. W ciągu wieków nauka o życiu nagromadziła ogromną liczbę faktów. Z konieczności musimy ograniczyć się do poznania zaledwie jej części, jaką jest anatomia i fizjologia człowieka, w zakresie niezbędnym do opanowania obranego zawodu. Biologia współczesna dzieli się na dwa duże działy: morfologię, czyli naukę o kształtach istot żywych, i ergologię - naukę o ich czynnościach. Należy pamiętać, że jest to jedynie podział umowny, gdyż obie te części są ze sobą ściśle powiązane. Zarówno jedna, jak i druga omawiają świat istot żywych, podzielony na świat roślinny i zwierzęcy. W każdym z nich znajdujemy bardzo wiele form, różniących się od siebie zarówno budową, jak i trybem życia. W długim szeregu form ewolucyjnych zwierząt musimy ograniczyć się do Człowieka, który w gromadzie Naczelnych należy do ssaków, będących kręgowcami stałocieplnymi. W rozwoju wewnątrzmacicznym płód ludzki jest zaopatrzony między innymi w błonę płodową, zwaną owodnią, o czym będzie mowa dalej. Umieszczamy więc człowieka wśród o w o d n i o w c ó w , w przeciwieństwie do bezowodniowców, a więc ssaków, u których owodnia się nie rozwija. 1
Człowiek, jako najwyżej zorganizowany w ś r ó d Naczelnych, jest przedmiotem badań antropotomii, którą zwykle nazywamy anatomią Człowieka, oraz jego fizjologii. Badając budowę człowieka, możemy łatwo zaznajomić się z jego kształtami zewnętrznymi. Ukryte są jednak przed nami jego narządy wewnętrzne, położone w jamach ciała: w jamie czaszki (cavum cranii), w jamie klatki piersiowej (cavum thoracis), w jamie brzusznej (cavum abdominis) i w jamie miednicy (cavum pelvis). Nie dostrzegamy również leżących w głębi ciała naczyń krwionośnych, nerwów, mięśni, jak również szczegółów ich budowy. Do tych ukrytych narządów możemy dostać się podczas sekcji i preparowania zwłok. Metoda ta jest niezbędna do poznania stosunków przestrzennych. Do tego też celu służy również metoda rentgenowska, która daje jednak tylko zarysy narządów, możliwe do zarejestrowania na kliszy fotograficznej. Natomiast przy wprowadzeniu do naczyń (krwionośnych, limfatycznych) lub do przewodu pokarmowego środków cieniujących można tą metodą badać przebieg naczyń, kształty odcinków tego przewodu, jak również obserwować bezpośrednio szereg czynności fizjologicznych. Przytoczone metody nie tłumaczą nam jednak złożonych czynności narządów. Możemy, na przykład, poznać kształty wątroby, stwierdzić wydzielanie żółci. Ale żeby pojąć szczegółowo czynności tego narządu, m u s i m y poznać jego b u d o w ę mikroskopową, korzystając z przyrządów optycznych, wymagających specjalnej metody badań. W związku z t y m dzielimy anatomię na dwa duże działy - anatomię makroskopową i mikroskopową, czyli histologię, naukę o tkankach, z której wydzielono naukę o komórce - cytologię. Anatomia makroskopowa dzieli się z kolei na szereg gałęzi, z których każda omawia b u d o w ę ciała ludzkiego z innego punktu widzenia. Medyk najpierw poznaje anatomię p r a w i d ł o w ą (opisową). Dla potrzeb artystów-plastyków powstaje anatomia artystyczna albo plastyczna, która jest właściwie anatomią powierzchni ciała ludzkiego. Chirurga interesuje położenie i układ wzaj e m n y narządów wewnętrznych oraz drogi dostępu do nich, jak również układ kości, stawów, mięśni, naczyń krwionośnych i n e r w ó w , zwłaszcza położonych na kończynach. Stąd też wzięła początek anatomia topograficzna, korzystająca m.in. z metody przekrojów zarówno całego ciała, jak i jego części. A n a t o m i a kliniczna zwraca szczególną uwagę na b u d o w ę i położenie narządów wewnętrznych, co jest szczególnie ważne dla lekarza ze względów praktycznych. Anatomia rentgenowska (rentgenoanatomia) szczegółowo opisuje kości i stawy oraz narządy wewnętrzne, zawarte w jamach ciała. Szczególne znaczenie należy przypisać stosunkowo n o w e m u kierunkowi nauczania, mianowicie anatomii osobnika żywego. Omawia ona szczegóły b u d o w y ciała, dostępne badaniu bez sekcji jedynie metodami klinicznymi. Należą do nich oglądanie (inspectio), badanie d o t y k i e m ( p a l p a t i o ) , o p u k i w a nie ( percussio ), osłuchiwanie ( auscultatio) oraz w s p o m n i a n e wyżej badanie promieniami Roentgena. Metody te mogą być stosowane bezpośrednio lub 2
przy użyciu szeregu narzędzi, jak słuchawki, wzierniki, narzędzia pomiarowe itp. Fizjologia korzysta również z rozmaitych, nieraz bardzo skomplikowanych, metod i urządzeń do badań czynności osobników zdrowych, jak również do prac laboratoryjnych, klinicznych i doświadczeń na zwierzętach. Jak już wspomniano, badanie skomplikowanych czynności organizmu jest związane z koniecznością uprzedniego dokładnego poznania jego budowy.
WIADOMOŚCI O G Ó L N E
BUDOWA I CZYNNOŚCI KOMÓRKI Najprostsze organizmy żywe składają się z jednej komórki, która spełnia wszystkie ich czynności życiowe. Człowiek należy do istot wielokomórkowych, zbudowanych z bardzo dużej liczby komórek (ok. 30 bilionów) o rozmaitych kształtach i czynnościach. W zasadzie jednak budowa tych komórek jest podobna. Komórka, stanowiąca podstawową jednostkę żywej substancji, składa się ż drobnej masy protoplazmy, zawierającej jądro i otoczonej błoną komórkową. Kształty komórek są bardzo różne. Spotykamy komórki kuliste, cylindryczne, sześcienne, wydłużone, spłaszczone, gwiaździste i in. Od ciała komórki mogą odchodzić różnego rodzaju wypustki, często w postaci włókien pojedynczych lub rozgałęzionych. Wiele komórek po, izolowaniu przyjmuje kształt kulisty. Komórki takie występujące w wielkiej masie mają kształt wielościanów, ściśle stykających się ze sobą.
Ryc. 1. Różne kształty komórek.
Wielkość ciała komórek jest na ogół związana z ich czynnością i waha się w granicach od paru mikrometrów do paru milimetrów. Komórki mięśniowe mogą być znacznie dłuższe. Natomiast długość wypustek jest nieraz bardzo znaczna i może dochodzić do kilkudziesięciu centymetrów, jak to widzimy w niektórych komórkach nerwowych. Protoplazma komórkowa, zwana cytoplazmą, ma bardzo skomplikowaną 5
budpwę, związaną z jej czynnościami. Cytoplazma większości komórek ma postać sieci złożonej z cienkich parzystych błon elementarnych, tworzących tzw. siateczkę śródplazmatyczną, która tworzy granice jam różnego kształtu wąskich kanałów, cewek, okrągłych pęcherzyków itp. Są w niej zawarte tzw. organelle komórkowe, jak centrosom, odgrywające dużą rolę przy podziale komórki; mitochondria, związane z dostarczaniem energii komórce; różne włókienka (mięśniowe, nerwowe); aparat siateczkowy wewnętrzny (Golgiego), który jest związany z produktami wytwarzanymi przez komórkę. Poza nimi znajdujemy w niej inne zmienne wtręty komórkowe w postaci ziarenek czy grudek oraz przestrzenie, zwane wakuolami, wypełnione płynami.
Ryc. 2. Schemat komórki widzianej w mikroskopie elektronowym.
W ciele komórki leży jądro komórkowe, otoczone błoną jądrową, wyraźnie odgraniczające jego zawartość od cytoplazmy. Protoplazmę tworzącą jądro nazywamy karioplazmą. Jądro miewa różne kształty - kulisty, wydłużony, kiełbaskowaty, segmentowany i in. Jest ono niezbędnym składnikiem komórki. Komórki pozbawione jąder, jak np. krwinki czerwone, które tracą jądra w trakcie rozwoju, mogą żyć przez pewien czas, ale nie mogą się rozmnażać. Mówiąc o czynnościach komórki, należy wspomnieć o tym, że w różnych organizmach nie są one jednakowe. Jak wiemy, istnieją organizmy j e d n o i wielokomórkowe. Pierwsze składają się z jednej komórki, która spełnia wszystkie czynności życiowe. U drugich znajdujemy wielką rozmaitość kształtów komórek, przystosowanych do określonych czynności. Dla przykładu wymienimy czynności ruchu, tego najbardziej widocznego przejawu życia. Niektóre organizmy jednokomórkowe, jak np. ameba, poruszają się ruchem pełzakowatym, wypuszczając w pewnym kierunku część swej 6
Ryc. 3. Komórka poruszająca się ruchem pełzakowatym.
protoplazmy w postaci nibynóżek ( pseudopodia ) , za którymi podąża - jak gdyby przelewając się - reszta protoplazmy (p. ryc. 3). U człowieka, jako organizmu wielokomórkowego, istnieją zgrupowane specjalne komórki mięśniowe, które kurcząc się powodują ruch, odbywający się na zupełnie innych zasadach. Jednokomórkowce odbierają bodźce ze świata zewnętrznego przez kontakt z otoczeniem, w którym przebywają i reagują na te bodźce bezpośrednio, zbliżając się lub oddalając od ich źródła i wykazując w ten sposób tropizm dodatni lub ujemny. Człowiek natomiast ma specjalne komórki nerwowe, odbierające bodźce świetlne, termiczne, chemiczne i inne, które przekazują je do ośrodka układu nerwowego, powodując odruchową lub świadomą czynność organizmu. Jednokomórkowce pobierają pożywienie bezpośrednio ze środowiska płynnego, w którym żyją. U wielokomórkowców istnieją całe masy wysoko wyspecjalizowanych komórek, wypełniających różne zadania w bardzo złożonym procesie trawienia. Z powyższych przykładów wynika, że u najniższych istot żywych jedna komórka spełnia zadania całego organizmu. Dlatego też niektórzy proponują nazywać je nie pierwotniakami, lecz organizmami jednokomórkowymi, spełniającymi wszystkie czynności życiowe. Przy dzisiejszym stanie wiedzy rO można podać ścisłej definicji życia. Możemy jednak określić cechy istot żywych, odróżniające je od martwych. Są 7
to przede wszystkim przemiana materii i energii , czyli metabolizm, oraz zdolność rozmnażania. Metabolizm odbywa się w każdej komórce, natomiast jedynie niektóre komórki żywe w organizmie dojrzałym nie wykazują zdolności podziału, jak np. komórki nerwowe czy wspomniane krwinki czerwone człowieka, nie posiadające jąder komórkowych. Mówiliśmy wyżej o najbardziej widocznym przejawie życia, jakim jest ruch. Charakteryzuje on głównie organizmy zwierzęce, lecz nie jest cechą wszystkich komórek. Jest on właściwością pewnych tkanek lub nawet pojedynczych komórek, jak niektóre białe krwinki. Występuje on jedynie w określonych miejscach i w warunkach, podyktowanych przez całość organizmu.
Podział komórki Na skutek przebiegających w komórce skomplikowanych procesów metabolicznych ciało jej powiększa się - komórka wzrasta. Proces ten nie może być nieograniczony i kiedy masa komórki wystarcza na wytworzenie komórki potomnej, następuje podział jej na dwie części. Istnieją dwa rodzaje podziału komórki - bezpośredni, amitotyczny, i pośredni, mitotyczny, czyli mitoza. U organizmów wyższych, zarówno roślinnych, jak i zwierzęcych, występuje zwykle podział pośredni. Mitoza polega na równomiernym podziale materii, przenoszącym cechy dziedziczne na dwie komórki potomne. Cząsteczki materii, przenoszące cechy dziedziczne, jak np. kształt nosa czy małżowiny usznej, barwa oczu, włosów, skóry itp., które mieszczą się w jądrze komórkowym, noszą nazwę genów. Są one zbudowane z cząsteczek kwasu dezoksyrybonukleinowego, oznaczanego skrótem DNA, które mają zdolność samopomnażania się i powiększania w ten sposób ilości materii, przenoszącej cechy dziedziczne. Podział komórki odbywa się w jądrze i cytoplazmie. W jądrze następuje rozluźnienie zbitego kłębka chromatyny, w cytoplazmie tworzy się tzw. aparat podziałowy. Składa się on z dwóch centriol, pochodzących z centrosomu, które rozchodzą się do przeciwległych biegunów komórki, i wrzeciona utworzonego z przebiegających między nimi promieniowań biegunowych. W tym czasie błona jądrowa ulega rozpuszczeniu i zawarta w jądrze chromatyna układa się w poszczególne pętle, zwane chromosomami. Liczba ich u rozmaitych gatunków jest różna, u człowieka wynosi ona 46. Tworzą one 23 pary dobranych do siebie pod względem wielkości i kształtu tworów, zwanych autosomami, i dwa różne chromosomy płciowe. Trzeba zaznaczyć, że w „garniturze chromosomów" występują formy różnego kształtu, ale zawsze tworzące ten sam zespół (ryc. 4). W trakcie podziału komórki wszystkie chromosomy ulegają rozszczepieniu podłużnemu. Każda z połówek chromosomów wędruje do przeciwległego bieguna i w ten sposób każda komórka, powstała z podziału, zachowuje pierwotną liczbę chromosomów. 8
Ryc. 4. Mapa genetyczna przedstawiająca układ niektórych genów w chromosomach. iu>iorem czerwonym oznaczono geny determinujące skład krwi; zielonym - syntezę enzymów biorących udział w przemianie materii; niebieskim - syntezę innych enzymów; żółtym - geny przekazujące pewne choroby dziedziczne; fioletowym - geny wyznaczające różne inne funkcje.
Jednocześnie dzieli się cytoplazma razem z organoidami komórkowymi i dwie powstałe z podziału komórki oddzielają się od siebie. Tak zakończona zostaje mitoza, która trwa około godziny. Do następnej mitozy dochodzi po dłuższej przerwie 10-20 h. W ten sposób rozmnażają się komórki w organizmach istot żywych, powodując wzrastanie organizmu lub zamianę komórek starzejących się czy uszkodzonych.
Rozmnażanie Podaliśmy wyżej zasady podziału różnych komórek organizmu. Występuje jednak w nim również całkiem odmienny, bardzo złożony podział, spotykany w komórkach płciowych, który opiszemy jedynie ogólnie. Wiemy, że w organizmach jednego gatunku znajduje się stała liczba chromosomów. Wiemy też, że organizmy potomne rozwijają się z dwóch łączących się komórek płciowych. Otóż gdyby po zapłodnieniu komórki jajowej przez wniknięcie do niej plemnika nastąpił zwykły podział mitotyczny tej podwójnej komórki, to w jej ciele znalazłaby się podwójna liczba chromosomów, prowadząca do powstania komórek potomnych o podwójnym garniturze chromosomów. Byłyby to komórki niezdolne do życia.
Zapobiega temu podział redukcyjny komórek płciowych, czyli mejoza. Jej zadaniem jest, jak wspomnieliśmy, zapewnienie zapłodnionej komórce jajowej właściwej liczby chromosomów, a z drugiej strony - wprowadzenie do masy zarodkowej genów z chromosomów zarówno męskiej, jak i żeńskiej komórki płciowej. Komórki płciowe u obu płci, zwane gametami, przechodzą w ciągu rozwoju i dojrzewania skomplikowane przemiany, prowadzące do zmniejszenia liczby chromosomów o połowę. Proces ten nosi nazwę redukcji chromosomów. Komórka jajowa jeszcze niedojrzała, zwana oocytem I rzędu, dokonuje dwóch podziałów, w wyniku których zostają przez nią wytworzone dwa ciałka kierunkowe, zabierające z niej połowę garnituru chromosomów, zwanych autosomami, oraz jeden chromosom płciowy X. Podobne przemiany zachodzą w niedojrzałej komórce płciowej męskiej, zwanej spermatocytem I rzędu. Dzieli się on na dwie komórki, zwane spermatocytami II rzędu, z których każdy zabiera połowę autosomów i jeden chromosom płciowy. Ponieważ w spermatocycie I rzędu istnieją dwa różne chromosomy p ł c i o w e - X i Y, to powstające z jego podziału dwa spermatocyty II rzędu mają garnitury chromosomów różniące się od siebie - jeden ma chromosom X, determinujący płeć żeńską, a drugi Y. Mejoza odbywa się w trakcie dojrzewania gamet w ten sposób, że chromosomy homologiczne (determinujące te same cechy) układają się parami. W każdej takiej parze znajdują się ściśle przylegające do siebie chromosomy podobnego kształtu i wielkości, z których jeden pochodzi z jaja, a drugi z plemnika. Proces ten, zwany koniugacją chromosomów, prowadzi do wymiany niektórych odcinków chromosomów, zawierających określone geny. Powstaje w ten sposób nowa całkowicie przypadkowa kombinacja genów ojca i matki, prowadząca do powstania nowych cech dziedzicznych. Dalsze podziały blastomerów odbywają się na drodze mitozy.
KRÓTKI ZARYS ROZWOJU ZARODKOWEGO CZŁOWIEKA Na podstawie badań paleontologicznych wiemy, że życie nie pojawiło się w postaci takich i tylu form, jak obecnie. Rozwijało się ono przez długi okres, kiedy na ziemi powstały warunki do jego rozwoju, a więc kiedy kula ziemska oziębiła się, utworzyła się atmosfera, pojawiła się woda. Pierwsze formy życia musiały powstać w wodzie, gdyż reakcje chemiczne, na których opiera się życie w tej postaci, jaką znamy, mogą zachodzić jedynie w środowisku wodnym. Znamy już ogólne drogi rozwoju organizmów zwierzęcych. Wśród zwierząt wodnych znalazły się takie, które mogły żyć w wodzie i na lądzie. Potem formy lądowe zaczęły się różnicować, przechodząc stopniowo do form wyższych i wreszcie Naczelnych, wśród których najwyższy stopień osiągnął Człowiek, dzięki najwyżej rozwiniętemu układowi nerwowemu, a szczególnie mózgowi. 10
Ten rozwój świata zwierzęcego nazwano rozwojem filogenetycznym lub filogenezą. Otóż badając rozwój osobniczy zarodka zwierząt wyższych i człowieka, czyli rozwój ontogenetyczny, embriolodzy zauważyli, że w ontogenezie człowiek na ogół przechodzi kolejne stadia filogenezy. Na początku zarodek ma postać zapłodnionej komórki jajowej, z której tworzą się inne formy embrionalne (zarodkowe) podobne do organizmów najprostszych, w niczym nie przypominających człowieka. Następnie formy embrionalne, złożone z wielu komórek, stają się w bardzo ogólnym stopniu podobne do zarodków zwierzęcych, posiadających kręgosłup, głowę, tułów i cztery kończyny, żeby wreszcie zyskać formy noworodka, jeszcze jednak odmienne od człowieka dorosłego, zwłaszcza pod względem proporcji poszczególnych części ciała. Wśród organizmów zwierzęcych napotykamy najprostsze formy, wykształcające się przez podział zapłodnionych komórek płciowych, z których powstaje skupienie w kształcie grudki, noszące nazwę moruli. Jest to stadium rozwojowe komórki, po podziale jej na dwa blastomery pierwsze. Dalsze podziały na blastomery drugie (4), blastomery trzecie (8), czwarte (16) itd. prowadzą do powstania moruli.
•Kjflk
V)
m
Ryc. 5. Początkowe stadia zapłodnionej komórki jajowej: A - k o m ó r k a jajowa, S - d w a blastomery pierwsze, C - blastomery drugie, D - blastomery trzecie.
Dopóki morula nie jest zbyt duża, komórki jej mogą czerpać pożywienie z otaczającego ją płynu. Jednak przy powiększaniu się ich liczby muszą się one układać na powierzchni rosnącego tworu, żeby móc czerpać pożywienie z otoczenia. W związku z t y m wewnątrz moruli tworzy się jama wypełniona płynem, a cały zarodek przedstawia jak gdyby kulę podobną do piłki. Ta forma nosi nazwę blastuli (ryc. 6). Dzielące się komórki na jednym z biegunów są wyższe, na drugim niższe. W dalszym ciągu u zarodka lancetnika następuje stadium gastrulacji. Biegun blastuli z wyższymi komórkami rośnie wolniej, natomiast przeciwległy, szybszy w rozwoju, zaczyna obrastać cały rozwijający się twór. W wyniku tego następuje wpuklenie się bieguna z wyższymi komórkami do przeciwległej ściany blastuli, utworzonej z niższych komórek, na podobieństwo uciśniętej pięścią piłki. W ten sposób powstaje twór, zwany gastrulą, który można porównać do worka o podwójnych ścianach, z nieco zwężonym wejściem. Między warstwami tego worka wytwarza sę nieco tkanki. Warstwy komórek noszą nazwę listków zarodkowych. Warstwa zewnętrzna, z której powstaje 11
Blastula
Gastrula
Ryc. 6. Rozwój zapłodnionego jaja w stadium blastuli i gastruli.
powłoka zewnętrzna i układ nerwowy, nosi nazwę ektodermy (derma, grec. skóra). Wewnętrzna - entoderma, tworzy cewę jelitową. Wreszcie z listka zarodkowego środkowego - mezodermy, powstają kości, mięśnie i niektóre inne narządy. Jak w s p o m n i a n o , niektóre organizmy zatrzymują się w s w y m rozwoju na stadium gastruli. W podobny sposób jest zbudowane ciało mieszkającej w naszych wodach stułbi. Z całych kolonii podobnych t w o r ó w są utworzone gąbki i inne f o r m y , zwane jamochłonami. U tych zwierząt pokarm jest pobierany przez otwór gębowy do wnętrza j a m y ciała, gdzie ulega on trawieniu. Produkty trawienia są wchłaniane przez warstwę entodermalną, a nie strawione resztki są wyrzucane przez ten sam o t w ó r na zewnątrz. U zarodka ludzkiego proces gastrulacji przebiega w sposób nieco odmienny, lecz w zasadzie podobny. W dalszym rozwoju zarodka następuje jego wydłużenie się przez tworzenie się wzdłuż osi ciała segmentów, czyli podobnych do siebie odcinków, zwanych też metamerami. Ślady tego podziału pozostają w organizmie rozwiniętym, n p . w postaci kręgów, żeber, mięśni, naczyń i n e r w ó w międzyżebrowych oraz w budowie niektórych innych narządów. Natomiast niektóre części ciała całkowicie zatraciły b u d o w ę metameryczną, jak np. głowa czy kończyny. Jak w s p o m n i a n o wyżej, komórka jajowa człowieka zawiera bardzo małą ilość materii, z której może powstać jedynie zarodek we wczesnych stadiach rozwojowych. Stąd też wszystkie potrzebne dla jego rozwoju materiały zarodek, rozwijający się w macicy, czerpie z organizmu matki drogą krwi, dochodzącej do narządu, zwanego łożyskiem, które rozwija się w jamie macicy ciężarnej i zostaje wydalone po porodzie. Naczynia krwionośne matki nie łączą się z naczyniami płodu, który ma własne krążenie krwi, odbywające się między płodem i łożyskiem. Wymiana ciał potrzebnych do życia i rozwoju płodu oraz wydalanie produktów przemiany materii płodu odbywa się w łożysku drogą przenikania (osmozy). Płuca płodu są nieczynne, a krew jego natlenia się również w łożysku. 12
Organogeneza Jak w s p o m n i a n o , w stadium gastruli zarodek składa się z trzech listków zarodkowych - ektodermy, meżodermy i entodermy. W dalszym rozwoju z w y m i e n i o n y c h listków kształtują się narządy ciała. Najczęściej jednak powstają one nie z jednego listka zarodkowego, lecz z ich kombinacji. M ó w i l i ś m y , że z ektodermy powstaje p o w ł o k a wspólna ciała (integumentum commune) i układ n e r w o w y . Otóż jedynie bardzo powierzchowna jej warstewka, zwana naskórkiem, oraz paznokcie, włosy i gruczoły skóry powstają z ektodermy. Warstwa głębsza - skóra właściwa - powstaje z meżodermy, jak również leżące jeszcze głębiej mięśnie szkieletowe i kości. Z tego też listka powstają naczynia krwionośne i krew. Przewód p o k a r m o w y powstaje z entodermy, tak jak i płuca oraz wszystkie gruczoły przewodu p o k a r m o w e g o gruczoły jelitowe, wątroba i trzustka. Jednak znaczna część ścian przewodu pokarmowego powstaje z meżodermy. Pomimo zespalania się i mieszania tkanek w rozwoju zarodka wspólne pochodzenie z jednego listka zarodkowego odgrywa niekiedy rolę w patologii. Dla przykładu zaznaczymy, że miejsca bolesnych w y p r y s k ó w na skórze, występujących przy półpaścu (herpes zoster), odpowiadają na ogół przebiegowi n e r w ó w rdzeniowych lub też niektórych czaszkowych pochodzących z ektodermy, podobnie jak nabłonek skóry. Rozwój zarodka odbywa się szybko. Już w drugim miesiącu życia płodowego formują się ogólnie kształty głowy i twarzy, zawiązuje się stosunkowo bardzo duże serce, pojawiają się zawiązki kończyn w postaci parzystych guzków, z których górne układają się bliżej głowy, dolne zaś bliżej zawiązka ogonowej części kręgosłupa. W t y m czasie zarodek ma długość ok. 12 m m . Zarodek dwumiesięczny ma już kształty ludzkie, jednak o całkiem zmienionych proporcjach. Wybitnie duża jest głowa i bardzo małe kończyny. W t y m okresie m ó w i m y już o płodzie (fóetus), który w ciągu dziesięciu miesięcy księżycowych od chwili zapłodnienia dojrzewa i jest przygotowany do życia poza organizmem matczynym. W t y m czasie wykształcają się na rządy wewnętrzne, zmieniają się proporcje, które jednak przy końcu ciąży jeszcze bardzo różnią się od proporcji dziecka, a jeszcze bardziej człowieka dorosłego. Po dojrzeniu płodu następuje poród (partus). Noworodek zaczyna oddychać i krążenie krwi między płodem i łożyskiem drogą naczyń pępkowych zostaje przerwane. Noworodek sam więc zdobywa tlen, a pożywienie czerpie z mleka matki, które trawi za pomocą własnego przewodu pokarmowego, a nie strawione resztki pożywienia wydala na zewnątrz. TKANKI Występujące w organizmie ludzkim narządy mają różne, nieraz bardzo skomplikowane zadania, wymagające udziału licznych komórek o podobnej budowie. Takie skupienia komórek o podobnej budowie i czynnościach noszą nazwę tkanek. Rozróżniamy cztery podstawowe rodzaje tkanek. Są to: 13
1) tkanka 2) tkanka 3) tkanka 4) tkanka
nabłonkowa, łączna, mięśniowa, nerwowa.
Każda z nich ma inną budowę i zadania, jednak żadna nie może samodzielnie pełnić określonej czynności, co można stwierdzić w sztucznej hodowli tkanek. Dopiero różne tkanki, połączone w określony sposób mogą tworzyć narządy, czyli części organizmu, przeznaczone do wykonywania określonych czynności. Tak więc narządem nazwiemy serce, zbudowane głównie z tkanki mięśniowej; żołądek, zbudowany z tkanki nabłonkowej i mięśniowej; oko, zbudowane ze wszystkich rodzajów tkanek itp. Każdy narząd jest zaopatrzony w odżywiające go naczynie krwionośne oraz nerwy, kierujące jego pracą. Jednak do wykonywania złożonych czynności życiowych, jak np. odżywianie, nie wystarczy jeden narząd. Potrzeba tu narządów przyjmujących i rozdrabniających pożywienie, jak zęby i język; trawiących je, jak żołądek i jelita, za pomocą soków trawiennych wytwarzanych przez gruczoły trawienne. Dla przykładu w y m i e n i m y ślinę, sok żołądkowy, sok jelitowy, sok trzustkowy, żółć i in. Szereg narządów, wykonującyh złożoną czynność organizmu, nazywamy układem (systema, apparatus). M a m y w i ę c układ narządów ruchu ( systema organorum motus), układ oddechowy ( apparatus respiratorius s. systema respiratorium) itd. Podział tkanek przeprowadzamy na podstawie ich budowy i czynności.
Tkanka nabłonkowa Tkanka nabłonkowa składa się z komórek ściśle do siebie przylegających i nie wytwarzających istoty międzykomórkowej. Tworzy ona warstwy, pokrywające powierzchnie skóry, narządów wewnętrznych lub wyściełające ściany jam ciała. Komórki nabłonka mają duże, pęcherzykowate jądra, niezbyt silnie się barwiące. Zależnie od kształtów komórek odróżniamy nabłonek płaski, sześcienny (kostkowy) i wałeczkowaty (cylindryczny) (ryc. 7). Kształty nabłonka określamy według kształtów komórek warstwy powierzchniowej, nato.miast warstwa podstawowa składa się zawsze z komórek wałeczkowatych, które rozmnażając się powodują przechodzenie komórek do warstw powierzchownych, stąd starzejące się komórki ulegają złuszczeniu. Nabłonek płaski składa się z komórek, w których jeden w y m i a r (grubość) jest znacznie mniejszy od dwóch pozostałych, ściśle przylegających do siebie. Nabłonek płaski w i e l o w a r s t w o w y pokrywa skórę i najbardziej powierzchowne jego warstwy ulegają zrogowaceniu. Dzięki t e m u skóra ochrania głębsze w a r s t w y ciała przed wysychaniem, promieniowaniem pozafiołkowym oraz przenikaniem do organizmu trucizn i bakterii. 14
Ryc. 7. Różne rodzaje nabłonka.
Nabłonek kostkowy na powierzchni ma komórki kształtu zbliżonego do sześcianu. Wyścieła on głównie drogi moczowe. Nabłonek cylindryczny wyścieła przewód pokarmowy i drogi oddechowe. Może on mieć na powierzchni rzęski, które poruszają się w jednym kierunku; nazywamy go wówczas nabłonkiem m i g a w k o w y m . Odgrywa on dużą rolę w drogach oddechowych, usuwając z płuc drobne cząsteczki kurzu czy węgla, pochodzące z dymu. Nabłonek jest tkanką łatwo regenerującą się dzięki szybkiemu rozmnażaniu się komórek. Komórki nabłonka spełniają szereg ważnych dla całego organizmu czynności. Poza wymienioną czynnością ochronną niektóre komórki nabłonkowe mają zdolność wchłaniania pewnych ciał na drodze osmozy albo też wydzielania lub wydalania określonych produktów przemiany materii. Wspomniany nabłonek cylindryczny, wyściełający przewód jelitowy, ma zdolność wchłaniania produktów trawienia, które następnie wydziela do dróg chłonnych, z których te produkty trafiają do krwi. Inny rodzaj komórek nabłonkowych ma zdolność wydzielniczą. W błonie śluzowej jelita, zwłaszcza grubego, między komórkami cylindrycznymi są 15
rozrzucone komórki kubkowe, wydzielające śluz. Często komórki wydzielnicze zbierają się w większe skupienia, tworząc gruczoły. Gruczoły te, mające kształt pęcherzyków lub ceweczek, mogą występować pojedynczo, jako gruczoły pęcherzykowe lub cewkowe proste. Jeżeli tworzą one skupienie ceweczek lub pęcherzyków, powstają gruczoły pojedyncze rozgałęzione, natomiast grupa takich gruczołów, mających wspólny przewód wyprowadzający, tworzy gruczoł złożony rozgałęziony (ryc. 8). W ten sposób są np. zbudowane niektóre gruczoły trawienne, gruczoły sutkowe i inne. aa
Gruczoł cewkowy prosty
Gruczoł pęcherzykowy prosty
Gruczoł cewkowy zwinięty
Gruczoł pęcherzykowy gronkowy
Gruczoł pęcherzykowy rozgałęziony
Gruczoł cewkowy rozgałęziony
Gruczoł pęcherzykowy złożony
Ryc. 8. Różne rodzaje gruczołów zewnątrzwydzielniczych.
Komórki wydzielnicze tworzą czasami skupienia, pozbawione przewodów wyprowadzających. Nazywamy je gruczołami wydzielania wewnętrznego albo gruczołami dokrewnymi. Nazwa ta pochodzi stąd, że gruczoły takie są otoczone gęsto siecią naczyń krwionośnych, wchłaniających ich wydzielinę wprost do krwi. Należą do nich: gruczoł tarczowy, przysadka, nadnercze i inne.
Tkanka łączna W przeciwieństwie do tkanki nabłonkowej tkanka łączna charakteryzuje się wytwarzaniem istoty międzykomórkowej, w której przebiegają liczne włókna. Kształt komórek bywa bardzo różny i jest powiązany z istotą międzykomórkową. Tkanka łączna występuje w kilku postaciach, a mianowicie: 16
Ryc. 9. Różne rodzaje tkanki łącznej.
1. Tkanka łączna właściwa: a) wiotka, b) zbita, 2. Tkanka łączna tłuszczowa, 3. Tkanka łączna chrzęstno-szklista, 4. Tkanka łączna włóknista, 5. Tkanka łączna sprężysta, 6. Tkanka łączna kostna. Istnieje jeszcze kilka innych rodzajów rzadziej występujących. 2 - Anatomia i fizjologia
Odmianą tkanki łącznej jest również krew, w której komórki są zawieszone w płynnej istocie międzykomórkowej. Dokładny opis krwi znajduje się w odrębnym rozdziale. Tkanka łączna właściwa wiotka składa się z komórek gwiaździstych i licznych włókien. Jedne z nich są miękkie, wiotkie i tworzą grube, falisto przebiegające pęczki. Są to włókna klejodajne. Drugie - włókna sprężyste, są znacznie cieńsze i przebiegają pojedynczo tworząc załamania. Tkanka ta tworzy pajęczynowate sieci lub nieco grubsze warstwy otaczające i łączące narządy, np. skórę z leżącymi głębiej narządami, dzięki czemu skóra może się łatwo przesuwać nad leżącymi pod nią mięśniami lub kośćmi. Natomiast tkanka zbita tworzy pęczki silnych włókien sprężystych, występujących w organizmie w postaci ścięgien lub więzadeł. Tkanka tłuszczowa składa się z kulistych komórek, których ciało poza niewielką ilością plazmy jest wypełnione płynnym tłuszczem, stanowiącym materiał zapasowy. Komórki chrzęstne mają kształt owalny i leżą w torebkach otoczonych istotą międzykomórkową. W chrząstce szklistej istota ta nie wykazuje wyraźnej budowy - stąd jej nazwa. Natomiast w chrząstce włóknistej występuje dużo grubych pęczków włókien klejodajnych przebiegających równolegle między rzędami komórek. W chrząstce sprężystej występują liczne włókna sprężyste, silnie pokrzyżowanemiędzy sobą i tworzące nieregularną siateczkę. Chrząstka szklista występuje np. na końcach kości, tworzących stawy. Chrząstka włóknista tworzy m.in. krążki (chrząstki) międzykręgowe, a sprężysta występuje np. w małżowinie usznej. Chrząstka jest tkanką dość odporną, tworzącą w pewnym okresie życia płodowego szkielet rozwijającego się płodu. Jeszcze bardziej odporna i wytrzymała jest następna odmiana tkanki łącznej - tkanka kostna. Tworzy się ona na podstawie tkanki chrzęstnej, która ulega kostnieniu przez osadzanie się w niej soli węglanów i fosforanów wapnia. Kostnienie odbywa się stopniowo, rozpoczynając od wczesnych okresów życia płodowego i kończy się w wieku od kilkunastu do dwudziestu paru lat. Tkanka kostna występuje pod dwiema postaciami - jako istota zbita oraz istota gąbczasta. Pierwsza występuje w trzonach kości długich, druga zaś w końcach tych kości oraz w kościach różnokształtnych. Istota zbita składa się z jednostek architektonicznyph, zwanych osteonami. Osteon jest tworem cylindrycznym, powstającym dokoła kanaliku, w którym przebiega drobna tętniczka, otoczona blaszką kostną. Na zewnętrznej powierzchni tej blaszki układają się koncentryczne łańcuchy komórek kostnych w tzw. jamkach kostnych, połączonych drobniutkimi kanalikami, łączącymi jam ki kostne w jedną sieć. W tych kanalikach leżą drobne wypustki komórek kostnych, powiązanych w ten sposób ze sobą. Komórki te są otoczone następną blaszką kostną, łańcuchem komórkowym itd. Należy zaznaczyć, że w tych koncentrycznych blaszkach kostnych przebiegają skośne włókna tkanki łącznej, krzyżujące się ze sobą. Układ taki wzmacnia znacznie całą kość. Jak wspomniano, kość powstaje na podstawie łącznotkankowej lub chrzęstnej. Sole wapnia stanowią dwie trzecie części kości, natomiast resztę tworzy 18
tzw. chrząstka kostna, czyli oseina. Dzięki solom mineralnym kość nabiera twardości i wytrzymałości, natomiast oseina nadaje jej pewien stopień elastyczności. Dzięki obu składnikom kość staje się doskonałym materiałem do budowy szkieletu, jako narządu podpory ciała. Podany stosunek między składnikami organicznymi i mineralnymi występuje u osobników dorosłych. U noworodka i dziecka oseiny jest stosunkowo więcej, kości są bardziej elastyczne i rzadziej ulegają złamaniom niż u osobników dorosłych. Natomiast w wieku starczym zwiększa się stosunkowo ilość soli mineralnych, a zmniejsza się zawartość oseiny - kości stają się bardziej kruche i stąd razem z wiekiem rośnie liczba złamań kości. Tkanka siateczkowata składa się z niezbyt licznych komórek gwiaździstych, wytwarzających gęstą siateczkę o drobnych oczkach, w których leżą liczne limfocyty (p. Krew). Z tej tkanki są zbudowane liczne gruczoły chłonne, występujące w układzie krążenia.
Tkanka mięśniowa Zasadniczą właściwością tkanki mięśniowej jest kurczliwość. Tkanka mięśniowa występuje w dwóch postaciach. Bardziej pierwotna i prostsza w budowie tkanka mięśniowa gładka składa się z wrzecionowatych komórek długości kilkunastu do paruset mikrometrów i występuje w postaci błon w ścianach przewodu pokarmowego, w drobnych rozgałęzieniach oskrzeli, w przewodach gruczołów, w ścianach naczyń, w skórze i szeregu narządów wewnętrznych. Komórki mają w zgrubiałej części środkowej pałeczkowate jądro i zaostrzają się ku końcom. Układają się one obok siebie w błony, przebiegające zwykle w jednym kierunku. W protoplazmie komórek mięśniowych, zwanej sarkoplazmą, znajdują się włókienka kurczliwe, które na obwodzie komórki tworzą wiązki zwane miofibrylami. Pod wpływem rozmaitych bodźców włókienka się kurczą, powodując zmianę kształtów komórki - skraca się ona i grubieje. W ten sposób błony mięśniowe założone w ścianach przewodów, naczyń krwionośnych lub narządów mogą podnosić ciśnienie w ich wnętrzu i powodować przemieszczanie się ich treści, np. przechodzenie treści żołądka i jelit przez cały przewód pokarmowy. Do bodźców, powodujących skurcze mięśniówki gładkiej należą przede wszystkim bodźce nerwowe, niezależne od naszej woli. Bodźce te wychodzą z ośrodka układu nerwowego na drodze odruchów, wywołanych również stanami psychicznymi. Na przykład pod wpływem strachu kurczą się drobne mięśnie owłosionej części głowy, powodując podnoszenie się włosów mówimy, że włosy się „jeżą". Chłód może wywołać „gęsią skórkę", która również powstaje przy skurczu mięśni przywłosowych, o czym będzie jeszcze mowa w rozdziale o skórze. Tkanka mięśniowa poprzecznie prążkowana ma budowę znacznie bardziej 19
skomplikowaną. Jej nazwa pochodzi od jaśniejszych i ciemniejszych prążków poprzecznych, widocznych pod mikroskopem przy oglądaniu włókna mięśniowego. Włókna mięśniowe poprzecznie prążkowane są tworami o wielu jądrach, otoczonymi cienką błonką, zwaną sarkolemmą. Włókna te mają stosunkowo duże wymiary - grubość ich wynosi 10-100 ^m, zaś długość od jednego milimetra do kilkunastu centymetrów. Włókno jest wypełnione włókienkami kurczliwymi - miofibrylami. Włókna mięśniowe występują w dużych skupieniach, tworząc narządy, zwane mięśniami. Dzięki znacznej długości włókien mięśnie poprzecznie prążkowane, zwane również szkieletowymi, przyczepione do kości mogą wykonywać rozległe ruchy różnych części ciała. Mięśnie poprzecznie prążkowane, podobnie jak mięśnie gładkie, kurczą się odruchowo pod wpływem różnych bodźców - mechanicznych, chemicznych, elektrycznych, które odbywają się bez udziału naszej woli. Nie mogąc panować nad ruchami, możemy jednak dowolnie kurczyć pewne grupy mięśni dla wykonania określonych zamierzonych ruchów. M ó w i m y więc, że są one zależne od naszej woli.
Ryc. 10. Rodzaje tkanki mięśniowej: A-mięsień gładki, B-mięsień poprzecznie prażkowany, Cmięsień sercowy.
20
Odmienny rodzaj tkanki poprzecznie prążkowanej stanowi tkanka mięśniowa serca. Włókna mięśniowe są również prążkowane, lecz tworzą odgałęzienia, łączące się z sąsiednimi włóknami. Poza tym włókna są podzielone na odcinki długości kilkudziesięciu mikrometrów przegrodą, zwaną wstawką, dobrze widoczną pod mikroskopem. Tkanka mięśniowa serca - w przeciwieństwie do innych mięśni poprzecznie prążkowanych, kurczy się całkowicieniezależnie od naszej woli. Tkanka mięśniowa poprzecznie prążkowana ma jedynie niewielką zdolność regeneracji.
Tkanka nerwowa Tkanka nerwowa jest najwyżej zorganizowaną tkanką człowieka. Jest ona szczególnie wrażliwa na brak tlenu, którego zużywa więcej od innych tkanek i bez którego szybko ginie. Komórki nerwowe nie rozmnażają się po urodzeniu i człowiek przynosi na świat stały komplet tych komórek, których liczba wynosi ok. 30 miliardów. Komórki nerwowe mają zdolność wytwarzania stanów pobudzenia, które mogą być wysyłane do innych komórek. Stany te są przewodzone drogą wytworzonej przez komórkę wypustki, zwanej wypustką osiową, aksonem lub neurytęm. Neuryt odchodzi od komórki kształtu gruszkowatego w jej zwężonym końcu, zwanym biegunem. Nie oddaje on większych odgałęzień i dopiero w końcu rozpada się na cienkie gałązki.
Opisana komórka nerwowa nosi nazwę jednobiegunowej. Istnieją też wydłużone komórki dwubiegunowe, wytwarzające drugą wypustkę rozgałęzioną, zwaną dendrytem. Drogą tej wypustki dochodzą dó komórki bodźce z obwodu. Dendrytów może być znacznie więcej i wtedy komórka riosi nazwę wielobiegunowej*. W niektórych miejscach, np. w zwojach rdzeniowych, spotykamy komórki, w których neuryt i dendryt łączą się bezpośrednio po odejściu od komórki i biegną razem tworząc wrażenie komórki jednobiegunowej. Po krótkim przebiegu neuryt i dendryt rozdzielają się i biegną w przeciwnych kierunkach. Tego rodzaju komórki nazywamy pozornie jednobiegunowymi. Ciało komórki ze wszystkimi wypustkami nosi nazwę neuronu. Komórki nerwowe, zależnie od ich czynności, dzielimy na czuciowe i ruchowe. Długość wypustek komórki nerwowej bywa bardzo różna, gdyż zależy ona od czynności komórki. Ponieważ bodźce biegną między zakończeniami wypustek na obwodzie a komórką, droga ich może być bardzo długa. Na przykład dendryt komórki nerwowej czuciowej, leżącej w zwoju kręgowym przy kręgosłupie, może dochodzić do ciałka czuciowego, umiejscowionego w skórze opuszki palca; neuryt komórki ruchowej, leżącej w rdzeniu kręgowym w kręgosłupie może zaopatrywać komórki mięśnia kończyny dolnej, leżącego w stopie. W t y c h wypadkach wypustki mają długość kilkudziesięciu lubwięcej centymetrów. Wiemy, że komórki nerwowe nie rozmnażają się, mogą jednak one odbudowywać niektóre swe części. Dotyczy to też wypustek, z których tworzą się nerwy. Uszkodzony nerw może wrócić do swych czynności po pewnym, zwykle dłuższym, okresie czasu. Przewodzenie bodźców odbywa się drogą wypustek, które prowadzą liczne włókienka nerwowe, zwane neurofibrylami. Neurofibryle występują również w protopłazmie komórki nerwowej, czyli neuroplazmie, tworząc siateczkę otaczającą jądro komórkowe. W neuroplazmie występują również liczne ziarenka substancji zasadochłonnej, zwanej tigroidem, z której składają się tzw. ciałka Nissla, odgrywające rolę w przemianie materii komórki. Z tkanki nerwowej zbudowane jest mózgowie i rdzeń kręgowy. Wypustki komórek - poza ośrodkowym układem nerwowym - wchodzą w skład nerwów. Tkanka nerwowa jest zbudowana na zrębie, utworzonym z różnokształtnych komórek rozgałęzionych, powiązanych ze sobą i z komórkami nerwowymi. Komórki te tworzą glej, stanowiący rusztowanie podtrzymujące tkankę nerwową, a z drugiej strony pośredniczą w jej odżywianiu. Komórki glejowe pochodzą z zewnętrznego listka zarodkowego, podobnie jak i komórki nerwowe. * Wielkość takich komórek bywa czasem znaczna. Spotykamy je np. w rogach przednich rdzenia kręgowego i na cienkich preparatach po ich zabarwieniu możemy dojrzeć te komórki gołym okiem. Komórki wielobiegunowe mają kształt gwiazdowaty.
22
POSTAĆ CZŁOWIEKA JAKO CAŁOŚĆ Omawiając postać człowieka należy zaznajomić się z szeregiem określeń anatomicznych, dotyczących linii, zaznaczanych na skórze, służących do określania rzutów narządów wewnętrznych na powierzchnię ciała. Również dla opisu stosunków topograficznych niezbędne jest poznanie płaszczyzn ograniczających przestrzenie, w których układają się narządy wewnętrzne, oraz kierunków wyznaczanych w t y c h płaszczyznach. Znającte pojęcia, będziemy moglj przedstawić okolice ciała ludzkiego, niezbędne do umiejscowienia zmian czy omawianych szczegółów na powierzchni ciała. Dalej zwrócimy uwagę na zmiany budowy i proporcji ciała, zachodzące wraz z wiekiem w ciągu całego życia. Przystępując do opisu ciała umawiamy się, że przedstawiamy je w pozycji
ssm w§m
JIIf ., Ryc. 12. Anatomiczna pozycja ciała z zaznaczeniem płaszczyzn orientujących.
23
pionowej, zwrócone do nas przodem, z rękami opuszczonymi, dłońmi skierowanymi do przodu. Jak wiadomo, ciało ludzkie wykazuje na ogół symetrię dwustronną, o której szczegółach będzie mowa dalej. Linia pośrodkowa przednia, biegnąca przez środek czoła, nosa, bródki, szyi i tułowia, stanowi granicę między prawą i lewą połową ciała. Po stronie tylnej, od szczytu głowy wzdłuż kręgosłupa aż do jego końca - kości guzicznej, biegnie linia pośrodkowa tylna. Jedna i druga stanowią osie pionowe ciała. Jeżeli te dwie linie połączymy linią prostą, poziomą, to otrzymamy oś strzałkową, której nazwa pochodzi od kierunku strzały. Jak wiemy, dwie przecinające się linie proste określają płaszczyznę. Ta więc płaszczyzna, dzieląca ciało na symetryczne połowy, nosi nazwę płaszczyzny pośrodkowej i jest jedną z płaszczyzn strzałkowych, których nieskończoną liczbę możemy przeprowadzić po obu stronach ciała, równolegle do płaszczyzny pośrodkowej. Jeżeli przeprowadzimy oś poziomą poprzecznie prostopadle do linii pośrodkowej ciała, to liniete określają płaszczyznę czołową, mniej więcej równoległą do płaszczyzny czoła. Płaszczyzn czołowych równoległych do opisanej możemy przeprowadzić bardzo wiele, bliżej czy dalej w stosunku do ściany przedniej tułowia. Wreszcie trzeci rodzaj płaszczyzn, to płaszczyzny poziome, określone osiami strzałkową i poprzeczną, równoległe do podstawy. Liczba tych płaszczyzn również może być bardzo duża, gdyż można je przeprowadzić na dowolnej wysokości. Na każdej z tych płaszczyzn określamy kierunki. Na każdej płaszczyźnie strzałkowej możemy zaznaczyć kierunek przedni i tylny (oznaczany czasem jako brzuszny i grzbietowy). Na płaszczyznach czołowych odróżniamy, podobnie jak na strzałkowych, kierunek górny (prowadzący do głowy) i dolny w stronę stóp. Wreszcie na płaszczyznach poziomych odróżniamy kierunki boczny i przyśrodkowy oraz przedni i tylny, podobnie jak na płaszczyznach strzałkowych. Na kończynach m ó w i m y o kierunkach bliższym i dalszym, określanym w stosunku dó tułowia. Te same określenia stosujemy w stosunku do położenia na płaszczyznach czy w przestrzeni. M ó w i m y więc np., że żołądek leży w górnej części jamy brzusznej, nerki leżą na jej ścianie tylnej, płuca są położone bocznie do śródpiersia itp. Określając położenie narządów, stosujemy również nazwy: zewnętrzny, wewnętrzny i środkowy, powierzchowny i głęboki, podłużny i skośny oraz dla oznaczenia strony - prawy i lewy.
Okolice ciała Aby umożliwić określenie miejsca na powierzchni ciała, korzystamy z umownych linii pionowych oraz poziomych. Z linii pionowych poznaliśmy: 1. Linię pośrodkową przednią i 2 - tylną. Przebiegają one w płaszczyźnie 24
symetrii na powierzchni przedniej i tylnej, a więc pierwsza przez środek czoła, nosa, warg, bródki, szyi, mostka i wzdłuż kresy białej brzucha do środka spojenia łpnowego, druga zaś od miejsca połączenia obu kości ciemieniowych i kości potylicznej do kręgosłupa i wzdłuż kręgosłupa do szczytu kości guzicznej. Do nich dochodzą jeszcze następujące: 3. Linia s u t k o w a ( linea mamillaris), biegnąca pionowo przez brodawkę sutkową, leżącą zazwyczaj u mężczyzn na wysokości czwartego międzyżebrza. Ze względu na to, że u kobiet położenie brodawki sutkowej jest zmienne zależnie od rozwoju i czynności gruczołu sutkowego, możemy określić położenie tej linii, opuszczając ją pionowo ze środka obojczyka, jako linię środkowoobojczykową (linea medioclavicularis), która pokrywa się z linią sutkową. 4. Linia pachowa ( linea axillaris) biegnąca pionowo po ścianie bocznej klatki piersiowej od szczytu dołu pachowego do jej brzegu dolnego. 5. Linia ł o p a t k o w a (linea scapularis), biegnąca pionowo po powierzchni tylnej klatki piersiowej przez kąt dolny łopatki, równolegle do kręgosłupa. O m ó w i o n e linie pozwalają się zorientować co do położenia określanego punktu w poszczególnych okolicach, których nazwy są powiązane z nazwami ciała. Odróżniamy więc okolice głowy, szyi, klatki piersiowej itd., które kolejno omówimy. Okolice g ł o w y . Odróżniamy tu okolice nieparzyste - czołową i potyliczną (regio frontalis et r. occipitalis), oraz parzyste - ciemieniowe i skroniowe (r. parietalis et r. temporalis). Odpowiadają one mniej więcej kościom czaszki 0 tych nazwach. Granice ich są zaznaczone na ryc. 12. Okolice twarzy. Okolica nosowa (r. nasalis) obejmuje grzbiet, powierzchnie boczne i podstawę nosa. Okolica ustna (r. oralis) otacza szparę ustną, a więc obejmuje wargi i kąty ust. Okolica bródkowa (r. mentalis) leży na bródce, pod okolicą ustną. Okolica oczodołowa (r. orbitalis) obejmuje powierzchnię ograniczoną brzegami oczodołu, od góry graniczy z okolicą czołową, od przyśrodka - z nosową, zaś od dołu - z następną okolicą podoczodołową (r. infraorbitalis), która od strony bocznej sąsiaduje z okolicą jarzmową (r. zygomatica). Do, tyłu od jarzmowej leży wymieniona już okolica skroniowa. Bocznie od okolicy ustnej przechodzimy na okolicę policzkową (r. buccalis), która graniczy z okolicą przyuszniczo-żwaczową (r. parotideomasseterica), dochodzącą do tylnego 1 dolnego brzegu żuchwy. Okolice szyi. Granicę między twarzą i szyją stanowi łatwo wyczuwalny brzeg dolny żuchwy. Na szyi rozróżniamy cztery parzyste okolice. Nad skośnie przebiegającym w a ł e m mięśnia mostkowo-obojczykowo-sutkowego leży okolica tej samej nazwy (r. sternocleidomastoidea). Między nią i linią pośrodkową ciała leży okolica przednia szyi. (r. collianterior), ograniczona od góry brzegiem żuchwy. W tej okolicy odróżniamy trójkąt podżuchwowy ( t r i g o n u m submandibulare), leżący pod brzegiem żuchwy i nieco niżej trójkąt tętnicy szyjnej (fr. caroticum). Bocznie od okolicy mostkowo-obojczykowo-sutkowej leży okolica boczna szyi (r. collilateralis), dochodząca do wyraźnie odcinającego się brzegu mięśnia czworobocznego, przyczepiającego się do obojczyka. Granicę dolną tej okolicy stanowi obojczyk. Okolicę tę zajmuje trójkątłopatko25
wo-obojczykowy (tr. omoclaviculare), zwany też dołem nadobojczykowym większym, szczególnie dobrze widocznym u osób szczupłych przy pociągniętych ku górze barkach. Do tyłu od tego trójkąta rozciąga się okolica tylna szyi (r. colli posterior), zwana potocznie okolicą karku. Okolice klatki piersiowej. Okolica podobojczykowa (r. infraclavicularis) leży w dole tej samej nazwy i graniczy od dołu z okolicą sutkową (r. mammalis), dochodzącą do poziomu dolnego końca mostka. Między ramieniem a ścianą boczną klatki piersiowej leży dół pachowy ( fossa axillaris), ograniczony powierzchnią klatki piersiowej, zajętą przez okolicę pachową (r. axillaris). Okolice brzucha. Jeżeli na ścianie przedniej tułowia przeprowadzimy poprzeczne linie poziome: jedną - przez podstawę wyrostka mieczykowatego mostka, drugą - przez najniższe punkty boczne klatki piersiowej i trzecią przez kolce biodrowe górne przednie, to podzielimy ścianę przednią brzucha na trzy obszary: górny - nadbrzusze ( e p i g a s t r i u m ) , środkowy - śródbrzusze (mesogastńum) i dolny - podbrzusze ( h y p o g a s t r i u m ) . Następnie przedłużając linię sutkową z każdej strony do pachwiny, dzielimy każdy z w y m i e n i o n y c h obszarów na trzy pola stanowiące dziewięć okolic. W części pionowej środkowej leży u góry okolica nadbrzusza (r. epigastrica), pod nią okolica pępkowa (r. umbilicalis) i najniżej okolica łonowa (r. pubica). W częściach bocznych znajdujemy okolice parzyste (prawe i lewe) u góry podżebrową (r. hypochondriaca), niżej - boczną brzucha (r. lateralis abdominis) i u dołu - pachwinową (r. inguinalis). Okolice grzbietu. Część pośrodkową grzbietu obejmuje okolica kręgowa (r. vertebralis). Na kości krzyżowej znajdujemy okolicę krzyżową (r. sacraliś). Bocznie od okolicy kręgowej i do dołu od granicy z karkiem aż do poziomu kąta dolnego łopatki leży okolica łopatkowa (r. scapularis). Poniżej łopatki, na klatce piersiowej leży okolica podłopatkowa (r. subscapularis), a do kości miednicznej ciągnie się okolica lędźwiowa (r. lumbalis). Okolica kroczowa (r. perinealiś) ma kształt ukośnika, leżącego na d o l n y m końcu tułowia. Ograniczają ją linie łączące szczyt kości guzicznej z guzami kulszowymi, oraz guzy kulszowe z brzegiem d o l n y m spojenia łonowego. Linia poprzeczna łącząca guzy kulszowe dzieli.krocze na dwa trójkąty, z których przedni stanowi okolicę moczowo-płciową (r. urogenitalis), tylny zaś okolicę odbytową [r. analis). Okolice kończyny górnej. Bark jest zajęty przez okolicę naramienną (r. deltoidea). Ramię ma okolicę ramienną przednią (r. brachiianterior) i ramienną tylną (r. brachii posterior). Od stawu łokciowego biorą nazwę otaczające go okolica łokciowa tylna (r. cubitiposterior) i okolica łokciowa przednia (r. cubiti anterior), której środek zajmuje dół łokciowy ( fossa cubitalis). Na przedramieniu przy dłoni odwróconej do przodu w pozycji stojącej odnajdujemy okolicę przednią przedramienia (r. antebrachii anterior) i okolicę tylną przedramienia (r. ante brachii posterior). Wreszcie na ręce odróżniamy grzbiet ręki ( d o r s u m manus) - i dłoń ( palma manus). Kończyna dolna. Powierzchnię zewnętrzną miednicy zajmuje okolica pośladkowa (r. glutea). Przednią i tylną powierzchnię uda obejmują okolice tej 26
<-
samej nazwy (r. femoris anterior et posterior). Okolice przednia i tylna kolana (r. genus anterior et posteriori otaczają staw kolanowy. Między stawem kolanowym i stopą leżą okolice przednia i tylna goleni (r. cruris anterior et posterior). Na stopie leży okolica piętowa (r. calcaneus), wreszcie odróżniamy na niej grzbiet stopy (dorsum pedis) i podeszwę (planta pedis).
Budowa i proporcje ciała w zależności od wieku W organizmie ż y w y m toczą się bez przerwy różne procesy, w których przejawia się życie. Czasami natężenie ich się zmniejsza, jak np. praca serca i płuc w czasie snu, albo się zwiększa, jak przy ciężkiej pracy. Inne pracują z przerwami, jak np. żołądek. Wynikiem tych procesów są też zmiany zachodzące w organizmie. Jak już w i e m y , życie organizmu rozpoczyna się z chwilą zapłodnienia komórki jajowej przez plemnik. Początkowe f o r m y zarodka w niczym nie przypominają człowieka (ryc. 14). Po czterech tygodniach zarodek ma kształt silnie zagiętego walca. U zarodka pięciotygodniowego, którego długość prawie podwoiła się w ciągu tygodnia, można już poznać zarysy części ciała (ryc. 14). Silnie zgięta głowa, stanowiąca prawie połowę długości tułowia, jeszcze bez zarysów szyi, przylega do dużego uwypuklenia ściany przedniej tułowia - guza sercowego, opierającego się na pępowinie, pojawiają się też pierwsze zarysy kończyn.
r
R
j *
Ryc. 13. Względne rozmiary jaja u człowieKa, ryb i ptaków.
Tl
Zarodek sześciotygodniowy ma już wyraźnie zaznaczone kończyny, chociaż jeszcze bardzo małe, natomiast znacznie lepiej rozwiniętą głowę, której szczegóły wybitniej zarysowują się u zarodka siedmiotygodniowego. Wreszcie zarodek ośmiotygodniowy ma już kształty ludzkie, chociaż o całkiem innych proporcjach. Głowa jest prawie tej wielkości co tułów, a kończyny są więcej niż o połowę krótsze od tułowia. Część twarzowa głowy jest bardzo mała w stosunku do części mózgowej, wąskie fałdy powiekowe nie zasłaniają jeszcze oczu (ryc. 14).
Ryc. 14. Wczesne stadia rozwoju zarodka człowieka. A- 4 tyg., 6 - 5 tyg., C- 6 tyg., D-7 tyg., £ - 8 tyg.
Dalszy okres rozwoju zarodka polega na nierównomiernym wzrastaniu części ciała i jego narządów, a więc np. wolniejszy rozwój głowy, natomiast szybszy tułowia i kończyn, wolniejszy wzrost wątroby, zaś szybszy przewodu jelitowego itp., co doprowadza do ustalenia się proporcji właściwych dla noworodka i rozwoju narządów do stanu, w którym dalszy ich rozwój może odbywać się poza organizmem matczynym. W t y m okresie rozwijający się organizm nazywamy płodem ( foetus). 28
3
4
5
6
7
8
9
10
Ryc. 15. Wzrost i rozwój zewnętrznych kształtów ciała w okresie płodowym (cyfry oznaczają miesiące księżycowe).
Zmiany zachodzące w organizmie w ciągu życia Nieprzerwane procesy biologiczne zachodzące w organizmie prowadzą do szeregu zmian zarówno w jego budowie, jak i czynnościach. Inna jest budowa noworodka, inna człowieka dojrzałegó, podobnie jak i czynności rozmaitych narządów. Dlatego też nie można dać jednego ogólnego opisu człowieka, tylko należy uwzględnić zmiany zachodzące w jego budowie i czynnościach narząd ó w związane z wiekiem osobnika. Z tych względów życie osobnika dzielimy na pięć okresów, występujących kolejno i przechodzących stopniowo bez wyraźnych granic w następny. Po urodzeniu się osobnik wchodzi w w i e k dziecięcy ( i n f a n t i a ) , który trwa do okresu pokwitania. Wiek ten należy podzielić na wyraźnie różniące się od siebie podokresy: wiek oseska, wczesnego dzieciństwa i późnego dzieciństwa. Wiek oseska trwa od urodzenia do początku ząbkowania, które normalnie rozpoczyna się w 6 - 8 miesiącu życia. W t y m czasie toczą się w organizmie dziecięcym niezmiernie intensywne procesy wzrastania, wyrażające się w przyroście długości i ciężaru ciała. W ciągu pierwszych sześciu miesięcy wzrost noworodków płci męskiej (c?) powiększa się średnio o 15 cm,u nieco mniejszych noworodków płci żeńskiej ( $ ) o 16 c m , a c i ę ż a r s i ę p o d w a j a . W t y m też okresie (około 1 1 / 2 mies.) osesek zaczyna podnosić główkę, co prowadzi do wytworzenia się normalnej krzywizny szyjnej kręgosłupa. Wiek wczesnego dzieciństwa trwa od początku ząbkowania do momentu wyrzynania się pierwszych zębów stałych, które następuje w 6 - 7 roku życia 29
Płód 2 mies.
Płód 4 mies.
Noworodek
2 rok życia
6 lat
12 lat
25 lat
Ryc. 16. Dwa okresy życia płodowego i pięć okresów po urodzeniu. Dla ukazania charakterystycznych zmian proporcji różnych części ciała w różnych okresach rozwoju postacie przedstawiono w jednakowych rozmiarach.
dziecka. W t y m okresie dziecko rozpoczyna chodzić (koniec pierwszego początek drugiego roku życia), co prowadzi do wytworzenia się krzywizny lędźwiowej kręgosłupa. W t y m też okresie 6-12-krotnie powiększa się większość narządów szyi, klatki piersiowej, j a m y brzusznej i miednicy, zaś jeszcze silniej wzrastają przewód pokarmowy, kościec i mięśniówka. Trzewia szyi i serce są ustawione wysoko. Również wysoko ustawiona jest stosunkowo duża (zwłaszcza we wcześniejszych okresach) wątroba. Klatka piersiowa jest krótka i zaokrąglona, podściółka tłuszczowa obfita. Wiek późnego dzieciństwa trwa od początku wyrzynania się zębów stałych do pojawienia się 28 zębów (brak jeszcze zębów mądrości), co zbiega się z początkiem pokwitania (12-14 lat). Wiek młodzieńczy (iuvenilitas) trwa od czasu wyrżnięcia się 28 zębów (14-16 lat) do ukończenia wzrostu wysokości (20-25 lat). Wiek dorosły ( v i r i l i t a s ) i w i e k dojrzały (maturitaś) - to lata największej wydolności i przydatności do pracy. Przypada on na lata 25-55 u mężczyzn i 20-50 u kobiet. Granica między t y m i okresami nie jest wyraźna i u różnych osobników występuje w różnych latach. Przypada ona na pierwsze słabe objawy starzenia się, jak początek kostnienia szwów czaszki. Wiek dojrzały trwa do czasu przekwitania ( c l i m a c t e r i u m ) u kobiet i do występowania zmian starczych u obu płci, jak wypadanie w ł o s ó w , silne zużycie zębów, kostnienie szwów czaski, starczowzroczność, przyrost ciężaru ciała zwłaszcza u pykników. Górna granica wieku dojrzałego jest obecnie zmienna osobniczo i może się przesuwać w granicach 10 lat. W t y m okresie w organizmie dają się zauważyć zmiany wsteczne, które stopniowo wzmagają się w następnym okresie. 30
Nowo-4 mies. 8 mies. 1 rok 2 )4 roku 4 lata rodek
7 lat
12 lat
15 lat
Osobnik dorosły
Ryc. 17. Wzrost i rozwój zewnętrznych form ciała.
Wiek starości (senium) trwa od 55-60 lat do końca życia. Niektórzy rozróżniają trzy stopnie starości: pierwszy - t o lata 60-70, drugi - w ł a ś c i w a s t a r o ś ć do 80 lat i trzeci - po latach 80. Po 60 latach włosy siwieją, występują zmarszczki z p o w o d u zaniku tkanki tłuszczowej i zmian skóry, która traci napięcie. Narządy wewnętrzne starzeją się i są mniej sprawne, wzrost nieco obniża się, chociaż postawa ciała może jeszcze pozostać sprężysta i prosta. Około 70 lat w ł o s y bieleją, stają się rzadsze, łysina się powiększa, zazwyczaj występuje wyraźne pochylenie tułowia lub nawet starcze wygięcie kręgosłupa piersiowego. Napięcie mięśni zmniejsza się, występuje zwiotczenie więzadeł. W ostatnim okresie występują wyraźne objawy zanikowe, chociaż często sprawność umysłowa bywa zachowana.
Symetria i asymetria Już wyżej w s p o m n i a n o o t y m , że ciało ludzkie wykazuje budowę dwustronnie symetryczną, to znaczy, że połowy przedzielone płaszczyzną pośrodkową mają się do siebie, jak odbicia w lustrze. Stąd też człowiek ma szereg narządów parzystych, jak oczy, uszy, kończyny i inne. Jednakta symetria nie jest idealna. 31
Zwykle kończyna górna prawa jest nieco dłuższa i silniejsza, w przeciwieństwie do kończyny dolnej; ponieważ stopa lewa w większości przypadków jest nieco dłuższa, obuwie należy przymierzać na lewą nogę. Również i twarz nie jest całkowicie symetryczna - często dostrzegamy asymetrię, zwłaszcza w budowie nosa, oczu lub uszu. Dotyczy to również innych szczegółów. Natomiast znaczną asymetrię dostrzegamy w położeniu i budowie niektórych narządów wewnętrznych. Niektóre narządy nieparzyste zarodka, jak serce, wątroba, śledziona, żołądek, jelito, wyrostek robaczkowy, zmieniają swe położenie i leżą niesymetrycznie: serce w większej części po stronie lewej, wątroba po prawej, śledziona i żołądek po stronie lewej, wyrostek robaczkowy z jelitem ślepym - po stronie prawej, zaś pętle jelita cienkiego - nieregularnie układają się w jamie brzusznej. / Różnice te tłumaczymy głównie zmianami rozwojowymi, uzależnionymi wzajemnie. Tak więc płuco lewe jest mniejsze od prawego, gdyż wcześniej rozwijające się serce zajmuje więcej miejsca po stronie lewej klatki piersiowej. Nerka prawa leży niżej od lewej z p o w o d u silnie rozwijającej się po stronie prawej wątroby, która spycha nerkę prawą ku dołowi.
Metameria We wczesnych okresach rozwoju zarodka dostrzegamy występowanie u niego w części tylnej okolicy g ł o w o w e j kształtowanie się odcinków o podobnej budowie, formujących się w miarę rozwoju zarodka w kierunku od zawiązka głowy do końca tułowia. Te odcinki noszą nazwę praczłonów altto s o m i t ó w . Powstają one ze środkowego listka zarodkowego, który daje zawiązek kośćca, mięśni i skóry, powstających z poszczególnych somitów. Budowa organizmu z s o m i t ó w w dalszym ciągu się zaciera, jednak niektóre jej szczegóły pozostają w organizmie ludzkim na stałe. Widzimy więc, że • kręgosłup składa się z podobnych w zasadzie do siebie kręgów, między którymi przebiegają podobne więzadła. Z kręgami są powiązane żebra kostne, które są w pełni rozwinięte w części piersiowej kręgosłupa, natomiast w innych jego częściach występują w postaci szczątkowej. W obrębie klatki piersiowej żebra są połączone ze sobą pasmami mięśniowymi, tworzącymi mięśnie międzyżebrowe zewnętrzne i wewnętrzne. Również w miedzyżebrzach przebiegają naczynia krwionośne i nerwy międzyżebrowe, zaopatrujące mięśnie i pokrywające je pasma skófy. T w o r y te, stanowiące podobne do siebie części ściany klatki piersiowej, nazywamy metamerami (greckie: metapo, meros - część, a więc kolejne części); m ó w i m y więc, że klatka piersiowa ma budowę metameryczną. Ślady tej b u d o w y znajdziemy i w innych częściach ciała, natomiast nie widzimy ich w budowie kończyn.
32
Ryc. 18. Krzyż św. Andrzeja - kanon rzymskiego architekta Witruwiusza uzupełniony przez Leonarda da Vinci, który dorysował postać w rozkroku z uniesionymi nieco ramionami. Wokół tej figury nakreślił koło, którego środek znajduje się w pępku. Wysokość głowy wg tego kanonu jest mała (Vg wysokości ciała).
Proporcje ciała ludzkiego i ich zmiany w ciągu życia Od dawna wielcy malarze i rzeźbiarze, którzy byli jednocześnie znawcami anatomii lub też badaczami ciała ludzkjego, jak Leonardo da Vinci, Michał Anioł Buonarotti, czy Rafael Santi, dążyli do ustalenia klasycznych, a więc najbardziej harmonijnych, proporcji ciała ludzkiego. Taki „ k a n o n " , w którym człowiek z rozkrzyżowanymi kończynami dawał się wpisać w koło, ustanowił Leonardo. Dzisiaj przyjmujemy, że siąg, czyli odległość między końcami opuszek palców środkowych rąk przy poziomo odwiedzionych kończynach równa się w przybliżeniu wysokości ciała. Jednak należy zaznaczyć, że proporcje te można określić tylko bardzo ogólnie, gdyż zależą one od szeregu czynników dziedzicznych, konstytucyjnych i zewnętrznych, a także płci i przede wszystkim od wieku. W i e m y już, że płód ma stosunkowo dużą głowę, która w wieku 2 miesięcy życia płodowego w y n o s i tyle, co cały t u ł ó w z kończynami dolnymi. Natomiast wysokość g ł o w y człowieka dorosłego w y n o s i Vs wysokości całego ciała. 3 - Anatomia i fizjologia
33
Z tego widać, że podczas wzrastania osobnika głowa rośnie znacznie wolniej niż tułów i kończyny dolne. Wysokość głowy noworodka wynosi 1 / 3 część długości jego tułowia z kończynami. Kończyny.dolne płodu dwumiesięcznego stanowią mniej niż V 4 jego długości, u noworodka około 3 / 8 , podczas gdy u człowieka dorosłego stanowią one połowę długości ciała. A więc w miarę wzrastania zmniejsza się stosunkowo wysokość głowy, a zwiększa się długość kończyn dolnych. Zmieniają się również proporcje obu kończyn. U płodu kończyny dolne są znacznie krótsze od górnych i choć te różnice w ciągu życia płodowego zmniejszają się, to jeszcze noworodek ma kończyny dolne krótsze od górnych. Dopiero w pierwszych pięciu latach życia dziecka długość kończyn staje się mniej więcej równa, a w dalszych latach długość kończyn dolnych zaczyna przeważać. Jak wspomniano, na proporcje ciała wpływa również płeć. Otóż proporcje i kształty ciała kobiet są bardziej zbliżone do występujących u dzieci niż u mężczyzn. Kończyny dolne u kobiet są stosunkowo krótsze niż u mężczyzn, również klatka piersiowa na przekroju poziomym jest u kobiet, podobnie jak u małych dzieci, bardziej okrągła, podczas gdy u mężczyzn jest bardziej płaska.
Różnice konstytucyjne budowy ciała Jak podano wyżej, proporcje ciała ludzkiego zależne są również od typu konstytucyjnego. Pewne proporcje wiązano ze skłonnością do pewnych chorób, czy też uważano za wynik procesów chorobowych. Były różne próby ustalenia typów konstytucyjnych; na ogół można jednakzgodzićsięz Kretschmerem, że wszystkie formy dadzą się ująć w trzy typy, które pokrótce' opiszemy. Typ leptosomiczny odznacza się większymi wymiarami długościowymi, natomiast mniejszymi szerokościowymi i obwodami. Leptosomicy mają postać wydłużoną, długie kończyny dolne, długą wąską klatkę piersiową, wąskie opadające barki. Szyja długa, cienka, twarz wąska, długa, nos duży, wąski, wargi cienkie. Na ogół szczupli, nie mają skłonności do tycia. Typ eurysomiczny, czyli pykniczny charakteryzują, w przeciwieństwie do leptosomicznego, mniejsze wymiary długościowe, natomiast większe szerokościowe i obwody. Pyknicy mają budowę krępą, przysadzistą, krótkie grube kończyny, duży obwód brzucha, klatki piersiowej i szyi, co spowodowane jest również skłonnością do odkładania się tłuszczu. Klatka piersiowa krótka, szeroka i głęboka, kąt podmostkowy rozwarty. Barki szerokie, kark gruby. Twarz szeroka, okrągła, cera rumiana, nos krótki, szeroki, oczy płytko osadzone. Typ atletyczny charakteryzuje jego nazwa. Stanowi on rodzaj pośredni między opisanymi poprzednio, zbliżony do typu klasycznego lekkoatlety. Budowa wysmukła, harmonijna, kościec i mięśnie dobrze rozwinięte, klatka piersiowa dobrze wysklepiona, kąt podmostkowy prosty. Barki szerokie. 34
natomiast mniejszy o b w ó d w pasie i wąskie biodra. Szyja miernego o b w o d u i długości, twarz owalna, nos prosty, wargi mierne. Należy zaznaczyć, że opisane typy konstytucyjne występują w czystej postaci niezbyt często, natomiast typy mieszane spotyka się w różnych formach, z przewagą tego czy innego typu. Typ eurysomiczny nieraz występuje w latach starszych, kiedy bardziej przejawia się skłonność do tycia.
PRZEGLĄD UKŁADÓW NARZĄDÓW Jak już w i e m y , liczne narządy ciała ludzkiego, przeznaczone do rozmaitych czynności, wypełniają swe zadania łącząc się w większe czy mniejsze układy. Również układy narządów nie pracują samodzielnie, tylko łączą swe czynności z pracą innych układów. Nadrzędnym układem jest układ nerwowy, który reguluje wszystkie czynności organizmu, a poza t y m jest siedliskiem świadomości. Jedynie harmonijna współpraca wszystkich narządów prowadzi cfo normalnych czynności całego organizmu. Pamięta o t y m lekarz, który nie leczy chorego narządu, lecz chory organizm. Rozróżniamy następujące układy narządów: 1. Układ narządów ruchu. 6. Układ nerwowy. 2. Układ trawienny. 7. Układ gruczołów dokrewnych. 3. Układ oddechowy. 8. Układ narządów zmysłów. 4. Układ naczyniowy. 9. Powłoka wspólna. 5. Układ moczowo-płciowy.
Układ narządów ruchu Najłatwiej dostrzegalnym przejawem życia jest ruch, którego źródłem jest tkanka mięśniowa, z której są zbudowane mięśnie. Są one przeważnie umocowane na kościach, które odgrywają rolę dźwigni, nie mogących poruszać się samodzielnie. Dlatego też omawiany układ dzielimy na układ narządów ruchu biernego, do którego należą kości, stawy i wiązadła, oraz czynnego, który tworzą mięśnie szkieletowe. Nauka o kościach ( o s t e o l o g i a ) dzieli się na ogólną i szczegółową. W t y m przeglądzie o m ó w i m y jedynie podstawy osteologii ogólnej, natomiast poszczególne kości będą o m ó w i o n e w osobnym rozdziale. Kości (ossa) obok zębów są najbardziej wytrzymałymi narządami organizmu, z których zbudowany jest szkielet, narząd p o d p o r o w y ciała. Na zgniatanie kość jest wytrzymalsza od granitu, na zgięcie odporniejsza od dębu, natomiast najsłabsza jest na rozciąganie. Obok tak dużej wytrzymałości kość jest również sprężysta. Nieduże odkształcenia, np. małe wygięcie kości wraca do n o r m y natychmiast po ustaniu siły działającej. Jeżeli siła ta przekracza odporność kości, ulega ona złamaniu. Te właściwości kość zawdzięcza s w y m składnikom - organicznej chrząstce 35
kostnej, zwanej oseiną i solom wapnia (głównie w ę g l a n o m i fosforanom). Oseina nadaje kościom sprężystość, sole wapnia - twardość i odporność. Kości w wieku m ł o d y m zawierają więcej oseiny i są dzięki temu bardziej elastyczne, natomiast u ludzi starych łatwiej ulegają złamaniom. Pod względem kształtu dzielimy kości na długie, krótkie i płaskie. Kości długie leżą głównie w kończynach; do płaskich należą kości pokrywy czaszki, łopatka; kości różnokształtne, jak np. kręgi i kości nadgarstka zaliczamy do kości krótkich. Kości długie składają się z trzonu ( corpus) i d w ó c h k o ń c ó w (extremitas). Trzon jest zbudowany z istoty zbitej (p. Tkanki), natomiast końce - z istoty gąbczastej, pokrytej pa powierzchni kości istotą gąbczastą. Wewnątrz trzonu znajduje się jama szpikowa (cavum medullare), wypełniona szpikiem ż ó ł t y m (medulla ossium flava), stanowiącym substancję tłuszczową. U płodu jama szpikowa jest wypełniona szpikiem c z e r w o n y m (medulla ossium rubra), w y twarzającym krwinki czerwone - erytrocyty. Kości krótkie i płaskie są zbudowane podobnie do końców kości długich; w siateczce istoty gąbczastej leży szpik czerwony. Kości są pokryte mocną błoną łącznotkankową, zwaną okostną (periosteum). Jest ona obficie unaczyniona i unerwiona, od niej też w głąb kości wchodzą naczynia odżywne. Kość pozbawiona okostnej, a t y m samym źródła odżywiania, obumiera. Na wewnętrznej powierzchni okostnej leży warstwa komórek kościotwórczych (osteoblastów), które mogą budować tkankę kostną w okresie wzrastania kości lub łączyć jej części po złamaniach. Połączenia kości. Niektóre kości przeznaczone do ochrony wrażliwych narządów, jak np. kości czaszki, są połączone ze sobą nieruchomo, tworząc szwy (suturae). Inne są połączone ze sobą s t a w a m i (articulationes), pozwalającymi na ruchy w różnych zakresach. W niektórych stawach ruchy zachodzą dokoła jednej tylko osi, w innych dokoła dwóch czy większej liczby osi. Nazywamy je odpowiednio stawami jedno-, dwu- lub wieloosiowymi. Do stawów jednoosiowych należą np. stawy palców ręki, które możemy zginać i prostować. Przykładem stawu dwuosiowego jest staw nadgarstkowy, w którym możemy rękę zgiąć (w kierunku dłoni) i wyprostować (w kierunku grzbietu) oraz odwodzić i przywodzić rękę (w stronę kciuka i palca małego). Wreszcie w stawach wieloosiowych, jak ramienny i biodrowy, możemy wykonywać dowolne ruchy: zgięcia i prostowania (do przodu i do tyłu), odwodzenia i przywodzenia (ku bokowi i do tułowia), oraz o b r o t ó w (na zewnątrz i do wewnątrz). Staw łączy dwie lub więcej kości, na których występują gładkie, ograniczone powierzchnie stawowe. Jedna z kości ma na końcu wypukłą g ł o w ę s t a w o w ą (caput articulare) w postaci wycinka kuli, walca lub bardziej skomplikowaną, druga - odpowiednio zagłębione wydrążenie, zwane panewką czy wydrążeniem s t a w o w y m (cavitas glenoidalis). Zarówno głowa, jak i wydrążenie są pokryte szklistą chrząstką s t a w o w ą (cartilago articularis). Chrząstki są zwilżone śliską mazią s t a w o w ą ( s y n o v i a ) , dzięki czemu tarcie w stawie między głową a panewką jest minimalne. 36
rj •ć
i;
ll It .. MSi2 '•W 1S
f i %
1 1
Ryc. 19. Stawy człowieka o różnym stopniu swobody ruchów: 1 - stawy wieloosiowe, 2 - stawy jednoosiowe, 3 - staw dwuosiowy.
Staw otacza torebka s t a w o w a ( capsula articularis), złożona z zewnętrznej warstwy włóknistej i wewnętrznej maziowej. Przytrzymuje ona kości względem siebie i zrastając się z obu kośćmi zamyka jamę s t a w o w ą (cavum articulare), do której błona maziowa wydziela potrzebną ilość mazi. Torebka stawowa nie jest jedynym czynnikiem przytrzymującym kości w stawach. Dużą rolę gra też ciśnienie atmosferyczne, lecz najsilniej przeciw37
działają rozchodzeniu się kości (a więc i zwichnięciu stawu) mięśnie, które, przytrzymują kości tworzące staw. Również dużą rolę odgrywają silne nieraz więzadła wzmacniające torebkę stawową lub przebiegające wewnątrz stawu i bezpośrednio między kośćmi, jak to widzimy w stawie kolanowym.
Mięśnie Jak widzimy, same kości i stawy nie mogą wykonywać ruchu. Ruch może się odbywać pod wpływem określonych sił, np. siły ciężkości. Jeżeli uniesiemy kończynę górną do poziomu, a potem zwolnimy mięśnie, to kończyna opadnie. Podobnie opada głowa na piersi siedzącego w pociągu i zasypiającego pasażera. Są to ruchy bierne, zachodzące bez udziału mięśni, które stanowią
E
f-
G
Ryc. 20. Rodzaje mięśni. Układ włókien mięśniowych. A - m . wrzecionowaty, 6- m. dwubrzuścowy, C - m . półpierzasty, D - m . pierzasty, C - m . płaski, F - m . z e smugami ścięgnistymi, G - m . dwugłowy, H- m. okrężny.
38
narządy ruchu czynnego. Rozpoczynają się one i kończą na kościach, które stanowią układ dźwigni połączonych stawami. Mięśnie szkieletowe są zbudowane ztkanki mięśniowej poprzecznie prążkowanej, zależnej od naszej woli. Jednostką czynnościową jest mięsień (musculus). Mięśnie mają różne kształty. Najczęściej są to wydłużone t w o r y walcowate, grubsze w środku, przechodzące na końcach w mocne ścięgna ( t e n d i n e s ) , zbudowane z tkanki łącznej włóknistej. Mięśnie tego rodzaju nazywamy wrzecionowatymi. Występują one głównie na kończynach. Część środkowa mięśnia wrzecionowatego nosi nazwę brzuśca ( venter ), koniec bliższy nazyw a m y głową (caput), koniec dalszy ogonem ( cauda ). Mięsień mający dwa przyczepy bliższe nazywamy d w u g ł o w y m (m. biceps), trzy - t r ó j g ł o w y m (m. triceps), cztery - c z t e r o g ł o w y m ( m. quadriceps). Jeżeli koniec dalszy mięśnia dzieli się na kilka ścięgien, m ó w i m y o mięśniu wieloogoniastym. W ścianach tułowia napotykamy mięśnie o kształcie błon, zwane mięśniami płaskimi (musculi piani). Mięsień płaski nie ma brzuśca ani głów, a jego włókna przechodzą w cienkie, płaskie ścięgna, zwane rozcięgnami ( aponeuroses). Działanie mięśni. Mięśnie przyczepiające się do dwóch kości, przy skurczu skracają się i zbliżają te kości do siebie. Jak już w i e m y , ruchy te są określone budową stawów i kształtami kości, do których się przyczepiają. Dla wyjaśnienia działania mięśni rozpatrzymy przykłady dźwigni, na których zasadach one pracują. Dźwignią z punktu widzenia mechaniki jest każda kość
Ryc. 21. Rodzaje dźwigni: A - dźwignia równoważna, B - dźwignia siłowa, C - dźwignia szybkościowa.
39
długa, jako belka sztywna podparta na nieruchomym punkcie oporu. Jeżeli punkt ten leży na j e d n y m z końców belki, to m a m y do czynienia z dźwignią jednoramienną, natomiast jeżeli leży on pod jakimkolwiek miejscem między końcami belki, to stanowi ona dźwignię dwuramienną (ryc. 21). Ramieniem dźwigni nazywamy więc część belki, leżącą między punktem oparcia i miejscem przyłożenia siły działającej na dźwignię, za pomocą której m a m y pokonać opór O dla wykonania pewnej pracy. Dźwignia jest zrównoważona, jeżeli siła i opór są równe, jak np. odważnik i masa t o w a r o w a na szalach wagi. Jeżeli do jednego z ramion dźwigni zrównoważonej przyłożymy siłę, to drugie ramię wykona ruch w o d w r o t n y m kierunku. Waga jest przykładem dźwigni dwuramiennej zrównoważonej; w nauce 0 mięśniach będziemy mieli do czynienia z dźwigniami różnoramiennymi. Zasada ich działania - p. ryc. 22. Są one zrównoważone, jeżeli iloczyn siły P przez ramię AB dźwigni będzie równy iloczynowi siły Pi przez ramię BC dźwigni. Z tego wynika, że przy dźwigni równoramiennej wystarczy bardzo mała siła do wyprowadzenia jej ze stanu równowagi, natomiast przy dźwigni różnoramiennej trzeba różnych sił do jej zastosowania. Przykładem może być staw łokciowy, którego schemat przedstawia ryc. 22: punkt podparcia B, wyrostek łokciowy AB i przedramię BC. Do wyrostka łokciowego przyczepia się bardzo silny mięsień t r ó j g ł o w y ramienia, PA. Przy jego skurczu zajmuje on pozycję PAh zaś przedramię - położenie AiCh Jest to przykład dźwigni szybkościowej, w której silny mięsień skraca się mało, a wykonuje obszerny ruch ręki, jak przy ciosie pięścią czy kuciu młotem. W dźwigni jednoramiennej zarówno punkt przyłożenia siły, jak i punkt przyłożenia oporu leżą na j e d n y m ramieniu dźwigni, między punktem oparcia 1 w o l n y m końcem dźwigni. Na t y m s a m y m przykładzie stawu łokciowego rozpatrzymy działanie mięśnia d w u g ł o w e g o ramienia. Rozpoczyna się on na łopatce u góry i kończy się na kości promieniowej w pobliżu stawu. Jest to silny mięsień, który m.in. działa na przedramię, powodując zgięcie stawu łokciowego. Przy nie obciążonej ręce przedramię działa jak dźwignia szybkościowa, np. przy rzutach czy uderzeniu rakietką pingpongową. Natomiast mięsień ramienno-promieniowy, rozpoczynający się na dolnej części kości ramiennej, a kończący się na kości promieniowej w pobliżu ręki, działa na staw łokciowy z dużą siłą, zginając jednak przedramię stosunkowo w o l n o . M a m y tu przykład dźwigni siłowej. Ruchy są w przeważających wypadkach wynikiem działania kilku lub większej liczby mięśni, które współdziałają jako synergiści. Mięśnie te przeważnie wykonują określone ruchy, jako zginacze, odwodziciele, zwieracze itp. Trzeba jednak pamiętać, że rozkurcz mięśnia nie powoduje ruchu przeciwnego, a więc •np. ustanie skurczu mięśnia zginacza palców ręki nie powoduje ich prostowania. Do tej czynności są prżeznaczone mięśnie przeciwnicze - antagoniści, a 'więc prostowniki, przywodziciele, rozwieracze itd. Czasem działają jednocześnie synergiści i antagoniści, a jeżeli napięcie ich jest jednakowo silne, to nie powodują one ruchu, a ustalają stawy, hamując czynności. Siła mięśnia nie zależy od długości, ale od liczby jego włókien. Znamy to 40
A
B
Ci Ryc. 22. Schemat działania m. trójgłowego w stawie łokciowym (dźwignia dwuramienna szybkościowa).
z codziennej obserwacji - mięśnie grube widzimy u ludzi silnych, natomiast cienkie u dzieci i osobników słabo wyćwiczonych. Liczba włókien wiąże się z przekrojem mięśnia: im większy jest przekrój, tym większa jest siła mięśnia. Takie stosunki spotykamy jednak nie we wszystkich mięśniach, lecz tylko w takich, gdzie włókna mięśniowe biegną równolegle do ich osi długiej (ryc.20). Najczęściej są to mięśnie wrzecionowate, których przekrój poprzeczny w najgrubszym miejscu jest jednocześnie przekrojem fizjologicznym, to znaczy trafiającym wszystkie Włókna prostopadle do ich przebiegu. Znamy też 41
p
••
wm
»
•
Xtt i
B
A
C
Ryc. 23- Schemat działania m. dwugłowego ramienia w stawie łokciowym (dźwignia jednoramienna szybkościowa).
takie mięśnie, w których włókna dochodzą do ścięgna skośnie na j e d n y m lub, na obu końcach. Są to mięśnie półpierzaste i pierzaste (ryc.20). Żeby wykonać ich przekrój fizjologiczny, należy robić przekroje poprzeczne w kilku miejscach, żeby wszystkie włókna były raz przecięte. Suma powierzchni przekrojów da przekrój fizjologiczny mięśnia. Stwierdzono eksperymentalnie, że mięsień o przekroju 1 cm 2 ma siłę ok. 10 kg (7-11,1 kg). Jest to tzw. jednostka siły mięśniowej, której wielkość jest zmienna i zależy od wieku, stanu ogólnego i stanu psychicznego osobnika (napięcie w o l i , stres itp.). Na podstawie przekroju fizjologicznego mięśnia można określić jego siłę. Wynikiem skurczu mięśnia jest ruch. Mięsień kurcząc się, jak już w i e m y , skraca się i grubieje i taki skurcz nosi nazwę izotonicznego. Jednak skurcz mięśnia może nie w y w o ł a ć ruchu, jak np. skurcz mięśni szczęk przy ich zaciśnięciu albo skurcz przy utrzymywaniu uniesionej ręki. Tego rodzaju skurcz nazywamy izometrycznym, tzn. nie zmieniającym długości. Pewna część włókien pozostaje w stanie skurczu nawet w spoczynku. Taki stan nosi nazwę napięcia stałego {tonus). Stopień jego jest różny i zależy od wieku, stopnia zmęczenia, jak również konstytucji osobnika. Napięcie to spada w czasie snu i może być zniesione w stanie omdlenia lub przez głęboką narkozę. Ustaje całkowicie po śmierci. Do układu narządów ruchu czynnego należą urządzenia pomocnicze mięśni. Składają się na nie wspomniane już ścięgna, pochewki ścięgien i mięśni, troczki ścięgien, bloczki, kaletki maziowe, powięzie. Pochewki otaczają ścięgna, chronią je i, praktycznie biorąc, znoszą ich tarcie. Pochewki mięśni otaczają mięśnie i przytrzymują je w określonym położeniu, wpływając w niektórych wypadkach na rodzaj ich ruchów. Powięzie otaczają grupy mięśniowe, łącząc je w grupy czynnościowe. Szczególnie silne są one na kończynach. Bloczki są to wyrostki kostne, oczka chrząstkowe lub pętelki ścięgniste, przez które przerzucają się ścięgna, zmieniając kierunki swego przebiegu. 42
«in
^ k ' ' c Ryc. 24. Schemat działania m. ramienno-promienjowego (dźwignia jednoramienna siłowa).
W miejscach, gdzie ścięgna mogą być uciśnięte lub ocierają się o kości przy ruchach, spotykamy włączone do ścięgna drobne zazwyczaj kostki, zwane trzeszczkami. Wyjątkowo dużą trzeszczką jest rzepka ( patella ), ochraniająca staw kolanowy, która pełni jednocześnie rolę bloczku dla ścięgna mięśni prostujących staw kolanowy.
Układ trawienny Do podtrzymania wszystkich procesów życiowych, jak w i a d o m o , niezbędny jest dopływ pokarmów, w o d y i tlenu. Woda i pokarmy są przyjmowane przez układ trawienny, do przyswajania tlenu służy układ oddechowy. Woda jest wchłaniana bezpośrednio przez narządy trawienia, w stanie nie 43
zmienionym, natomiast pokarmy muszą być strawione, czyli doprowadzone do stanu, w jakim mogą być wchłonięte przez jelita i włączone do krwiobiegu. Do tych celów służy skomplikowany układ narządów trawienia. Układ t r a w i e n n y jest w zasadzie cewą długości kilku metrów, o ścianch z błony śluzowej, mięśniowej i surowiczej, do której uchodzą przewody licznych gruczołów trawiennych. Pierwszym odcinkiem przewodu pokarmowego jest jama ustna ( cavum oris), przyjmująca pokarm. Mieści się w niej język (lingua) i zęby ( dentes), rozdrabniające pokarmy. Do jamy ustnej uchodzą przewody ślinianek ( glandulae salivares), którymi płynie ślina (saliva). Język za pomocą śliny f o r m u j e kęsy i przesuwa je do gardła ( p h a r y n x ) , gdzie zostają one przełknięte. Gardło jest w s p ó l n y m odcinkiem dróg pokarmowych i oddechowych, które tu się # krzyżują. Z gardła kęsy pokarmu przechodzą do przełyku i dalej do żołądka, gdzie pokarm ulega trawieniu mechanicznemu pod w p ł y w e m jego ruchów i chemicznemu pod działaniem soku żołądkowego ( succus gastricus). Częściowo strawiony pokarm, dzięki ruchom perystaltycznym żołądka, zostaje przesunięty do pierwszego odcinka jelita cienkiego - dwunastnicy ( duoder num), gdzie zostają strawione składniki pokarmów - białka, tłuszcze i węglowodany. Trawienie odbywa się za pomocą soków trawiennych, wydzielanych przez gruczoły j e l i t o w e (glandulae intestinaleś), trzustkę (pancreas) i w ą t r o b ę (hepar). Pod w p ł y w e m soków trawiennych składniki pokarmów o dużych cząsteczkach zostają rozbite na ciała prostsze, o małych cząsteczkach, które mogą przejść przez błony półprzepuszczalne jelita. W dalszych odcinkach jelita cienkiego (intestinum tenue) - w jelicie czczym i krętym (jejunum et ileum), następuje wchłanianie produktów trawienia do naczyń krwionośnych i chłonnych, przebiegających w ścianie jelita, które rozprowadzają je po całym organizmie. Pozostała treść jelita cienkiego przechodzi do jelita grubego ( intestinum crassum),\N którym następuje trawienie resztek pokarmów, wchłanianie w o d y i f o r m o w a n i e kału, wydalanego następnie przez odbyt (anus).
Układ oddechowy Do przemian fizykochemicznych, zachodzących w organizmie, jest niezbędny tlen, którego g ł ó w n y m źródłem jest powietrze atmosferyczne, pobierane przez układ oddechowy. Powietrze wchodzi przez nozdrza przednie do j a m y n o s o w e j (cavum nasi), podzielonej przegrodą nosa (septum nasi) i przepływa przez szereg.przestrzeni, ograniczonych m a ł ż o w i n a m i n o s o w y m i (conchae nasales). Małżowiny są pokryte bogato unaczynioną błoną śluzową. Przepływające powietrze zostaje ogrzane i nasycone parą wodną, powstającą z w y dzieliny błony śluzowej. Na wilgotnej błonie śluzowej osiadają też cząsteczki kurzu. Jamę nosową otaczają przestrzenie dodatkowe, zwane zatokami przynosow y m i (sinus paranasales), otrzymujące nazwy od kości, w których się znajdu44
ją. Są to zatoki parzyste: czołowa, klinowa, sitowa i największa z nich szczękowa. Zatoki te są wypełnione powietrzem, dzięki czemu chronią wnętrze czaszki przed oziębieniem powietrzem przepływającym przez jamę nosową. Grają one również rolę rezonatorów, nadających odmienną barwę głosu każdemu osobnikowi. Z j a m y nosowej klimatyzowane w opisany sposób powietrze przechodzi przez nozdrza tylne ( choanae ) do jamy gardła, gdzie krzyżuje się z drogą pokarmową. Dalej powietrze wchodzi do krtani (larynx), która jest narządem głosu, będąc jednocześnie odcinkiem granicznym górnych dróg oddechowych. Następnymi narządami układu są dolne drogi oddechowe, do których należy tchawica (trachea), dzieląca się na dwa oskrzela g ł ó w n e ( b r o n c h i principales), wchodzące do wnęk płuc, rozgałęziające się na drobniejsze oskrzela. Właściwymi narządami oddechowymi są płuca ( pulmones). Leżą one w jamie klatki piersiowej, w której znajduje się również serce. Każdy z tych trzech narządów leży w oddzielnej jamie surowiczej. Serce zajmuje jamę osierdzia, o czym będzie m o w a dalej. Każde płuco leży w zamkniętej przestrzeni, wysłane} błoną surowiczą, zwaną opłucną, która jako opłucna ścienna ( p l e u r a parietalis) wyścieła ściany Jamy opłucnej (cavum pleurae) i przechodzi na płuca jako opłucna płucna ( pleura visceralis). Płuco szczelnie wypełnia jamę opłucnej, która staje się jedynie szczeliną włosowatą, leżącą między w y m i e nionymi blaszkami opłucnej. W jamach opłucnej znajduje się trochę płynu surowiczego, ułatwiającego ruchy płuca, zachodzące wewnątrz klatki piersiowej podczas oddychania. Płuco prawe jest nieco większe od lewego, gdyż część jego miejsca zajmuje serce, leżące po stronie lewej klatki piersiowej. Wewnątrz płuca, jak w s p o m niano, oskrzela dzielą się na drobniejsze gałązki, którym towarzyszą naczynia krwionośne. Dochodzą one razem z oskrzelikami do pęcherzyków płucnych, w których zachodzi w y m i a n a gazów między krwią i powietrzem, wypełniającym pęcherzyki. Krew przepływająca przez naczynia włosowate, oplatające pęcherzyki płucne, oddaje do pęcherzyków dwutlenek węgla, a zabiera z nich tlen i odpływa do żył płucnych, a następnie do serca, jako krew natleniona. Przestrzeń leżąca między prawą i lewą jamą opłucnej nosi nazwę śródpiersia (mediastinum). Leży w n i m serce, grasica, przełyk, duże naczynia i nerwy.
Układ krążenia Wszystkie komórki organizmu ludzkiego potrzebują niezbędnych do życia materiałów, jak tlen, woda, związki organiczne i związki nieorganiczne. Jednocześnie muszą one wydalać produkty przemiany materii, zbędne lub nawet szkodliwe dla organizmu. Do zapewnienia tych warunków służy układ krążenia. Należą do niego naczynia krwionośne, w których krąży krew, wprowadzana w ruch przez silny narząd mięśniowy - serce, oraz naczynia chłonne, odprowadzające płyny tkankowe drogą chłonki (limfy) do krwi. Krew dostar45
cza komórkom tlen i pożywienie, jak również zabiera produkty przemiany materii z tkanek i przekazuje je narządom wydalającym je z organizmu (płuca, nerki, skóra). Krew, limfa i płyny tkankowe stanowią środowisko wewnętrzne organizmu. Krew ( sanguis) składa się z płynnego osocza i składników morfotycznych. Osocze, zawierające około 92% w o d y , składa się z surowicy (serum) i fibrynogenu. Fibrynogen jest ciałem białkowym, które po wypłynięciu z naczynia krwionośnego ścina się i tworzy skrzep. Do składników morfotycznych krwi należą elementy komórkowe, jak krwinki czerwone i białe oraz płytki krwi. Krwinki czerwone, erytrocyty, są to małe komórki o średnicy około 7,5 (im, zawierające barwnik zwany hemoglobiną. Hemoglobina tworzy nietrwałe połączenie z tlenem - oksyhemoglobinę. W atmosferze dwutlenku węgla oddaje ona tlen i łączy się z dwutlenkiem węgla, tworząc związek C0 2 -hemog lobinę. Na tej właściwości polega zdolność zabierania z tkanek C0 2 i oddawanie im tlenu oraz o d w r o t n y proces zachodzący w płucach, gdzie stężenie tlenu w powietrzu znajdującym się w pęcherzykach płucnych jest wysokie, zaś dwutlenku niskie. Krwinki czerwone rozwijają się w szpiku kostnym. Początkowo są to komórki jądrzaste, które w toku dojrzewania tracą jądra i trafiają do krwiobiegu jako komórki bezjądrzaste. Krwinki te żyją krótko, bo zaledwie około 100 dni, po czym zostają wyłapane z krwiobiegu przez wątrobę i przetwarzane. Krwinki białe dzielą się na granulocyty, limfocyty i monocyty. Granulocyty są nieco większe od krwinek czerwonych i mają 6 - 1 2 (j,m średnicy. Granulocyty mają duże płatowate jądra, a w protoplazmie ich znajdują się trzy rodzaje ziarnistości, które dają się wykryć barwnikami obojętnymi, kwaśnymi lub zasadowymi. Na tej podstawie rozróżniamy najliczniejsze granulocyty obojętnochłonne (neutrofile) oraz znacznie rzadsze kwasochłonne (eozynofile) i zasadochłonne (bazofile). Limfocyty są mniejsze od erytrocytów, mają duże kuliste jądra leżące bliżej o b w o d u komórki. Monocyty są największymi krwinkami białymi. Mają pasmo barwiące się, jądro nerkowatego kształtu oraz są obdarzone zdolnością ruchu i fagocytozy. Płytki k r w i (trombocyty) są bardzo małymi komórkami jądrzastymi o średnicy około 0,2 |xm. Biorą one udział w procesie krzepnięcia krwi. V
Serce Serce (cor) jest narzędziem mięśniowym, wtłaczającym krew do układu tętniczego. Chociaż jest ono zbudowane ztkanki mięśniowej poprzecznie prążkowanej (nieco różniącej się od mięśni szkieletowych), to jednak skurcze jego nie są zależne od woli. Serce ma kształt stożka, w którym rozróżniamy podstawę serca i jego koniuszek. Podstawa jest zwrócona ku górze, t y ł o w i i stronie prawej, koniuszek - ku dołowi, przodowi i stronie lewej. Jak wspomniano wyżej (p. Układ oddechowy), serce leży w śródpiersiu 46
w jamie osierdzia. Otoczone jest w o r k i e m osierdziowym ( p e r i c a r d i u m ) , zbud o w a n y m z d w ó c h w a r s t w - włóknistej i surowiczej. Warstwa zewnętrzna, zwana osierdziem w ł ó k n i s t y m ( p e r i c a r d i u m fibrosum), jest mocną błoną ochronną. Warstwa wewnętrzna (osierdzie surowicze) u podstawy serca przechodzi na jego ścianę jako nasierdzie ( e p i c a r d i u m ) . Między blaszką surowiczą osierdzia i nasierdziem znajduje się włosowata jama osierdzia, w której jest nieco płynu surowiczego, zmniejszającego tarcie między w o r k i e m osierdziowym i sercem. Wnętrze serca jest podzielone przegrodą serca (s eptum cordis) na połowy prawą i lewą. Każda z tych połów jest podzielona poprzeczną przegrodą na dwie części - górną i dolną. W ten sposób tworzą się cztery jamy serca: u jego podstawy leżą dwa przedsionki - p r a w y i l e w y ( a t r i u m dextrum etsinistrum), zaś poniżej przegrody poprzecznej, zwanej przegrodą przedsionkowo-komorową (s eptum atrioventriculare), leżą dwie k o m o r y serca - prawa i lewa (ventriculus dexter et sinister). Z podstawy serca wychodzą dwa duże naczynia, zwane tętnicami, tętnica główna, czyli aorta, i pień płucny, dzielący się na dwie tętnice płucne. Aorta po krótkim przebiegu ku górze zatacza łuk i oddając szereg gałęzi biegnie ku dołowi przy kręgosłupie. W jamie brzusznej kończy się podziałem na dwie tętnice biodrowe wspólne, przechodzące do kończyn dolnych. Wszystkie gałęzie aorty dzielą się na coraz drobniejsze tętniczki, które wreszcie przechodzą w naczynia w ł o s o w a t e o średnicy 7 - 8 (xm. Naczynia te przenikają do wszystkich narządów, doprowadzając do nich tlen i pożywienie. W t y c h naczyniach prąd krwi zostaje bardzo zwolniony, co umożliwia w y m i a n ę gazów i produktów przemiany materii między krwią i tkankami. Po spełnieniu swych zadań krew zbiera się w większe naczynia, zwane żyłami (venae), prowadzące ją w kierunku serca. Z żył kończyn dolnych i tułowia powstaje duża żyła g ł ó w n a dolna (vena cava interior), z kończyn górnych głowy i szyi - żyła główna górna (vena cava superior). Obie te żyły uchodzą do przedsionka prawego. Opisana droga krwi nosi nazwę krążenia albo krwiobiegu wielkiego. Po wypełnieniu się przedsionka prawego następuje jego skurcz (jednoczesny z lewym) i krew przepływa do komory przez otwór przedsionkowo-komorowy prawy. Jest on zaopatrzony w zastawkę tej samej nazwy, złożoną z 3 - 7 płatków. Dawniej nie rozróżniano w tej zastawce płatków dodatkowych i nazywano ją zastawką trójdzielną ( valva tricuspidalis), a ta tradycyjna nazwa dotrwała do dziś. Zastawka ta może otwierać się tylko w stronę komory. Kiedy po skurczu przedsionków kurczą się komory, to zastawka zamyka się i nie pozwala na cofnięcie się krwi do przedsionka, natomiast cała krew zostaje wyrzucona do pnia płucnego {truncus pulmonalis) oraz jego rozgałęzień - tętnic płucnych i zostaje skierowana do płuc. Tętnice płucne ( arteriae pulmonales) są jedynymi tętnicami, prowadzącymi krew żylną, to znaczy odtlenioną, bogatą w C0 2 . W płucach oddaje ona dwutlenek węgla i ulega natlenieniu, a więc zostaje przygotowana do ponow47
nego skierowania do tkanek. Płynie w i ę c ona z płuc do serca czterema (czasem trzema) żyłami płucnymi (venae pulmonales), które prowadzą krew tętniczą* natlenioną i uchodzą do przedsionka lewego. W przedsionku t y m kończy się krążenie płucne, czyli krwiobieg mały. Z przedsionka lewego krew przepływa o t w o r e m przedsionkowo-komorow y m l e w y m do komory lewej. Otwór ten zasłania zastawka dwudzielna (valva bicuspidaliś), która działa na zasadach podanych wyżej. Przy skurczu k o m o r y lewej krew zostaje wyrzucona do aorty, o czym już była m o w a wyżej. W krążeniu wielkim krew odbywa dużą drogę, na której musi przezwyciężyć znaczne opory. Dlatego komora lewa ma ściany prawie trzykrotnie grubsze od prawej, która wypełnia znacznie lżejszą pracę. Krążenie chłonki, czyli limfy (lympha), rozpoczyna się w przestrzeniach międzykomórkowych d r o b n y m i w ł o s o w a t y m i naczyniami chłonnymi (vasa lymphatica). Prowadzą one produkty przemiany materii, które komórki wydalają do przestrzeni międzykomórkowych. Na swej drodze naczynia przechodzą przez szeregi w ę z ł ó w chłonnych (nodi lymphatici), zbudowanych z tkanki łącznej siateczkowatej, w których są wytwarzane limfocyty. Prąd limfy ulega w nich zwolnieniu i następuje jej kontrola oraz ewentualna walka z zarazkami, trafiającymi do organizmu przez skórę oraz drogi pokarmowe, oddechowe czy moczowe. Drobne naczynia chłonne łącząc się tworzą większe pnie, które towarzyszą naczyniom krwionośnym. Naczynia chłonne, rozpoczynające się w błonieśluzowej przewodu jelitowego, zbierają się w większe pnie, uchodzące do węzłów chłonnych trzewnych, leżących na tylnej ścianie jamy brzusznej. Płynąca w nich chłonka ma kolor mleczny, spowodowany domieszką przechodzących do niej z jelita cienkiego zemulgowanych ciał tłuszczowych, skąd pochodzi jej nazwa - mlecz ( c h y l u s ) . Z węzłów trzewnych mlecz płynie do zbiornika mleczu ( cisterna chyli), od którego rozpoczyna się duże naczynie chłonne - p r z e w ó d piersiowy (ductus thoracicuś). Biegnie on przez śródpiersie i uchodzi do połączenia dużych pni żylnych, prowadzących krew z lewej strony głowy, szyi i kończyny górnej lewej. Jak wynika z powyższego, krew dochodzi do najdrobniejszych tętniczek dzięki pracy serca, które podtrzymuje ciśnienie w tętnicach. Ale w naczyniach włosowatych spada ono tak znacznie, że krew nie może dojść do serca, zwłaszcza z kończyn i tułowia. Dotyczy to również limfy. Do podnoszenia się krwi i limfy służą zastawki żylne i dużo liczniejsze od nich limfatyczne. Mają one kształt f a ł d ó w półksiężycowatych, zrdśniętych w y p u k ł y m brzegiem ze ścianą naczynia, a w o l n y m brzegiem skierowanym ku ujściu naczynia. Ponieważżyły i naczynia limfatyczne biegną w sąsiedztwie mięśni, bywają one w pewnych miejscach uciskane przez kurczące się mięśnie. Ucisk naczynia powoduje dzięki zastawkom przesunięcie zawartego w n i m płynu ku górze o jedno lub więcej pięter zastawek. Na większe pnie, uchodzące do serca, działa również tzw. ciśnienie ujemne, panujące w klatce piersiowej, o czym będzie mowa przy oddychaniu. 48
Układ moczowo-płciowy Układ moczowo-płciowy ( systema urogenitale) składa się z d w ó c h części: układu wydalniczego (moczowego) i układu rozrodczego (płciowego).
Układ m o c z o w y Jak już w i e m y z poprzedniego rozdziału, produkty przemiany materii trafiają do krwi. Zadaniem układu moczowego jest oddzielenie ich od krwi i wydalenie z organizmu. Pierwszą rolę spełniają nerki (renes). Leżą ond na tylnej ścianie jamy brzusznej w przestrzeni zaotrzewnowej, po obu stronach kręgosłupa. Są to duże narządy o kształcie zbliżonym do ziaren fasoli. Brzeg przyśrodkowy nerki jest wcięty i tworzy jej wnękę, do której wchodzi duża tętnica nerkowa ( a r t e r i a renalis), a wychodzi żyła nerkowa (vena renalis) i m o c z o w ó d (ureter), odchodzący od miedniczki n e r k o w e j (pe/v/s renalis), która leży w głębi nerki. Tętnica nerkowa oddaje szereg gałęzi, które dzielą się na drobniejsze tętniczki, tworzące tzw. kłębuszki nerkowe (glomerulirenales). Są to kłębki naczyń włosowatych, prowadzących krew tętniczą, otoczone torebkami nerkowymi, do których zostaje odsączany silnie rozcieńczony mocz p i e r w o t n y (urina). Mocz ten przechodzi.przez skomplikowany układ kanalików nerkowych, w których woda zostaje wchłonięta z powrotem do krwi, przez co następuje jego zagęszczenie. Zagęszczony mocz ostateczny zbiera się w miedniczce nerkowej, z której moczowód prowadzi go do zbiornika - pęcherza m o c z o w e g o ( vesica urinaria). Po zebraniu się w n i m pewnej ilości moczu (250-500 cm 3 ) jest on wydalany przez cewkę moczową (urethra). Pęcherz moczowy leży w miednicy mniejszej, do tyłu od spojenia łonowego. Jest on zbudowany z silnej, kurczliwej błony mięśniowej, wewnątrz-wysłany błoną śluzową, odporną na działanie moczu. Cewka moczowa żeńska jest krótkim (3-5 cm) przewodem, o średnicy około 7 m m , otwierającym się w zatoce moczowo-płciowej, natomiast męska jest znacznie dłuższa i węższa.
Układ p ł c i o w y Narządy płciowe męskie. Służą one do wytwarzania plemników i w p r o w a dzenia ich do narządów żeńskich. Plemniki są wytwarzane w jądrach (testes) i po procesie dojrzewania wyprowadzane do zbiorników, zwanych najądrzami. Z każdego najądrza (epididymis)prowadzi długi przewód,zwany nasieniow o d e m ( ductus deferens), który wchodzi do j a m y otrzewnej przez kanał pachwinowy i kieruje się w okolicę dna pęcherza moczowego. W tej okolicy leżą też dwa pęcherzyki nasienne (vesiculae seminales), które produkują ciecz, 4-Anatomia i fizjologia
49
wchodzącą w skład nasienia (sperma). Również pod dnem pęcherzyka leży pojedynczy gruczoł k r o k o w y (prostata), produkujący składnik nasienia. iMasie-' n i o w ó d każdej strony łączy się w obrębie gruczołu krokowego z przewodem pęcherzyka nasiennego, tworząc krótki w s p ó l n y p r z e w ó d w y t r y s k o w y (ductus ejaculatorius), uchodzący do cewki moczowej. Przewód ten jest czynny w czasie wytrysku nasienia (ejakulacji). W akcie płciowym do cewki moczowej w y p ł y w a również wydzielina gruczołu krokowego. Cewka moczowa wchodzi do narządu kopulacyjnego - prącia (penis) i otwiera się ujściem zewnętrznym na jego szczycie. Narządy płciowe żeńskie. Narządem wytwarzającym komórki płciowe żeńskie jest jajnik { o v a r i u m ) , położony w miednicy mniejszej, w jamie otrzewnej. Komórka jajowa (ovum) może być zapłodniona przez plemnik, co prowadzi do rozwoju zarodka. Zarodek może się rozwinąć wyłącznie w organizmie matczynym, który też ma szereg narządów, umożliwiających mu to zadanie. Zapłodniona komórka jajowa musi być przede wszystkim wyprowadzona z jamy otrzewnej do narządu, w którym ma się rozwijać płód. Narządem t y m jest macica (uterus), do której jajo wędruje drogą j a j o w o d u (tuba uterina). Macica jest silnym narządem m i ę ś n i o w y m , obficie unaczynionym, leżącym w miednicy mniejszej. Wewnątrz niej znajduje się jama macicy (cavum uteri), w której zarodek może się rozwijać, zaopatrywany drogą krwi matki, przepływającej przez umieszczony w jamie macicy narząd, zwany łożyskiem (p/acenta). Zarodek po d w ó c h miesiącach zaczyna przyjmować kształty ludzkie i od tego czasu staje się płodem (foetus). Po dziewięciu miesiącach ciąży następuje poród (partus) i płód zostaje w y d a l o n y z macicy na skutek jej skurczów i działania tłoczni brzusznej matki. Płód przechodzi przez p o c h w ę (vagina) i zostaje wydalony z łona matki, połączony z nią jedynie pępowiną, narządem łączącym łożysko z płodem. Po porodzie pępowina zostaje podwiązana w d w ó c h miejscach i przecięta, a urodzony płód, zwany noworodkiem, rozpoczyna własne życie pierwszym oddechem. Wkrótce po porodzie na skutek skuręzów macicy łożysko oddziela się od ściany macicy i zostaje wydalone.
Układ nerwowy Wszystkie czynności organizmu muszą być powiązane ze sobą i ułożone we w s p ó l n y m planie. Funkcje te spełnia układ n e r w o w y (systema nervosum), który również pełni czynności psychiczne i jest siedliskiem świadomości. Czynnością podstawową układu nerwowego jest odruch, czyli proces odbywający się bez udziału świadomości, polegający na odebraniu podniety ze świata zewnętrznego i natychmiastowej reakcji, jak np. cofnięcie ręki przy dotknięciu gorącego przedmiotu, czy zamknięcie powiek przy zagrożeniu oka. Odruchy uświadamiamy sobie dopiero po ich zakończeniu, ale większość 50
w ogóle nie jest notowana w naszej świadomości, jak np. oddychanie, praca narządów wewnętrznych itp. Pewne podobieństwo odruchów mają praksje, a więc czynności wyuczone, zautomatyzowane, zachodzące również bez udziału świadomości, jak np. jazda na rowerze. Układ n e r w o w y dzielimy na ośrodkowy, czyli centralny, i o b w o d o w y . Do ośrodkowego należy m ó z g o w i e ( encephalon) i rdzeń kręgowy {medulla spina lis). Mózgowie składa się z pnia mózgu (truncus cerśbri), móżdżku (cerebellum) i kory m ó z g o w e j (cortex cerebri). Rdzeń zawiaduje odruchami prostymi, pień mózgu wraz z móżdżkiem zawiaduje odruchami złożonymi, wreszcie kora jest narządem wyższych czynności nerwowych i siedliskiem świadomości. Łącznikami między ośrodkowym układem n e r w o w y m i narządami są nerwy, utworzone z pęczków wypustek komórek nerwowych, które przewodzą podniety. Jedne z nich prowadzą podniety uświadamiane i nie uświadamiane z obwodu, a więc z narządów, do układu nerwowego ośrodkowego. Są to nerwy czuciowe. Inne prowadzą podniety w o d w r o t n y m kierunku: z mózgowia i rdzenia kręgowego do mięśni - są to nerwy ruchowe. Jeszcze inne, przenoszące z układu ośrodkowego podniety przez nas nie uświadamiane do narządów wewnętrznych, należą do układu nerwowego autonomicznego (systema nervosum autonomicum). Nazwą tą obejmujemy integralną część układu nerwowego, zawiadującą czynnościami narządów wewnętrznych, do których należą mięsień sercowy, mięśniówka gładka, wchodząca w skład naczyń i innych narządów, oraz gruczoły. W skład n e r w ó w wchodzą najczęściej jednocześnie włókna czuciowe, ruchowe i autonomiczne. Takie nerwy nazywamy mieszanymi. W i e m y już, że układ nerwowy o b w o d o w y tworzą nerwy, które odchodzą od mózgowia i rdzenia kręgowego. M a m y 12 par n e r w ó w czaszkowych i 31 par rdzeniowych, tworzących liczne rozgałęzienia, sploty i zespolenia, które będą omówione w anatomii szczegółowej.
Układ gruczołów do krewnych Gruczoły ( glandulae) są to narządy powstałe przeważnie z nabłonka, posiadającego zdolności wydzielnicze. Liczba gruczołów w organizmie jest bardzo duża, najwięcej jest ich w układzie trawiennym. Gruczoły z reguły wyprowadzają swą wydzielinę drogą przewodów - są to gruczoły wydzielania zewnętrznego. Niektóre jednak nie mają przewodów i wydzielają swe produkty bezpośrednio do krwi. Są to gruczoły bezprzewodowe (glandulae sine ductibus). Są one obficie unaczynione i wydzielina ich zostaje wchłonięta przez naczynia na drodze osmozy. Dlatego zostały one nazwane dokrewnymi. Gruczoły dokrewne wydzielają ciała zwane hormonami. Należą one, podobnie jak w i t a m i n y , do biokatalizatorów, których małe ilości wystarczają do przyspieszenia szeregu reakcji biochemicznych, mających zasadnicze znaczenie dla życia organizmu. Brak ich może spowodować ciężkie zaburzenia w jego 51
czynnościach, dlatego też schorzenia gruczołów dokrewnych nieraz mogą decydować o życiu chorego. Do gruczołów dokrewnych należą: przysadka, gruczoł tarczowy (tarczyca), gruczoły przytarczyczne, aparat wysepkowy trzustki, gruczoły nadnerczowe (nadnercza), gruczoły płciowe (jądra i jajniki), grasica, szyszynka. Przysadka ( h y p o p h y s i s ) spełnia rolę nadrzędną w ś r ó d gruczołów dokrewnych, wpływających na ich wydzielanie. Leży ona w jamie czaszki, na powierzchni dolnej mózgu. Poza t y m h o r m o n y przysadki wpływają na zasadnicze czynności organizmu, jak wzrastanie, rozwój osobnika itd. Gruczoł tarczowy (glandula thyroidea) leży na szyi i jest największym gruczołem dokrewnym, zresztą o znacznych wahaniach w y m i a r ó w i ciężaru. Hormony tarczycy wpływają na przemianę materii, dojrzewanie i szereg innych czynności organizmu. Gruczoły przytarczyczne górne i dolne ( glandulae parathyroideae), w liczbie 3 - 4 , leżą na tylnej powierzchni tarczycy i mają w p ł y w na gospodarkę wapniową organizmu. Aparat w y s e p k o w y trzustki jest założony w samym ciele tego gruczołu w postaci wysepek słabo barwiących się komórek. Wydziela insulinę i glukagon, h o r m o n y regulujące gospodarkę cukrową. Nadnercze {glandula suprarenaliś), gruczoł parzysty, leży w jamie brzusznej, na górnych biegunach nerek. Wydziela około trzydziestu h o r m o n ó w , regulujących ważne czynności życiowe. Między innymi wytwarza adrenalinę, h o r m o n wpływający na układ krążenia. Gruczoły płciowe - jądro (testis) i jajnik (ovarium) - poza wytwarzaniem komórek płciowych, są ważnymi gruczołami dokrewnymi, powodującymi dojrzewanie płciowe organizmów oraz występowanie drugorzędnych cech płciowych, jak np. zarostu na twarzy u mężczyzn czy budowy miednicy u kobiet. Jajniki wydzielają również h o r m o n y regulujące przebieg ciąży. Grasica ( t h y m u s ) jest gruczołem wieku dziecięcego, na ogół czynnym do okresu pokwitania i potem stopniowo zanikającym. Co do czynności grasicy brak dostatecznych wyjaśnień. Przypuszczalnie pobudza ona wzrastanie i chroni organizm przed zakażeniem i zatruciem. Szyszynka {corpus pineale) jest t w o r e m wielkości małego ziarnka fasoli, leżącym w jamie czaszki pod płatem spoidła wielkiego mózgu. Zalicza się do gruczołów dokrewnych, chociaż o jej czynnościach brak pewnych wiadomości. Prawdopodobnie hamuje ona dojrzewanie, broniąc organizm dziecięcy przed niewłaściwym rozwojem czynności narządów płciowych.
UKŁAD NARZĄDÓW RUCHU
W przyrodzie spotyka się różne rodzaje sił powodujących ruch. Ruch mechaniczny powstaje wskutek działania sił grawitacji, pola magnetycznego i elektrycznego, różnicy ciśnień itp. W biologii najczęściej spotykany jest ruch powodowany siłą kurczących się mięśni, tzw. ruch miogenny. U kręgowców siła mięśni przenoszona jest na układ szkieletowy, który składa się z kości połączonych ruchomo stawami. W skład układu narządów ruchu wchodzą zatem: 1) układ kostny, 2) układ stawowo-więzadłowy, 3) układ mięśniowy. Układ kostny i s t a w o w o - w i ę z a d ł o w y stanowią' bierną część układu narządów ruchu, podczas gdy układ mięśniowy jest jego częścią czynną. UKŁAD KOSTNY - OSTEOLOGIA
Dane ogólne W skład szkieletu człowieka dorosłego wchodzi 206 oddzielnych kości. Liczba ta jednak jest zmienna: u dziecka 15-letniego jest większa (356), u starców mniejsza. Różnica wynika ze zrastania się poszczególnych części kości u ludzi młodych i ze zrastania się niektórych kości u osób w wieku podeszłym. Kość zbudowana jest z dwóch podstawowych składników: organicznego, tzw. oseiny, nadającej kości elastyczność^ i mineralnego, głównie śoli kwasu fosforowego i w ę g l o w e g o (p. Tkanka kostna). Fosforan wapnia i magnezu oraz węglan wapnia nadają kości odpowiednią twardość. Oseinę Tnożna wydzielić poddając kość działaniu silnych kwasów, które usuwają z niej sole mineralne. Odwapniona kość zachowuje kształt, jest bardzo sprężysta i łatwo odkształca Ina. Poddanie kości działaniu wysokiej temperatury usuwa z niej elementy organiczne (oseinę). Taka kość jest bardzo krucha i rozsypuje się nawet przy stosunkowo słabym uderzeniu. Wzajemny stosunek ilościowy i jakościowy oseiny i składnika mineralnego warunkuje właściwości mechaniczne kości. Kości u dzieci, bogate w składniki organiczne, są elastyczne i odporne nazłamania. Kości ludzi w wieku starszym są mało elastyczne i kruche ze względu na przewagę w nich składników mineralnych. Pod względem budowy wyróżnia się istotę k o s t n ą zbitą ( substantia compacta) i istotę gąbczastą (substantia spongiosa). 53
Z kości zbitej zbudowane są trzony kości długich i powierzchowne warstwy kości płaskich (sklepienie czaszki, mostek, łopatka, kości biodrowe). Istota gąbczasta wchodzi w skład końców kości długich, znajduje się między blaszkami zbitymi kości płaskich, jest również tworzywem większej części kości krótkich i różnokształtnych (kości nadgarstka i stępu). Wyróżnia się: 1) kości długie, 2) kości płaskie i 3) kości krótkie różnokształtne. ' Kości długie, których większość wchodzi w skład szkieletu kończyn, zbudowane są z trzonu {corpus) i położonych na jego końcach zgrubiałych nasad {epiphyses). W wieku dziecięcym nasada oddzielona jest od trzonu warstwą chrząstki nasadowej {cartilago epiphysialis). Chrząstka ta jest miejscem wzrostu kości na długość. Jak już wspomniano, zanika ona po ukończeniu wzrostu i wtedy nasada łączy się bezpośrednio z trzonem. Część trzonu sąsiadującą z chrząstką nasadową nazywa się przynasadą ( m e t a p h y s i s ) . Nasady kości długich pokryte są w całości lub w części chrząstką s t a w o w ą (cartilago articularis). Chrząstka stawowa umożliwia swobodne ruchy w stawie, a ponadto jest miejscem wzrostu nasad. Budowę kości długiej można zatem przedstawić następująco: Chrząstka stawowa Nasada blizsza Chrząstką nasadowi" Pi/yruisif i
HHHHHHHHb f"/,ii.i^id i
< I " / Iitk.l lid . i ' i r t y 1
Nasada dalsza Chrząstką stawowa Nasady zbudowane są przede wszystkim z istoty gąbczastej, trzony z istoty kostnej zbitej. W obrębie przynasad w miarę zbliżania się do nasady warstwa istoty zbitej staje się cieńsza, a masa istoty gąbczastej powiększa się. Do kości płaskich zalicza się kości sklepienia czaszki, łopatkę, mostek i kości biodrowe. Mają one kształt cieńszych lub grubszych, najczęściej powyginanych, płyt. Zewnętrzna i wewnętrzna warstwa kości płaskiej zbudowana jest z tkanki kostnej zbitej. Między nimi leży śródkoście ( d i p l o e ) zbudowane z substancji gąbczastej. Kości płaskie w m ł o d y m wieku mają niekiedy dodatkowe punkty kostnienia umieszczone najczęściej na obwodzie kości. W takich przypadkach oddzielone są one od właściwej kości płaskiej chrząstką nasadową. Kości krótkie i różnokształtne trudno zaliczyć do jednej z w y m i e n i o n y c h wyżej grup. Mają one najczęściej kształt nieregularnej bryły. Na ich powierzch54
niach występują dodatkowe guzki, bruzdy i wyrostki. Kości krótkie i różnokształtne w swojej masie zbudowane są z tkanki gąbczastej, którą pokrywa cienka warstwa istoty kostnej zbitej. Kształt kości jest uwarunkowany czynnością, którą one spełniają. Budowa kości, czyli jej architektonika, opiera się na zasadzie największej wytrzymałości przy najmniejszej ilości budulca. Stąd też trzony kości długich mają kształt rur wytrzymałych na obciążenie osiowe i zginanie. Nasady kości długich, kości różnokształtne i śródkoście kości gąbczastej zbudowane są z tkanki gąbczastej, której układ beleczek pozwala na przenoszenie odpowiednich dla danej kości sił ściskających i rozciągających (ryc. 25). Nawet już po ukształtowaniu się kośćca zmiana w a r u n k ó w mechanicznych (np. po złamaniu) powoduje zmianę architektoniki kości w kierunku dostosowania do nowych warunków. Układ beleczek kostnych zgodny jest zatem z przebiegiem linii sił tworząc tzw. trajektorie. W miejscach obciążeń beleczki występują obficiej i są grubsze. W polach mechanicznie martwych jest ich mniej i są cieńsze.
Ryc. 25. Schemat przebiegu beleczek kostnych w górnej nasadzie kości udowej.
Okostna Powierzchnię kości pokrywa błona zwana okostną (periosteum). Zbudowana jest ona z dwóch warstw. Zewnętrzna, bogato unaczyniona, warstwa utworzona jest z tkanki łącznej. Wewnętrzna, przylegająca bezpośrednio do kości, warstwa okostnej zawiera liczne elementy komórkowe, z których komórki kościotwórcze (osteoblasty) biorą udział w budowie i odtwarzaniu kości. Przechodzące z okostnej do powierzchownych warstw kości włókna klejodajne przytwierdzają okostną (włókna Sharpeya). Wraz z nimi wnikają do kości drobne naczynia krwionośne unaczyniające powierzchowną jej warstwę. 55
Części chrzęstne szkieletu pokrywa błona, zwana ochrzęstną ( peńchondrium). Zbudowana jest ona podobnie do okostnej. Od strony j a m y szpikowej kość pokrywa błona śródkostna (endosteum). Oddziela ona kość od szpiku kostnego. Szpik kostny ( m e d u l l a ossium), wypełniający j a m y szpikowe kości długich, ma u osób dorosłych zabarwienie żółtawe, stąd jego nazwa - szpik żółty. Barwa szpiku zależy od dużej ilości tkanki tłuszczowej, z której przede wszystkim jest zbudowany. Szpik kostny wypełnia również przestrzenie międzybeleczkowe tkanki kostnej gąbczastej. U n o w o r o d k ó w występuje wyłącznie szpik czerw o n y , w którym wytwarzane są krwinki czerwone. W miarę wzrostu organizmu szpik czerwony przemienia się w szpik żółty. Szpik czerwony pozostaje jedynie w niektórych kościach (kręgi, mostek, żebra i łopatka).
Czynność kości Kość jest żywą tkanką biorącą udział w przemianie materii. Jest ona ponadto największym zbiornikiem soli mineralnych, przede wszystkim soli wapnia. Wapń pobierany jest z przewodu pokarmowego i wydalany w większości przez Ca Szkielet 1000 g Tkanki 1 g
400 mg 400 mg
200 mg
Ryc. 26. Krążenie wapnia w ustroju.
56
nerki. Istnieje ciągła wymiana wapnia między kością a płynami tkankowymi w ilości ok. 400 mg na dobę (ryc. 26). Żywy udział w przemianie wapniowej biorą komórki. Komórki kościotwórcze (osteoblasty) mają zdolność odkładania wapnia w kości. W przeciwieństwie do nich osteoklasty zawierają enzymy wypłukujące wapń, który przechodzi do płynu międzykomórkowego. Prawidłową przemianę wapniową regulują hormony przytarczyc i kory nadnercza. Dla sprawnego przebiegu procesów niezbędna jest witamina D.
Rozwój i wzrost kości Tkanka kostna może rozwinąć się albo bezpośrednio na podłożu tkanki łącznej zarodkowej (mezenchymy), albo też za pośrednictwem chrząstki. -Mezenchyma • Chrząstka — Kość
Kość
Na podłożu łącznotkankowym rozwija się obojczyk, kości twarzy i sklepienia czaszki. Wszystkie inne kości powstają na podłożu chrzęstnym. Kostnienie na podłożu chrzęstnym może rozpoczynać się od powierzchni chrząstki (kostnienie odchrzęstne). Z chwilą wytworzenia się pierwszych warstw kości Nasada Przynasada-
Chrząstka nasadowa
Jądro kostnienia
— Trzon
Ryc. 27. Schemat rozwoju kości długich. Kolorem żółtym oznaczono kość, kolorem niebieskim chrząstkę. .
57
ochrzęstna zmienia się w okostną i dalej postępujące kostnienie nazywa się odokostnowym. Kostnienie rozpoczynające się w głębi chrząstki nazywamy śródchrzęstnym. W ten sposób rozwijają się jądra kostnienia nasad kości długich i kości krótkie. Przetrwała po skostnieniu nasady i granicząca z trzonem warstwa chrząstki nosi nazwę chrząstki nasadowej. Chrząstka nasadowa znika po zakończeniu wzrostu kości. Chrząstka nasadowa jest miejscem wzrostu kości na długość. Komórki chrzestne leżące w sąsiedztwie nasady rozmnażają się i przesuwając się w kierunku trzonu ulegają licznym zmianom, których w y n i kiem jest powstanie nowej kości. Zniszczenie chrząstki nasadowej zatrzymuje wzrost trzonu kości długiej. Rolę chrząstki wzrostowej nasad pełni chrząstka stawowa. Jej zniszczenie w okresie wzrostu organizmu powoduje zatrzymanie lub upośledzenie wzros tu nasady. Okostną umożliwia wzrost kości na grubość. Kości mają również zdolność regeneracji. Złamana kość, jeśli jest odpowiednio unieruchomiona, zrasta się. Zrost o d ł a m ó w przebiega w kilku fazach (ryc. 28). Podczas złamania dochodzi do w y l e w u krwi w sąsiedztwie odłamów. Krew ta krzepnie. Do skrzepu wrastają z otoczenia i jamy szpikowej drobne naczynia i dochodzi do rozplemu komórek. W przestrzeniach międzykomórkowych wytrącają się sole mineralne. W ten sposób powstaje kostnina pierwotna.
Łd®
i B
Ryc. 28. Fazy zrastania się złamanej kości: A - w y l a n a do szczeliny złamania krew, 6- rozwój ziarniny, C - powstanie tkanki kostnawej, D - blizna kostna.
W następnej fazie komórki kościogubne (osteoklasty) usuwają zwapniałą tkankę kostniny, a jednocześnie komórki kościotwórcze odbudowują kość. Pozostałością po złamaniu jest blizna kostna (callus). Kości mają zdolność dostosowywania swego kształtu do działających na nie sił. Właściwość ta jest szczególnie wyraźna u dzieci. Złamanie, w którym odłamy ustawiają się pod kątem, może wygoić się bez zniekształcenia (ryc. 29). 58
Ryc. 29. Modelowanie kształtu kości po zrośnięciu się odłamów w nieprawidłowym ustawieniu.
Osteologia szczegółowa W rozdziale o m ó w i m y kolejno: 1) kości kręgosłupa i klatki piersiowej, 2) kości kończyny górnej, 3) kości kończyny dolnej oraz 4) kości czaszki.
Kręgosłup Kręgosłup (columna vertebralis) składa się z kości, zwanych kręgami (vertebrae). Kręgi łącząc się ze sobą tworzą elastyczny słup będący osią tułowia. Górny koniec kręgosłupa podpiera czaszkę, dolny łączy się z kośćmi miednicy. Wewnątrz kręgosłupa leży rdzeń kręgowy. Kręgosłup składa się z 33-34 kręgów i dzieli się na pięć odcinków (tab. 1). T y p o w y kręg składa się z trzonu, łuku oraz siedmiu wyrostków. Masywnie zbudowany trzon kręgu zwrócony jest ku przodowi. Do jego tylnej powierzchni przylega łuk, który zamyka od tyłu otwór kręgowy, będący odcinkiem kanału kręgowego. Z łukiem łączą się dwie pary w y r o s t k ó w s t a w o w y c h służących do połączenia z sąsiednimi kręgami i stąd znajdują się na nich powierzchnie stawowe. Symetrycznie położone w y r o s t k i poprzeczne służą między innymi jako miejsce przyczepu mięśni. Wyrostek kolczysty przymocowuje się na 59
tylnej części łuku i tworzy mocną dźwignię dla mięśni. Kształt kręgów zmienia się w zależności od odcinka kręgosłupa, z którego pochodzą. Kręgi szyjne mają mały i spłaszczony w kierunku strzałkowym trzon. Wyrostki stawowe są niskie, ich powierzchnie stawowe ustawione skośnie w stosunku do płaszczyzny poziomej. Wyrostki poprzeczne, rozdwojone na
60
Tabela 1 Odcinki kręgosłupa Odcinek
Liczba kręgów 7 12 5 5 4-5
Szyjny Piersiowy Lędźwiowy Krzyżowy Guziczny
Wyrostek kolczysty
• Trzon
Otwór wyrostka
poprzecznego Wyrostki stawowe górne
,a/ 1 * i Wyrostek stawowy dolny
Wyrostek poprzeczny
Ryc. 31. Kręg szyjny.
końcach, przebite są pionowo o t w o r e m w y r o s t k a poprzecznego, przez który przechodzi tętnica kręgowa. Otwór kręgowy ma kształt trójkątny. Pierwszy kręg szyjny nosi nazwę kręgu szczytowego (atlas). Nie ma on trzonu, lecz składa się z dwóch łuków: przedniego i tylnego. Miejsca ich połączenia są zgrubiałe i noszą nazwę części bocznych. Na tylnej powierzchni łuku przedniego leży dołek zębowy służący do połączenia z zębem kręgu obrotowego. Na częściach bocznych leżą dołki s t a w o w e górne do połączenia z kłykciami kości potylicznej i dolne, przylegające do odpowiednich powierzchni stawowych kręgu obrotowego. Kręg o b r o t o w y (axis) jest d r u g i m kręgiem szyjnym. Wydłużony ku górze trzon tworzy ząb kręgu obrotowego. Przednia powierzchnia stawowa zęba przylega do dołka zębowego kręgu szczytowego, powierzchnia tylna - do więzadła poprzecznego. 61
Ostatni siódmy kręg szyjny ma długi wyrostek kolczysty i stąd został nazwany kręgiem w y s t a j ą c y m ( v e r t e b r a promineris). Kręgi piersiowe są większe od kręgów szyjnych. Trzony kręgowe są płaskie, ich w y m i a r strzałkowy jest większy od poprzecznego. Wyrostki stawowe są długie, ich powierzchnie stawowe ustawione w płaszczyźnie zbliżonej do czołowej. Długie i pochylone ku dołowi wyrostki kolczyste zachodzą na siebie dachówkowato. Na zgrubiałym końcu długich w y r o s t k ó w poprzecznych leży dołek ż e b r o w y wyrostka poprzecznego, służący do połączenia się z guzkiem żebra. Na bocznych powierzchniach trzonów w sąsiedztwie krawędzi znajdują się dołki żebrowe. Dołki dwóch sąsiadujących kręgów tworzą powierzchnie stawowe, służące do połączenia z g ł o w a m i żeber. Kręgi lędźwiowe, największe, mają trzony nerkowatego kształtu. Ich w y m i a r poprzeczny jest większy od strzałkowego. Duże wyrostki stawowe ustawione są strzałkowo. Wyrostki poprzeczne cienkie i długie są właściwie szczątkowymi żebrami, stąd też noszą one nazwę w y r o s t k ó w żebrowych. Wyrostki kolczyste kręgów lędźwiowych są wysokie, silnie rozwinięte i ustawione poziomo w płaszczyźnie strzałkowej.
A
B Ryc. 32. Kręg piersiowy widziany od góry (A) i od strony prawej (S).
62
Wyrostek żebrowy. Wyrostek kolczysty
Ryc. 33. Kręg lędźwiowy.
Kość krzyżowa (os sacrum) powstała ze zrośnięcia się ze sobą pięciu kręgów krzyżowych. Ma kształt zwężającego się ku dołowi graniastosłupa. Wyróżniamy w niej podstawę zwróconą ku górze, oraz wierzchołek, zwrócony ku t y ł o w i i dołowi. Od przodu znajduje się gładka powierzchnia miedniczna, grzbietowa powierzchnia kości krzyżowej jest nierówna. Na powierzchni miednicznej widoczne są cztery pary o t w o r ó w krzyżowych miednicznych, które łączą się z kanałem krzyżowym. Na nierównej powierzchni grzbietowej znajdują się grzebienie, powstające ze zrośnięcia się odpowiednich w y r o s t k ó w kręgowych. Grzebień p o ś r o d k o w y powstał ze zrośniętych wyrostków kolczystych. Wyrostki stawowe utworzyły grzebienie krzyżowe pośrednie, a wyrostki poprzeczne grzebienie boczne. Między grzebieniami pośrednim i bocznym leżą o t w o r y krzyżowe grzbietowe. Wzdłuż kości krzyżowej biegnie kanał krzyżowy - przedłużenie kanału kręgowego. Kanał kończy się rozworem krzyżowym. Na masywnie zbudowanych częściach bocznych kości krzyżowej leżą powierzchnie uchowate, łączące się z p o d o b n y m i powierzchniami kości miednicznej. Kość guziczna (os coccygis) ma u człowieka charakter szczątkowy. W skład niej wchodzi 4 lub 5 kręgów, mających postać trójkątnych lub okrągłych kostek. Podstawa kości guzicznej łączy się z kością krzyżową. 63
Szkielet klatki piersiowej Szkielet klatki piersiowej składa się z 12 kręgów piersiowych, 12 par żeber i mostka. Żebra (costae) mają postać wygiętych półksiężycowato blaszek kostnych. Każde z nich ma koniec przedni, czyli mostkowy, trzon i koniec t y l n y - k r ę g o s ł u powy. Koniec przedni kości żebrowej łączy się z chrzęstnym przedłużeniem żebra, zwanym chrząstką żebrową. Zgrubiały koniec tylny żebra nosi nazwę g ł o w y żebra. Nieco bocznie od niej leży guzek żebra. Głowa żebra łączy się z dołkami ż e b r o w y m i leżącymi na krawędziach trzonów kręgowych. Guzek żebra jest miejscem połączenia stawowego z dołkiem żebrowym wyrostka poprzecznego. Trzon żebra ma wypukłą powierzchnię zewnętrzną i wklęsłą wewnętrzną. Przy d o l n y m , ostrym brzegu żebra znajduje się płytka bruzda żebra, w której biegną naczynia i nerwy międzyżebrowe. W zależności od sposobu połączenia żeber z mostkiem dzielą się one na: 1) żebra p r a w d z i w e (I—VII), dochodzące bezpośrednio do mostka i łączące się z nim chrząstką żebrową; 2) żebra rzekome (Vlll-X), łączące się z mostkiem pośrednio poprzez chrząstkę żebra położonego wyżej; 3) żebra w o l n e (XI i XII), nie mające połączenia z mostkiem, leżące swobodnie w ścianie brzucha. Mostek ( sternum) należy do nieparzystych kości ustroju. Ma on kształt płaskiej kości ograniczającej od przodu klatkę piersiową. Rozróżniamy rozszerzoną część górną - rękojeść mostka, wydłużoną część środkową - trzon mostka i małą zwężającą się część dolną, zwaną w y r o s t k i e m mieczykowatym. Rękojeść łączy się z trzonem pod kątem rozwartym od tyłu, nazwanym kątem mostka. Na górnej krawędzi rękojeści znajduje się wcięcie szyjne. Po obu jego stronach leżą wcięcia obojczykowe łączące się z obojczykami. Na bocznych krawędziach rękojeści mostka leżą wcięcia żebrowe żeber I i II. Na krawędziach trzonu mostka znajdują się wcięcia dla żeber od III do VI. Wcięcie dla żebra VII znajduje się na granicy między trzonem a wyrostkiem mieczykowatym.
Ryc. 34. Połączenie żeber z kręgiem i mostkiem.
64
Wciocii
Pierwszy kręg piersiowy
mn-.ik-
Rękojeść mostka
obojczykow
Trzon - iV!sśślk, mostka
Wyrostek mieczykowaty
Żebra rzekome
Żebra wolne Ryc. 35. Szkielet klatki piersiowej.
Klatka piersiowa { t h o r a x ) jako całość ma kształt ściętego stożka, spłaszczonego od przodu ku tyłowi. Rozróżnia się o t w ó r górny klatki piersiowej oraz znacznie większy o t w ó r dolny. Otwór górny ograniczają trzon I kręgu piersiowego, pierwsze żebra i górny brzeg rękojeści mostka. Płaszczyzna otworu jest pochylona do przodu. Dolny otwór klatki piersiowej ograniczają trzon XII kręgu piersiowego, XII żebra, końce żeber XI, chrząstki żeber rzekomych i wyrostek mieczykowaty mostka. Płaszczyzna otworu dolnego klatki piersiowej pochylona jest ku tyłowi. Ścianę klatki piersiowej tworzą żebra, mostek i odcinek piersiowy kręgosłupa.
Kości kończyny górnej W skład szkieletu kończyny górnej wchodzi 37 kości, Dzielą się one na: 1) kości obręczy kończyny górnej, do których zalicza się obojczyk i łopatkę, oraz 2) kości kończyny górnej wolnej. Należą do nich: kość ramienna, 2 kości przedramienia, 8 kości nadgarstka, 5 kości śródręcza i 14 kości palców. 5 - Anatomia i fizjologia
1:
65
I
/ f— K. promieniowa
Kości obręczy kończyny górnej Obojczyk (c/av/cu/a) stanowi jedyne bezpośrednie połączenie między kośćcem kończyny górnej a szkieletem klatki piersiowej. Obojczyk łączy wyrostek barkowy łopatki z wcięciem obojczykowym mostka. Ma on kształt wydłużonej litery S. Koniec barkowy obojczyka jest spłaszczony od góry ku dołowi, a koniec mostkowy zgrubiały. Górna i przednia powierzchnia obojczyka jest gładka. Na powierzchni dolnej znajdują się nierówności, do których przymocowują się więzadła. Obojczyk jest dobrze widoczny i wyczuwalny przez skórę. 66
Brzeg górny
Wyrostek kruczy
Wcięcie łopatki
Powierzchnia , stawowa obojczyka -Wyrostek barkowy Wydrążenie stawowe
Dół nadgrzebieniowy
Grzebień łopatki , Brzeg kręgosłupowy
I
Dół podgrzebieniowy
:—Brzeg pachowy
Ryc. 37. Łopatka prawa.
Łopatka ( scapula) należy do kości płaskich. Ma ona kształt trój kąta. Dwa jego dłuższe boki tworzą zaokrąglony brzeg boczny i ostry brzeg przyśrodkowy. Na krótszym brzegu g ó r n y m znajduje się wcięcie łopatki. Bocznie od niego leży skierowany ku przodowi w y r o s t e k kruczy. Brzegi łopatki, łącząc się ze sobą, tworzą kąty: dolny, górny i boczny. Kąt boczny jest zgrubiały. Na nim znajduje się nieco wklęsła powierzchnia dla stawu ramiennego, zwana wydrążeniem s t a w o w y m . Łopatka ma dwie powierzchnie: przednia powierzchnia żebrowa, wklęsła, zwrócona jest ku żebrom, powierzchnię grzbietową łopatki przedziela wystający grzebień łopatki. Ponad mm leży dół nadgrzebieniowy, poniżej dół podgrzebieniowy. W przedłużeniu grzebienia łopatki znajduje się silnie rozwinięty w y r o s t e k barkowy. Na wyrostku t y m leży powierzchnia s t a w o w a dla połączenia z barkowym końcem obojczyka. Kości kończyny górnej w o l n e j Kość ramienna (humerus) należy do kości długich. Wyróżnia się w niej koniec górny, trzon i koniec dolny. Na końcu g ó r n y m znajduje się g ł o w a kości ramiennej. Głowę ogranicza szyjka anatomiczna kości ramiennej. Bocznie od szyjki leżą dwa guzki oddzielone od siebie bruzdą międzyguzkową. Ku t y ł o w i od niej leży guzek większy, ku przodowi - guzek mniejszy. Od g u z k ó w w kierunku trzonu ciągną się grzebienie g u z k ó w większego i mniejszego. Nieco poniżej guzków znajduje się tzw. szyjka chirurgiczna kości ramiennej, nazwana tak, ponieważ jest to miejsce częstych złamań. Trzon kości ramiennej, o przekroju zbliżonym do kolistego, na tylnej: powierzchni ma skośnie przebiegającą płytką bruzdę n e r w u promieniowegp.
Nierówność na bocznej powierzchni trzonu jest miejscem przyczepu mięśnia naramiennego i nosi nazwę guzowatości naramiennej. Koniec dalszy kości ramiennej jest spłaszczony w płaszczyźnie czołowej i ma dość złożoną budowę. Po obu stronach końca znajdują się wyniosłości, zwane nadkłykciami, z których przyśrodkowy jest znacznie dłuższy od bocznego. Pod nadkłykciem przyśrodkowym leży bruzda n e r w u łokciowego. Między nadkłykciem i nieco poniżej nich leży kłykieć kości ramiennej, na którym znajdują się powierzchnie stawowe dla połączenia z kośćmi przedramienia. Powierzchnia boczna służąca do połączenia się z kością promieniową leży na wyniosłości zwanej główką kości ramiennej. Przyśrodkowo leży bloczek łączący się z kością łokciową. Powyżej btoczka na tylnej powierzchni kości ramiennej leży głęboki dół wyrostka łokciowego, w który wsuwa sięwyróstek łokciowy kości łokciowej podczas prostowania przedramienia. Kość promieniowa ( radius) leży po bocznej stronie przedramienia. Górny jej koniec ukształtowany jest w postaci g ł o w y kości promieniowej. Na szczycie Bruzda międzyguzkowa
Głowa kości ramiennej
Guzek większy Szyjka— chiru™: k. rar guzka większego
v
Grzebień mniejszego
Trzon k. ramiennej
Nadkłykiec— borany
—-Nadkłykiec przyśrodkowy
k. ramiennej Ryc. 38. Kość ramienna widziana od przodu.
©8
Głowa < kości ^ kości ramiennej
x
^Szyjka anatomiczna — Guzek mniejszy ^Szyjka chirurgiczna k. ramiennej
Trzon—|k. ramiennej 1 ' .
.—Guzowatość ; „m;„ naramienna - Bruzda n. promieniowego
. Dół wyrostka łokciowego Nadkłykiecprzyśrodkowy
^ „
Bruzda n. łokciowego
|
.
f—Nadkłykiec boczny
Bloczek
Ryc. 39. Kość ramienna widziana od tyłu.
głowy znajduje się wklęsła powierzchnia stawowa dla połączenia z główką kości ramiennej, a dookoła głowy leży o b w ó d stawowy, który styka się zodpowiednim wcięciem kości łokciowej. Poniżej głowy znajduje się guzowatość kości promieniowej, będąca miejscem przyczepu mięśnia dwugłowego ramienia. Trzon kości promieniowej mą przekrój trójkątny. Jego przedni zgrubiały brzeg przechodzi ku dołowi w wyrostek rylcowaty. Szeroki, dolny koniec kości promieniowej ma powierzchnię stawową nadgarstka dla połączenia się z kośćmi nadgarstka. Na przyśrodkowej powierzchni dalszej nasady kości promieniowej znajduje się wcięcie łokciowe, służące do połączenia z głową kości łokciowej. Kość łokciowa (u/na) jest dłuższa od kości promieniowej i zwęża się stopniowo ku dołowi. Na końcu górnym znajduje się wcięcie bloczkowe. Ograniczają je od góry wyrostek łokciowy, a od dołu wyrostek dziobiasty. Na powierzchni bocznej u podstawy wyrostka dziobiastego leży wcięcie promieniowe, służące do połączenia z obwodem stawowym głowy kości promienio69
K. promieniowa
K łokciowa
Ryc. 40. Ułożenie kości przedramienia w odwróceniu.
Ryc. 41. Ułożenie kości przedramienia w nawróceniu.
Ryc. 42. Kości ręki.
wej. Nieco poniżej wyrostka dziobiastego znajduje się guzowatość kości łokciowej. Trzon kości, trójkątny w części górnej i środkowej, zwęża się i zaokrągla w części dolnej. Koniec dalszy kości łokciowej tworzy głowa kości łokciowej. Ma ona obrączkowatą powierzchnię stawową do połączenia z wcięciem ł o k c i o w y m kości promieniowej. Na przyśrodkowej powierzchni g ł o w y kości łokciowej leży krótki i cienki w y r o s t e k rylcowaty. Kości ręki (ossa manus) dzielą się na kości nadgarstka, śródręcza i kości palców. Osiem kości nadgarstka układa się w dwa szeregi: bliższy-sąsiadujący z kośćmi przedramienia, i szereg dalszy-łączącysięz kośćmi śródręcza. Ich ułożenie przedstawia tab. 2. Z kością promieniową łączą się trzy pierwsze kości szeregu bliższego. Łącznie tworzą one jajowatego kształtu powierzchnię stawową, wchodzącą w skład stawu promieniowo-nadgarstkowego. Dolne powierzchnie szeregu bliższego łączą się z kośćmi szeregu dalszego. Centralną kością nadgarstka jest kość główkowata. Dolne powierzchnie dalszego szeregu kości nadgarstka 71
łączą się z podstawami kości śródręcza. Dłoniowa powierzchnia nadgarstka jako całości jest wklęsła i tworzy bruzdę nadgarstka, w której leżą ścięgna zginaczy palców ręki. Do kości śródręcza należy pięć kości długich, które oznacza się kolejno, poczynając od kciuka. Kość śródręcza składa się z trzonu i dwóch końców. Koniec bliższy utworzony jest przez podstawę. Podstawa łączy się z kośćmi nadgarstka. Na końcu dalszym znajduje się głowa kości śródręcza, która łączy się z podstawą bliższego paliczka odpowiedniego palca. Kości palców ręki tworzą paliczki (phalanges). Palec I - kciuk, ma dwa paliczki, pozostałe palce mają ich po trzy. Paliczek sąsiadujący z kością śródręcza nazywa się bliższym, d r u g i - ś r o d k o w y m , a t r z e c i - d a l s z y m . Paliczki bliższe i środkowe mają podstawę, trzon i głowę. Paliczek dalszy jest spłaszczony i na jego końcu dalszym znajduje się półksiężycowatego kształtu guzowatość paliczka dalszego. Po stronie dłoniowej stawu śródręczno-palcowego I palca leżą dwie małe kosteczki, zwane trzeszczkami.
Tabela 2
Szereg bliższy Szereg dalszy
Strona promieniowa
Strona łokciowa
k. łódeczkowata, k. księżycowata k. czworoboczna, większa k. czworoboczna, mniejsza
k. trójgraniasta, k. grochowata k. główkowata, k. haczykowata
Kości kończyny dolnej W skład szkieletu kończyny dolnej wchodzi 31 kości, które dzielą się na kości obręczy kończyny dolnej i kości kończyny dolnej wolnej. K. krzyżowa -K. biodrowa
Kolec biodrowy przedni górny Kolec biodrowy przedni dolny
•Uff
• Fdnowka st;iwu biodrowego
Otwór zasłoniony
K. łonowa K kulbzowa Spojenie łonowe Ryc. 43. Miednica.
72
Kości obręczy kończyny dolnej Obręcz kończyny dolnej, w przeciwieństwie do obręczy kończyny górnej, zbudowana jest bardzo mocno i składa się z dwóch kości miednicznych i kości krzyżowej. Łączą się one ze sobą tworząc mocny pierścień kostny, zwany miednicą (pelvis). Kość miedniczna (os coxae) rozwija się z trzech oddzielnych kości, które w wieku dziecięcym oddziela warstwa chrząstki. Największa z nich, skierowana ku górze i ku bokowi, nazywa się kością biodrową (os ilium). Od strony dolnej i tylnej leży kość kulszowa (os ischii), od przodu i dołu - kość ł o n o w a (os pubis). W y m i e n i o n e kości łączą się w obrębie dużej i głębokiej panewki s t a w u biodrowego, służącej do połączenia z głową kości udowej. W dolnej części brzegu panewki widnieje wcięcie panewki. Poniżej panewki leży o t w ó r
Ryc. 44. Kości kończyny dolnej.
73
Grzebień biodrowy
Talerz biodrowej
Kolec biodrowy przedni górny
Kolec biodrowy tylny górny
Trzon k. biodrowej
Wcięcie kulszowe większe
Panewka Kolec kulszowy Wcięcie kulszowe
Otwór zasłoniony
Guz kulszowy-™
Ryc. 45. Kość miedniczna.
zasłoniony. Jego nazwa pochodzi od błony zasłonowej przymocowującej się do krawędzi otworu. Od tyłu i dołu otwór zasłoniony ograniczają trzon i gałąź kości kulszowej. W miejscu ich połączenia znajduje się guz kulszowy, a nieco powyżej niego kolec kulszowy. Kolec kulszowy oddziela wcięcie kulszowe mniejsze od leżącego powyżej głębokiego wcięcia kulszowego większego. Od przodu i dołu otwór zasłoniony ograniczają gałęzie kości łonowej górna i dolna. W miejscu połączenia obu gałęzi kości łonowej znajduje się powierzchnia spojenia służąca do połączenia się z taką samą powierzchnią drugiej kości miednicznej. Kość biodrowa, tworząca część kości miednicznej, zalicza się do kości płaskich. Bezpośrednio z panewką sąsiaduje trzon kości biodrowej, przechodzący następnie w talerz kości biodrowej. Powierzchnia wewnętrzna talerza biodrowego jest nieckowato zagłębiona i nosi nazwę dołu biodrowego. Dół ten ogranicza kresa łukowata. Na zewnętrznej powierzchni talerza kości biodrowej przyczepiają się mięśnie pośladkowe. Wyrazem tego są kresy pośladkowe. Od góry talerz kości biodrowej ogranicza grzebień biodrowy. Grzebień ten kończy się z przodu kolcem biodrowym przednim górnym, a od tyłu kolcem biodrowym tylnym górnym. Grzebień kości biodrowej i wymienione wyżej kolce są dobrze wyczuwalne przez skórę. Poniżej kolca biodrowego przedniego górnego leży kolec biodrowy przedni dolny. 74
Podobnie na tylnej części talerza kości biodrowej znajduje się kolec biodrowy t y l n y dolny. W jego sąsiedztwie na zgrubiałej przyśrodkowej części kości biodrowej widoczna jest powierzchnia uchowata, służąca do połączenia ztaką samą powierzchnią kości krzyżowej. Ku tyłowi od powierzchni uchowatej kość biodrowa jest nierówna i tworzy w t y m miejscu guzowatość biodrową, która jest miejscem przyczepu mocnych więzadeł łączących kość biodrową z kością krzyżową. Kości kończyny dolnej w o l n e j Kość u d o w a ( f e m u r ) jest największą kością ustroju. Składa się z końca bliższego, trzonu i końca dalszego. Na końcu bliższym znajduje się głowa kości u d o w e j {caputfemoris), o kształcie zbliżonym do kuli. Na jej szczycie leży dołek g ł o w y kości udowej. Głowa łączy się z szyjką kości udowej. Na granicy szyjki i trzonu kości udowej od przodu leży kresa międzykrętarzowa, od tyłu granicę tę stanowi grzebień międzykrętarzowy. Linia i grzebień łączą ze sobą dwie duże wyniosłości, zwane krętarzami. Krętarz większy leży w przedłużeniu trzonu, a jego szczyt znajduje się na wysokości środka głowy kości udowej. Krętarz mniejszy leży poniżej i nieco ku tyłowi od dolnej powierzchni szyjki kości udowej. Głowa k. udowej
Kresa 11 ńędzykrętarzowa Krętarz mniejszy
! i
1 1 Trzon
Nadkłykiec b o c z n y _ Kłykieć boczny
i
Nadkłykiec przyśrodkowy Kłykieć przyśrodkowy Powierzchnia rzepkowa
Ryc, 46. Prawa kość udowa od przodu.
75
Grzebień międzykrętarzowy
• -Kresa chropawa
-Dół międzykłykciowy Ryc. 47. Prawa kość udowa od tyłu.
Rurowaty trzon kości udowej jest nieco wygięty ku przodowi. Na jego tylnej powierzchni znajduje się kresa chropawa, będąca miejscem przyczepu mięśni. Koniec dalszy kości udowej ma dwa kłykcie służące do połączenia z kością piszczelową. Kłykieć przyśrodkowy jest większy. Kłykcie oddziela głęboki dół międzykłykciowy. Powierzchnie stawowe pokrywające kłykcie łączą się ze sobą od przodu tworząc powierzchnię rzepkową. Na powierzchni zewnętrznej każdego kłykcia znajdują się wyniosłości, zwane odpowiednio nadkłykciami przyśrodkowym i bocznym. Kształt kości udowej zmienia się wraz ze wzrostem dziecka. Podstawowym zmianom ulegają kąt szyjkowo-trzonowy i kąt przodoskręcenia szyjki. Kąt szyjkowo-trzonowy zawarty jest pomiędzy osią szyjki kości udowej a osią trzonu. U niemowlęcia przekracza on zwykle 135°, u osobnika dorosłego w y n o s i 115-120°. Kąt przodoskręcenia szyjki powstaje w w y n i k u skrzyżowania się nałożonych na siebie osi szyjki kości udowej i osi poprzecznej ciała leżącej w płaszczyźnie kłykci. Kąt przodoskręcenia szyjki u niemowląt przekracza zwykle 30°, u osób dorosłych w y n o s i ok. 12°. Prawidłowe wartości kątów mają bardzo ważne znaczenie w budowie i czynności stawu biodrowego. Kość piszczelowa (tibia) jest podstawową „ n o ś n ą " kością goleni i leży po jej stronie przyśrodkowej. Koniec bliższy kości piszczelowej (głowa kości piszcze76
Wyniosłość międzykłykciowa Głowa k. piszczelowejGuzowatość k. piszczelowej
Trzon
Kostka przyśrodkowa Ryc. 48. Kość piszczelowa.
Iowej) ukształtowany jest w postaci kłykci bocznego i przyśrodkowego, których powierzchnie stawowe wchodzą w skład stawu kolanowego. Między kłykciami leży wyniosłość międzykłykciowa złożona z d w ó c h g u z k ó w : bocznego i przyśrodkowego. U podstawy kłykcia bocznego po jego boczno-tylnej stronie leży powierzchnia s t a w o w a strzałkowa, służąca do połączenia z głową strzałki. Trzon kości piszczelowej ma przekrój trójkątny. Z trzech brzegów trzonu najbardziej w y d a t n y jest brzeg przedni. Rozpoczyna się on poniżej kłykci chropawą wyniosłością, zwaną guzowatością piszczeli. Brzeg przedni kości piszczelowej i jej powierzchnia przyśrodkowa przykryte są tylko skórą i dobrze przez nią wyczuwalne. Rozszerzony koniec dalszy piszczeli zakończony jest wyrostkiem kostnym, zwanym kostką przyśrodkową, dobrze wyczuwalną przez skórę. Po stronie bocznej dalszej nasady kości piszczelowej znajduje się wcięcie strzałkowe. Dolna powierzchnia nasady tworzy powierzchnię s t a w o w ą dolną i służy do połączenia z kością skokową wchodzącą w skład kości stępu. Rzepka (patella) jest oddzielną kością wchodzącą w skład stawu kolanowego. Wyróżnia się w niej podstawę oraz skierowany ku d o ł o w i wierzchołek. Tylna stawowa powierzchnia rzepki styka się z powierzchnią rzepkową kości udowej. Powierzchnia przednia jest chropowata i pokrywa ją rozcięgno mięśnia czworogłowego uda. 77
c
\
\f
-Głowa strzałkiPowierzchnia stawowa głowy strzałki
-Trzon-
-Kostka bocznaRyc. 49. Strzałka.
Strzałka (fibula) leży po stronie bocznej i nieco do tyłu w stosunku do piszczeli. Koniec bliższy nosi nazwę g ł o w y strzałki. Ma ona powierzchnię stawową do połączenia z powierzchnią stawową strzałkową, leżącą u podstawy kłykcia bocznego piszczeli. Długi i cienki trzon strzałki ma kształt trójkątny. Zgrubiały, dalszy koniec strzałki, nosi nazwę kostki bocznej. Kostka ta wchodzi w skład stawu skokowo-goleniowego. Jest ona dobrze widoczna i wyczuwalna przez skórę. Nieco powyżej kostki znajduje się powierzchnia stawowa wchodząca we wcięcie strzałkowe kości piszczelowej. Kości stopy (ossa pedis) dzielimy na: kości stępu, kości śródstopia i kości palców. Do kości stępu zalicza się kość skokową, kość piętową, kość łódkowatą, trzy kości klinowate i kość sześcienną. Ich układ przedstawia się następująco: Kość Kość piętowa
Kość łódko wata
Kość sześcienna
Kość klinowata boczna
Kość' klinowata' pośrednia
Kość klinowata przyśrodkowa
Kość śródstopia V
Kość śródstopia IV
Kości śródstopia III i II
Kość śródstopia
78
I
I
I
Kość skokowa (talus) leży pomiędzy kośćmi goleni, kością piętową i kością łódkowatą. Najwyżej położoną częścią kości skokowej jest bloczek pokryty powierzchniami s t a w o w y m i : górną, kostkową - przyśrodkową i boczną. Służą one do połączenia z kośćmi goleni. Ku przodowi i przyśrodkowo skierowana jest głowa kości skokowej, na której znajduje się powierzchnia stawowa, służąca do połączenia z kością łódkowatą. Na dolnej powierzchni kości skokowej znajdują się powierzchnie s t a w o w e piętowe do połączenia z kością piętową. Kość piętowa ( calcaneus) jest największą kością stępu. Leży ona pod kością skokową. Najsilniej rozwinięta, skierowana ku tyłowi, część kości piętowej nosi nazwę guza piętowego. Przednia część kości piętowej jest obniżona. Na jej górnej powierzchni leżą powierzchnie s t a w o w e skokowe. Przedni, płasko ścięty, koniec kości piętowej pokrywa powierzchnia s t a w o w a sześcienna, służąca do połączenia z kością o tej samej nazwię. Kość ł ó d k o w a t a (os naviculare) ma kształt miseczki. Jej wklęsła powierzchnia przylega do g ł o w y kości skokowej. Na krawędzi przyśrodkowej kości łódkowatej znajduje się wyczuwalne przez skórę zgrubienie, zwane guzowatością kości łódkowatej. Na stronie wypukłej „miseczki" znajdują się powierzchnie stawowe, służące do połączenia z kośćmi klinowatymi. Kości klinowate J (ossa cuneiformia) leżą między kością łódkowatą a kośćmi śródstopia. Mają one kształt klinów, ostrzami zwróconymi w stronę podeszw o w ą . Odróżnia się kość klinowatą przyśrodkową, pośrednią i boczną. skokowa
Paliczek K. śródstopia K. klinowata K. łódkowata podstawowy przyśrodkowa
K. piętowa
Ryc. 50. Kości stopy od strony przyśrodkowej. K. skokowa
Ryc. 51. Kości stopy od strony bocznej.
79
Kość sześcienna (os cuboideum), większa od kości klinowatych, leży między kością piętową od tyłu, IV i V kością śródstopia od przodu a kością klinowatą boczną od strony przyśrodkowej. r Kości śródstopia (ossa metatarsalia) w liczbie pięciu należą do kości długich. W każdej z nich wyróżnia się podstawę, trzon i głowę. Kość śródstopia I jest najgrubsza. Na podeszwowej stronie g ł o w y leżą dwie małe oddzielne kostki, zwane trzeszczkami. Podstawa V kości śródstopia jest wydłużona w kierunku bocznym, tworząc guzowatość V kości śródstopia. Kości palców stopy (ossa digitorum pedis) składają się z paliczków. Paluch ma dwa paliczki, pozostałe palce składają się z trzech paliczków.
Kości g ł o w y Dane ogólne Kościec g ł o w y nosi nazwę czaszki (cranium). Część górną i tylną czaszki nazywa się czaszką mózgową (mózgoczaszką), część przednio-dolną - c z a s z ką trzewną (trzewioczaszką). Mózgoczaszka jest mocną puszką kostną osłaniającą mózgowie. Czaszka trzewna składa się z szeregu kości o bardzo złożonej budowie. Mieszczą się w niej narządy zmysłów oraz narząd żucia, służący do rozdrabniania pokarmów. Czaszka składa się z kości połączonych szwami, jedynie żuchwa łączy się z pozostałą częścią czaszki za pomocą s t a w ó w żuchwowych. Kość gnykowa nie ma połączeń z czaszką. W budowie czaszki udział biorą następujące kości (p. tab. 3). Tabela3 Czaszka mózgowa K. potyliczna K. czołowa K. klinowa K. sitowa Kk. ciemieniowe Kk. skroniowe
Czaszka trzewna kk. szczękowe kk. jarzmowe kk. podniebienne kk. nosowe małżowiny nosowe dolne kk. łzowe żuchwa lemiesz k. gnykowa
Z praktycznego punktu widzenia wygodniej jest zapoznać się z czaszką jako całością bez wchodzenia w szczegóły anatomiczne poszczególnych kości składowych. Czaszka mózgowa pokryta sklepieniem (calvaria) jest puszką kostną dla pomieszczenia mózgowia. Przednią część tej puszki tworzy kość czołowa (os 80
Szew wieńcowy " "-V.
K. czołowa
^Kość ciemieniowa
Skrzydło większe k. klinowej Szew węgłowy Kanał nosowo-łzowy
K. skroniowa K. potyliczna Otwór słuchowy zewnętrzny
Wyrostek skroniowy żuchwy
Wyrostek kłykciowy
Szczęka
Wyrohluk sutkowaty
żutllwV
Wyrostek rylcowaty Kęt żuchwy
Ryc. 52. Czaszka od strony lewej.
frontale). Wypukłości kostne nad brzegami oczodołów nazywają się ł u k a m i b r w i o w y m i . Między łukami leży gładkie pole kostne, zwane gładzizną. Granicę oczodołów od góry stanowią brzegi nadoczodołowe. Posuwając się wzdłuż kości czołowej ku górze i ku tyłowi napotykamy na szew w i e ń c o w y , miejsce połączenia kości czołowej z kośćmi ciemieniowymi. Obie kości ciemieniowe łączy szew strzałkowy. Ku tyłowi od kości ciemieniowych znajduje się kość potyliczna, która łączy się z kośćmi ciemieniowymi s z w e m w ę g ł o w y m . Na tylno-dolnej powierzchni kości potylicznej leży guzowatość potyliczna zewnętrzna. Od niej w kierunku podstawy czaszki ciągnie się grzebień potyliczny zewnętrzny. Po obu stronach grzebie, ; a przebiegają poprzecznie kresy karkow e , będące miejscem przyczepu mięśni. Boczne zewnętrzne powierzchnie czaszki mózgowej utworzone są z kości skroniowych i częściowo kości klinowych. Spłaszczona i częściowo zaklęśnięta boczna powierzchnia czaszki nosi nazwę dołu skroniowego. Od przodu ograniczają go połączone szwem w y r o s t k i : j a r z m o w y kości skrbniowej i skron i o w y kości jarzmowej. Od zewnątrz dół skroniowy ogranicza łuk j a r z m o w y , powstały z połączenia się wyrostka skroniowego kości jarzmowej z wyrostkiem j a r z m o w y m kości skroniowej. Nieco poniżej i ku t y ł o w i od łuku jarzmow e g o leży przewód słuchowy zewnętrzny. Poza nim znajduje się duży, chropowaty w y r o s t e k s u t k o w a t y , doskonale wyczuwalny przez skórę. Wyrostek ten zawiera w sobie liczne komory wypełnione powietrzem. 6 - Anatomia i fizjologia
81
Wyrostek zebodołowy Podniebienie twarde —-
K. podniebienna _ Nozdrza tylne
Lemiesz -- - Otwór owalny Dół żuchwowy Wyrostek rylcowatyOtwór szyjny zewnętrzny Otwór ry Icowo - sutkowy Wyrostek sutkowaty
I
Dół skrzydłowy Otwór kolcowy Podstawa k. potylicznej Część skalista k. skroniowej Wyrostek kłykciowy Otwór potyliczny wielki
Ryc. 53. Podstawa czaszki od dołu.
Dolna ściana mózgaczaszki nosi nazwę podstawy czaszki. Jej budowa jest bardzo złożona. Oglądając czaszkę od dołu, spostrzega się przede wszystkim o t w ó r potyliczny wielki. Przez otwór ten rdzeń kręgowy łączy się z mózgowiem. Po obu stronach otworu leżą kłykcie kości potylicznej, służące do połączenia z dołkami stawowymi górnymi kręgu szczytowego. Ku przodowi od otworu potylicznego wielkiego leży podstawa kości potylicznej. Bocznie od podstawy po obu jej stronach znajdują się części skaliste kości skroniowych, zwane również piramidami. W bocznej części piramidy, w sąsiedztwie znanego już nam wyrostka sutkowatego kości skroniowej; leży długi" i cienki wyrostek rylcowaty, a obok niego znajduje się o t w ó r rylcowo-sutkowy dla nerwu twarzowego. W połowie długości części skalistej leży duży o t w ó r szyjny zewnętrzny prowadzący do kanału tętnicy szyjnej. Część skalistą kości skroniowej od sąsiednich kości potylicznej i klinowej oddzielają wąskie szczeliny. Na bocznej części kości skalistej, tuż ku przodowi od otworu słuchowego zewnętrznego, znajduje się dół żuchwowy - miejsce stawowego połączenia z wyrostkiem kłykciowym żuchwy. Ku przodowi od części skalistej kości skroniowej leży kość klinowa. W jej tylnej części znajduje się o t w ó r owalny, przez który opuszcza czaszkę nerw żuchwowy - III gałąź v nerwu trójdzielnego. Nieco ku tyłowi i bocznie leży niewielki o t w ó r kolcowy. Część kości klinowej sąsiadująca z nozdrzami 82
Część oczodołowa k. czołowej
Grzebień koguci Blaszka sitowa /Skrzydło niniejsze k. klinowej
Skrzydło mniejsze, k. klinowej
^Siodło • tureckie
Kanał wzrokowy.
- - Skrzydło większe k. klinowej
Otwór okrągłyOtwór owalny— Otwór kolcowy^ Cz. skalista k. klinowej
/
Otwór potyliczny wielki
w •
--Stok —Kanał kłykciowy
Grzebień potyliczny wewnętrzny
Ryc. 54. Wnętrze podstawy czaszki. Kolorami oznaczono podział na dół czaszkowy przedni, środkowy i tylny.
t y l n y m i nosi nazwę w y r o s t k ó w skrzydtowatych. Każdy z w y r o s t k ó w ma dwie blaszki. Blaszka przyśrodkowa ogranicza od tyłu i boku szeroki o t w ó r prowadzący do jamy nosowej, zwany nozdrzami t y l n y m i . Między blaszką przyśrodkową a blaszką boczną leży dół skrzydłowy, w którym przymocowany jest mięsień o tej samej nazwie. Nozdrza tylne przedzielone są przegrodą, której tylną część stanowi lemiesz (vomer). Kostną przegrodę między jamą nosową a jamą ustną tworzy podniebienie twarde. W części tylnej zbudowane jest ono z kości podniebiennej, pozostała, większa część podniebienia twardego, należy do szczęki. Wałkowate zgrubienie kości szczękowej, w którym tkwią zęby, nazywa się w y r o s t k i e m zębodołowym. Wewnętrzna powierzchnia podstawy czaszki podzielona jest na trzy części, zwane dołami czaszkowymi - przednim, środkowym i t y l n y m . Dół czaszkowy przedni Boczne, symetryczne części dołu czaszkowego przedniego tworzą części oczodołowe kości czołowej. Między nimi leży blaszka s i t o w a , nazwana t a k o d licznych przebijających ją otworków, przez które przechodzą nici nerwu węchowego. W środkowej części blaszki wystaje tzw- grzebień koguci, miejsce przyczepu sierpu mózgu. Tylną część dołu czaszkowego przedniego tworzą skrzydła mniejsze kości klinowej, kończące się wyrostkami pochyłymi 83
przednimi. U ich podstawy leży kanał w z r o k o w y , przez który przechodzi nerw wzrokowy. Pod skrzydłem mniejszym znajduje się szczelina oczodołowa górna, która oddziela je od skrzydła większego kości klinowej, wchodzącego w skład dołu czaszkowego środkowego. Dół czaszkowy ś r o d k o w y Dół czaszkowy środkowy ma kształt podobny do motyla. Część środkową dołu zajmuje trzon kości klinowej. Jest on wewnątrz pusty i ma połączenie z jamą nosową. Górna powierzchnia przypomina kształtem siodło, stąd nazwa siodło tureckie. Zagłębienie na górnej powierzchni trzonu nosi nazwę dołu przysadki. Tylną granicę dołu tworzy grzbiet siodła. Po obu stronach siodła leżą płytkie rynienkowate zagłębienia, zwane bruzdami tętnicy szyjnej. W skład każdej części bocznej dołu czaszkowego środkowego wchodzą: skrzydło większe kości klinowej i część piramidy kości skroniowej. W skrzydle większym kości klinowej widnieją trzy otwory. Najbliżej szczeliny oczodołowej górnej leży o t w ó r okrągły. Ku t y ł o w i i bokowi od niego znajdują się znane już nam o t w o r y : o w a l n y i kolcowy. Granicę dołu czaszkowego przedniego i tylnego stanowią górne brzegi części skalistej kości skroniowej oraz grzbiet siodła. Dół czaszkowy tylny Dół czaszkowy tylny ma postać głębokiej jamy. W części centralnej dołu widnieje o t w ó r potyliczny wielki, obok którego mają ujścia kanały n e r w u podjęzykowego. Bocznie od nich leżą kanały kłykciowe, przez które przechodzą żyły. Ku przodowi od otworu potylicznego wielkiego leży część podstawna kości potylicznej. Jej pochylenie w kierunku o t w o r u potylicznego wielkiego nosi nazwę stoku. Po przeciwległej stronie otworu widnieje grzebień potyliczny wewnętrzny który wraz z przebiegającą poprzecznie bruzdą zatoki poprzecznej tworzy wyniosłość krzyżowatą. W dole czaszkowym t y l n y m leży móżdżek. Wewnętrzna powierzchnia sklepienia czaszki jest stosunkowo gładka. Niewielkie zagłębienia, odpowiadające zakrętom mózgu, nazywają się w y c i s k a m i palczastymi. Poza nimi występują płytkie rozgałęzione bruzdy tętnicze i żylne. W sąsiedztwie szwu strzałkowego znajdują się liczne okrągłe zagłębienia, zwane dołeczkami ziarenkowymi. Czaszka trzewna Czaszka trzewna jest utworzona przez kości przedniej części czaszki. Częściowo była już o nich mowa przy opisywaniu przedniego dołu czaszkowego. Obecnie zajmiemy się przednią ścianą czaszki, czyli tzw. twarzą kostną. Poniżej znanych nam już ł u k ó w b r w i o w y c h kości czołowej znajdują się oczodoły. Oczodół (orbita) ma kształt czworościennego ostrosłupa. W jego wierzchołku znajduje się kanał wzrokowy, przez który przechodzą nerw wzrokowy i tętnica oczna. Podstawę wymienionego ostrosłupa stanowi wejście do 84
Ryc. 55. Czaszka widziana od przodu.
oczodołu. Na jego górnej krawędzi leży wcięcie lub o t w ó r nadoczodołowy, przez który przechodzi n e r w nadoczodołowy. Z czterech ścian oczodołu, górnej, dolnej, bocznej i przyśrodkowej, ta ostatnia ma najbardziej złożoną budowę. Ścianę przyśrodkową tworzą cztery kości, a mianowicie: część oczodołowa kości czołowej, blaszka oczodołowa kości sitowej, kość łzowa i powierzchnia oczodołowa szczęki. W obrębie małej, czworokątnej kości łzowej leży bruzda łzowa, która z podobną bruzdą leżącą na wyrostku czołowym szczęki tworzy początek kanału nosowo-łzowego, łączącego oczodół z jamą nosową. W głębi oczodołu widoczne są dwie szczeliny. Szczelina oczodołowa górna prowadzi do wnętrza czaszki. Przez szczelinę tę przechodzą nerwy unerwiające czuciowo gałkę oczną i okolicę podoczodołową oraz nerwy unerwiające mięśnie poruszające gałkę oczną. Między oczodołami leży jama nosowa ( cavum nasi). Przedziela ją kostna przegroda nosowa utworzona z blaszki pionowej kości sitowej i z lemiesza. Wejście do j a m y nosowej kostnej nazywa się o t w o r e m g r u s z k o w a t y m . Ograniczają go brzegi kości nosowych od góry i kości szczękowe od boku i od dołu. Obie kości szczękowe łączą się ze sobą s z w e m międzyszczękowym, a na dolnej krawędzi o t w o r u gruszkowatego tworzą kolec n o s o w y przedni. 85
Na bocznych ścianach jamy nosowej znajdują się blaszki kostne, zwane małżowinami. Widoczna w jamie nosowej przednia część małżowiny nosowej dolnej ma postać wywiniętej ku dołowi i ustawionej w kierunku strzałkowym blaszki kostnej. Powyżej leży częściowo widoczna małżowina nosowa środkowa. Małżowina nosowa górna położona jest najwyżej i jest od przodu niewidoczna. Małżowina nosowa środkowa i górna są częściami kości sitowej. Jama nosowa ma połączenia z zamkniętymi przestrzeniami kostnymi, leżącymi w sąsiedztwie i zwanymi zatokami obocznymi nosa. Rozróżniamy: 1) zatoki czołowe, leżące między przednią i tylną blaszką kości czołowej, 2) zatokę klinową, leżącą w trzonie kości klinowej, 3) zatoki szczękowe (największe), leżące we wnętrzu trzonów kości szczękowych, 4) k o m ó r k i s i t o w e , mające postać niewielkich jamek leżących w labiryncie kości sitowej. Część „twarzy kostnej" leżącą poniżej j a m y nosowej i oczodołów tworzą kości szczękowe i jarzmowe. Tuż poniżej oczodołu znajduje się o t w ó r podoczodołowy, przez który przechodzi nerw podoczodołowy wraz z towarzyszącymi naczyniami. Poniżej o t w o r u leży dół szczęki. Kość j a r z m o w a (oszygomaticum), tworząca wyniosłość policzkową, bierze udział w budowie oczodołu i, łącząc się z wyrostkiem jarzmowym kości skroniowej, tworzy łuk jarzmowy. Dolny zgrubiały brzeg szczęki tworzy w y r o s tek zębodołowy szczęki, w którym tkwią zęby. Żuchwa ( m a n d i b u l a ) jest jedyną ruchomą kością czaszki. Kość składa się z podkowiasto wygiętego t r z o n u żuchwy i zagiętych ku górze gałęzi żuchwy. Na zewnętrznej powierzchni trzonu leży guzowatość bródkowa. Po obu jej stronach na wysokości dolnego zęba przedtrzonowego znajduje się o t w ó r b r ó d k o w y , który prowadzi do kanału kończącego się o t w o r e m żuchwy, leżącym na przyśrodkowej powierzchni gałęzi żuchwy. Przez kanał żuchwy przechodzi III gałąź nerwu trójdzielnego. Gałąź ta unerwia czuciowo zęby żuchwy i dolną część twarzy. Dolna krawędź trzonu żuchwy jest zaokrąglona. W linii środkowej znajduje się tu guzowatość bródkowa. Brzeg górny trzonu żuchwy zajmuje w y r o s t e k zębodołowy, w którym t k w i w zębodołach 16 zębów. W miejscu przejścia trzonu w gałąź żuchwy leży rozwarty kąt żuchwy, który jest dobrze wyczuwalny przez skórę. Gałąź żuchwy kończy się d w o m a wyrostkami: od przodu znajduje się w y r o s t e k dziobiasty, służący jako przyczep mięśnia skroniowego, z tyłu w y r o s t e k kłykciowy. Wyrostek zakończony jest głową w kształcie bloczka, która wchodzi w skład stawu żuchwowego. Poniżej wyrostka kłykciowego leży szyjka żuchwy. Wyrostek dziobiasty od wyrostka kłykciowego oddziela wcięcie żuchwy. Kość g n y k o w a (os hyoideum) leży poniżej żuchwy i nie łączy się bezpośrednio z kośćmi czaszki. Kość gnykowa ma kształt podkowy. Jej środkowa część nosi nazwę trzonu. Do trzonu przymocowane są dwie pary wyrostków, zwanych rogami. Przedłużenie trzonu ku t y ł o w i tworzy rogi większe. Rogi mniejsze leżą na granicy trzonu i rogów większych i skierowane są ku górze. 86
Czaszka n o w o r o d k a Kości sklepienia czaszki rozwijają się złącznotkankowej pokrywy otaczającej mózgowie. Małe punkty kostnienia zapoczątkowują rozwój kości sklepienia, powiększają się one i stopniowo zbliżają do siebie. Na granicach ich zetknięcia dość długo utrzymuje się tkanka łączna. Błoniaste miejsca połączenia trzech lub czterech kości noszą nazwę ciemiączek ( f o n t i c u l i ) . Ciemiączko przednie (czołowe) ma kształt czworoboku. Znajduje się w miejscu połączenia parzystych u noworodka kości czołowych z kośćmi ciemieniowymi. Ciemiączko tylne (potyliczne) ma kształt trójkątny i leży w miejscu zetknięcia się kości potylicznej z kośćmi ciemieniowymi. Z pozostałych dWóch ciemiączko k l i n o w e leży między kośćmi czołową, ciemieniową, skrzydłem większym kości klinowej i łuską kości skroniowej. Ciemiączko sutkowate znajduje się między kośćmi ciemieniową, skroniową i łuską kości potylicznej. Obecność ciemiączek i nie skostniałych połączeń między kośćmi sklepienia czaszki umożliwia dostosowywanie się kształtu czaszki do kanału miednicy małej podczas porodu. Ciemiączka zarastają wkrótce po urodzeniu, tylko ciemiączko czołowe zamyka się w drugim roku życia. UKŁAD WIĘZADŁOWY - SYNDESMOLOGIA
Połączenia kręgów W kręgosłupie występują różnego rodzaju połączenia kręgów. Między trzonami kręgowymi leżą krążki międzykręgowe (discivertebrales), zbudowane z chrząstki. Obwodowa część krążka zawiera liczne włókna sprężyste i nazywa się pierścieniem w ł ó k n i s t y m . Część centralna, półpłynna, bogata w wodę, nosi nazwę jądra miażdżystego. Jądro miażdżyste amortyzuje wstrząsy i pozwala na ruchy między kręgami. Krążki międzykręgowe dostosowują swój kształt do kształtu trzonów. Są one stosunkowo niskie w odcinku szyjnym i wysokie w części lędźwiowej kręgosłupa. Nie ma krążka międzykręgowego między kręgiem szczytowym a o b r o t o w y m . Ostatni krążek międzykręg o w y leży pomiędzy trzonem V kręgu lędźwiowego a kością krzyżową. Połączenia trzonów kręgów wzmacniają więzadła. Więzadło podłużne przednie wiąże ze sobą przednie powierzchnie trzonów i krążków międzykręgowych. Więzadło podłużne tylne wzmacnia tylną część połączeń trzonów od strony kanału kręgowego. Więzadło to rozpoczyna się przy otworze potyliczn y m wielkim czaszki i biegnie do kości krzyżowej. Łuki kręgowe sąsiednich kręgów łączy tzw. więzadło żółte. Wypełnia ono w o l n e przestrzenie między łukami. Zawierając dużą ilość włókien sprężystych, więzadło żółte jest bardzo elastyczne. Napina się ono podczas zginania kręgosłupa i pomaga w jego prostowaniu. Wyrostki poprzeczne kręgów łączą więzadła międzypoprzeczne, a wyrostki kolczyste - więzadła międzykolczyste. Z ostatnimi współdziała więzadło nadkolcowe, przebiegające między szczytami w y r o s t k ó w kolczystych piersiow e g o i lędźwiowego odcinka kręgosłupa. W odcinku szyjnym więzadło nad87
.Vięzadło j ! podłużne
j;'
'•'
-Jądro '! miażdżyste
Więzadło żółte'
tfft-IA
Więzadło—f nadkolcowe I;/
Pierścień
Więzadłomiędzykoiczyste w / ^ ^ f ^ e " " ^
Więzadła stawumiędzykręgowego
ięzadło Podłużne 4" przednie
-
w łóknisty
'
f
i\
dii •A
Ryc. 56. Połączenie kręgów.
kolcowe przechodzi w więzadło karkowe, rozpięte między wyrostkami kolczystymi kręgów szyjnych a grzebieniem potylicznym zewnętrznym kości potylicznej. Wyrostki stawowe sąsiednich kręgów mają powierzchnie stawowe wchodzące w skład stawów międzykręgowych. Wyrostki zachodzą na siebie dachówkowato, tak że wyrostki kręgu leżącego wyżej pokrywają od góry i od tyłu wyrostki kręgu niższego. Wyrostki kręgów szyjnych ustawione są w płaszczyźnie zbliżonej do poziomej, w odcinku piersiowym ustawienie ich jest skośne, zaś w odcinku lędźwiowym pionowe. Poszczególne pary wyrostków stawowych łączy torebka stawowa i więzadła. Połączenie dwóch górnych kręgów szyjnych i kręgu szczytowego z kością potyliczną wymaga oddzielnego omówienia. Kłykcie kości potylicznej łącząc się z dołkami stawowymi górnymi kręgu szczytowego tworzą staw szczytowo-potyiiczny. Zachodzą w nim ruchy zginania i prostowania głowy. Kręg»szczytowy z kręgiem obrotowym łączy się w trzech miejscach; są to: stawy szczytowo-obrotowe boczne prawy i lewy oraz staw szczytowo-obrotowy pośrodkowy między zębem kręgu obrotowego a dołkiem zęba leżącym na łuku przednim kręgu szczytowego. Od tyłu ząb kręgu obrotowego przytrzymuje więzadło poprzeczne. Uniemożliwia ono przesuwanie się zęba kręgu obrotowego do tyłu w kierunku kanału kręgowego. Wszystkie trzy stawy tworzą czynnościową całość pod nazwą stawu szczytowo-obrotowego. Zachodzą w nim ruchy obrotowe wokół osi pionowej przechodzącej przez ząb kręgu obrotowego. 88
Czynność kręgosłupa Kręgosłup jako całość spełnia trzy podstawowe funkcje: 1) jest osią ciała i jego główną podporą, 2) jest narządem ruchu, 3) osłania rdzeń kręgowy. 1. W płaszczyźnie czołowej kręgosłup jest prosty. Pojawienie się wyraźnego bocznego wygięcia kręgosłupa może świadczyć o chorobie zwanej skoliozą. W płaszczyźnie strzałkowej kręgosłup jest wygięty w kształcie podwójnej litery S. Część szyjna kręgosłupa zwrócona jest wypukłością ku przodowi. Krzywizna piersiowa skierowana jest wypukłością do tyłu. Odcinek lędźwiowy kręgosłupa wygięty jest ku przodowi, a odcinek krzyżowy ku tyłowi. Wygięcie do przodu nazywa się tordozą ( l o r d o s i s ) , a do tyłu - kifozą (kyphosis). U n o w o r o d k ó w cały kręgosłup wygięty jest do tyłu. Pierwsza kształtuje się lordoza szyjna. Ma to miejsce w czasie unoszenia głowy przez dziecko leżące na brzuchu. Lordoza lędźwiowa powstaje w wyniku przybierania przez dziecko postawy stojącej. Charakterystyczne wygięcia kręgosłupa człowieka umożliwiają prawidłowe przenoszenie obciążenia masą ciała oraz amortyzują wstrząsy. 2. Budowa kręgosłupa zezwala na ruchy głowy, szyi i tułowia. Ruchy między poszczególnymi kręgami mają niewielki zakres, jednak sumując się umożliwiają dużą swobodę zmiany położenia poszczególnych jego odcinków. Wyróżniamy ruchy w trzech podstawowych płaszczyznach: zginanie i prostow a n i e w płaszczyźnie strzałkowej, pochylanie boczne w płaszczyźnie czołowej i obracanie w płaszczyźnie poprzecznej. Połączenie opisanych ruchów daje ruch zwany obwodzeniem kręgosłupa. Różnice w budowie kręgów powodują, że pochylenie boczne jest większe w odcinku szyjnym i piersiowym, a zginanie i prostowanie w odcinku szyjnym i lędźwiowym. Żebra utrudniają ruchy odcinka piersiowego. Charakterystyczna budowa w y r o s t k ó w stawowych odcinka lędźwiowego uniemożliwia jego ruchy obrotowe. 3. Otwory kręgowe poszczególnych kręgów tworzą łącznie kanał k r ę g o w y (canalis vertebralis), w którym mieści się rdzeń kręgowy. Po obu stronach kręgosłupa widnieją o t w o r y międzykręgowe, przez które przechodzą nerwy rdzeniowe. Rdzeń kręgowy leżący w kanale kręgowym ochraniają od przodu trzony kręgowe i krążniki międzykręgowe, od boku i tyłu rolę tę spełniają łuki kręgowe wraz z przyczepiającymi się do nich więzadłami. Sprężystość kręgosłupa, uwarunkowana jego rozczłonowaniem na elementy kostne i chrzęstne, przyczynia się również do ochrony rdzenia kręgowego przed wstrząsami. W miarę starzenia się organizmu zmienia się również kręgosłup. Wskutek utraty płynu przez jądro miażdżyste krążki międzykręgowe stają się niższe. T u ł ó w skraca się, nasila się kifoza piersiowa, zmniejsza lordoza szyjna i lędźw i o w a . Postawa staje się nieprawidłowa, t u ł ó w pochyla się ku przodowi, występuje tzw. garb starczy. Zmiany związane ze starzeniem mają charakter nieodwracalny, ale i w c i ą g u 89
doby kręgosłup ulega okresowym zmianom. Pod wpływem obciążenia w ciągu dnia krążki międzykręgowe spłaszczają się i kręgosłup się skraca. Po wypoczynku zmiany te cofają się.
Połączenia żeber Kość żebrowa łączy się z kręgosłupem w dwóch punktach. Głowa żebra tworzy staw głowy żebra z dołkiem żebrowym leżącym na krawędziach sąsiednich trzonów, zaś guzek żebra Obraca się na powierzchni dołka żebrowego'wyrostka poprzecznego (staw żebrowo-poprzeczny). Stawy żebrowo-kręgowe wzmacniają więzadła promieniste głowy leżące od przodu. Więzadła śródstawowe głowy dzielą staw na dwie połowy - górną i dolną. Żebra I, XI i XII nie mają tego więzadła, ponieważ każde z nich styka się tylko z jednym kręgiem. Stawy żebrowo-poprzeczne wzmacniają więzadła o tej samej nazwie. Oba stawy mechanicznie stanowią jedną całość. Jest to staw jednoosiowy, w którym ma miejsce obracanie się zebra wokół osi łączącej dołek żebrowy trzonów kręgowych z dołkiem żebrowym wyrostka poprzecznego. Skośny przebieg osi od góry i przodu ku dołowi i tyłowi powoduje, iż unoszenie żeber łączy się z ich odchylaniem ku bokowi. Ma to ważne znaczenie w mechanice oddychania. Przednie końce żeber łączą się z mostkiem za pośrednictwem chrząstek żebrowych. Chrząstki żebra VIII, IX i X łączą się ze sobą tworząc łuk żebrowy. Elastyczne i sprężyste chrząstki żebrowe odkształcają się podczas unoszenia żeber, a oddają energię opuszczając żebra. Twardnienie i kostnienie chrząstek żebrowych występuje w starszym wieku i zmniejszając elastyczność klatki piersiowej utrudnia oddychanie. Klatka piersiowa jako całość Klatka piersiowa człowieka jest spłaszczona w kierunku przednio-tylnym i poszerzona w wymiarze poprzecznym. Od przodu klatkę piersiową ogranicza mostek, z boku żebra, od tyłu część piersiowa kręgosłupa. Otwór górny klatki piersiowej ogranicza I żebro i górny brzeg rękojeści mostka. Otwór dolny klatki piersiowej ma kształt nieregularny i jest ograniczony XII kręgiem piersiowym, XII żebrem, końcem XI żebra, łukami żebrowymi i wyrostkiem mieczykowatym mostka. Klatka piersiowa kobiet jest szersza w części dolnej w stosunku do górnej. U mężczyzn otwór dolny klatki piersiowej jest stosunkowo mniejszy, a górna część klatki piersiowej szersza. Kształt klatki piersiowej jest również zmienny osobniczo. Podstawowe znaczenie ma tu wielkość kąta podmostkowego leżącego pomiędzy łukami żebrowymi obu stron. U osobników astenicznyćh jego wartość zbliżona jest do 60°, podczas gdy u osobników pyknicznych przekracza 90°. Klatka piersiowa bierze udział w oddychaniu. Na ruchy oddechowe klatki 90
piersiowej składają się ruchy żeber kostnych, chrząstek żebrowych, mostka i kręgosłupa. Ruchy żeber i chrząstek żebrowych łączą się z ruchem mostka, który przy unoszeniu żeber przesuwa się ku przodowi i ku górze. Prostowanie piersiowego odcinka kręgosłupa powoduje unoszenie żeber, a zginanie opuszczanie żeber. W w y n i k u opisanych ruchów zmienia się pojemność klatki piersiowej. Podczas wdechu klatka piersiowa zwiększa swój w y m i a r strzałkowy i czołowy. Jej w y m i a r wysokościowy zwiększa się również, ale wskutek skurczu przepony. Dokładniejsze dane dotyczące mechaniki oddychania przedstawiono w rozdz. Układ oddechowy.
Połączenia kończyny górnej Kończyna górna łączy się z t u ł o w i e m poprzez obręcz kończyny górnej. Jedyną kością łączącą kończynę górną wolną poprzez łopatkę z kośćcem tułowia jest obojczyk. Połączenie to ma miejsce w stawie mostkowo-obojczykowym (articulatio sternoclavicularis). Staw mostkowo-obojczykowy W skład tego stawu wchodzą: powierzchnia mostkowa obojczyka i powierzchnia wcięcia obojczykowego rękojeści mostka. Staw jest podzielony na dwie komory przez krążek śródstawowy. Torebkę stawową wzmacniają więzadła m o s t k o w o - o b o j c z y k o w e , międzyobojczykowe (łączące mostkowe końce obu obojczyków) i więzadło obojczykowo-żebrowe, biegnące od mostkowego końca obojczyka do I żebra. Staw obojczykowo-mostkowy jest stawem wieloos i o w y m i aczkolwiek rozległość ruchów w t y m stawie jest niewielka, to przeciwległy koniec obojczyka przesuwa się dość znacznie we wszystkich kierunkach. S t a w barkowo-obojczykowy Staw barkowo-obojczykowy ( a r t i c u l a t i o acromioclavicularis) łączy wyrostek barkowy z powierzchnią stawową barkowego końca obojczyka. Torebkę stawową wzmacniają więzadła barkowo-obojczykowe oraz więzadła łączące obojczyk z wyrostkiem kruczym łopatki. Więzadło kruczo-obojczykowe składa się z dwóch pasm i ono głównie ustala obojczyk względem łopatki. Szerokie mocne więzadło kruczo-barkowe, rozpięte między wyrostkami kruczym i barkowym, nie w p ł y w a na umocowanie obojczyka, a jedynie chroni od góry staw ramienny przed urazami i zapobiega w y s u w a n i u się g ł o w y kości ramiennej ku górze. Ruchy stawu barkowo-obojczykowego polegają na niewielkich przesunięciach o b u kości względem siebie. Zawartość stawu ma ważne znaczenie w mechanice ruchów stawu ramiennego. 91
S t a w ramienny Staw ramienny ( a r t i c u l a t i o humeri) łączy kończynę górną wolną z jej obręczą. Powierzchnie stawowe tworzą głowa kości ramiennej i wydrążenie stawowe łopatki. Płytkie wydrążenie uzupełnia i pogłębia chrzęstny obrąbek Torebka stawowa Ścięgno m. dwugłowego I
Obrąbek stawowy
Ryc. 57. P r z e k r ó j przez s t a w r a m i e n n y .
H
Główka k. ramiennej
-Bloczek k. ramiennej łokciowa
Dołek głowy k. promieniowej
Ścięgno m. dwugłowego ramienia
Ryc. 58. Staw łokciowy otwarty od przodu.
92
stawowy. Do jego wolnego brzegu przyczepia się torebka stawowa. Przyczep ramienny torebki leży w obrębie szyjki anatomicznej. Torebka stawu ramiennego jest luźna, jej uwypuklony ku dołowi fałd nazywa się zachyłkiem pachowym. Zachyłek umożliwia pełne odwodzenie ramienia, a jego zarośnięcie, np. po długotrwałym unieruchomieniu, znacznie ogranicza ruchy w stawie. Torebkę wzmacnia szereg krótkich więzadeł, najmocniejsze z nich to więzadło kruczo-ramienne. Przez torebkę stawu ramiennego przechodzi ścięgno
Ryc. 59. Zakres ruchów w stawie ramiennym.
93
I
0°
Ryc. 60. Ruchy obrotowe w stawie ramiennym: A - położenie pośrednie, B- obracanie na zewnątrz, C - o b r a c a n i e do wewnątrz, D- największy zakres obracania ramienia do wewnątrz.
g ł o w y długiej mięśnia d w u g ł o w e g o ramienia w drodze do przyczepu znajdującego się na guzku nadpanewkowym, leżącym powyżej wydrążenia panewkowego. Staw ramienny należy do s t a w ó w wieloosiowych. Podstawowe kierunki i zakresy ruchów podano na ryc. 59, 60.
Staw łokciowy S t a w ł o k c i o w y (articulatio cubiti) składa sięztrzech stawów: ramienno-łokciowego, ramienno-promieniowego i promieniowo-łokciowego bliższego. W skład stawu ramienno-łokciowego wchodzi wcięcie bloczkowe kości łokciowej i bloczek kości ramiennej. Staw ramienno-promieniowy łączygłówkę kości ramiennej z dołkiem g ł o w y kości promieniowej. Staw promieniowo-łokciowy bliższy powstaje z połączenia wcięcia promieniowego kości łokciowej z o b w o d e m s t a w o w y m głowy kości promieniowej. Obie kości przytrzymuje więzadło pierścieniowate przyczepiające się do krawędzi wcięcia promieniowego kości łokciowej i obejmujące głowę kości promieniowej. Wszystkie trzy stawy obejmuje wspólna torebka stawowa. Stabilność stawu w płaszczyźnie czołowej zapewniają więzadła poboczne. Więzadło poboczne łokciowe łączy nadkłykiec przyśrodkowy kości ramiennej z kością łokciową. Więzadło poboczne promieniowe wzmacnia torebkę stawową od strony bocznej. Ruchy w stawie łokciowym składają się z ruchów w poszczególnych stawach składowych. Staw ramienno-łokciowy i ramienno-promieniowy łącznie 94
tworzą staw jednoosiowy, w którym zachodzą ruchy zginania i prostowania. Ruch prostowania kończy się oparciem wyrostka łokciowego o dno dołu wyrostka łokciowego kości ramiennej. W stawie p r o m i e n i o w o - ł ó k c i o w y m bliższym głowa kości promieniowej ślizga się ruchem o b r o t o w y m we wcięciu p r o m i e n i o w y m kości łokciowej. Jednocześnie dołek głowy kości promieniowej obraca się na główce kości ramiennej. Obracanie kości promieniowej na zewnątrz nazywa się odwracaniem ( s u p i n a t i o ) przedramienia, ruch przeciwny nosi nazwę nawracania (pronatio). Odwracanie i nawracanie odbywa się w stawie promieniowo-łokc i o w y m bliższym i w sprzężonym z nim czynnościowo stawie promieniowo-łokciowym dalszym. Zakres ruchów w w y m i e n i o n y c h stawach przedstawia ryc. 61. /
Ryc. 61. Zakresy ruchów w stawie promieriiowo-łokciowym i łokciowym.
S t a w p r o m i e n i o w o - ł o k c i o w y dalszy tworzy wcięcie łokciowe kości promieniowej i o b w ó d stawowy głowy kości łokciowej. Jama stawu zamknięta jest torebką wzmocnioną więzadłami. Do ustalenia obu kości przedramienia przyczynia się również błoną międzykostna, rozpięta między zwróconymi do siebie krawędziami obu kości. Staw promieniowo-łokciowy dalszy jest stawem jednoosiowym. Oś obrotu łączy głowę kości łokciowej z głową kości promieniowej. Kość promieniowa s w y m końcem dalszym zatacza łuk dookoła g ł o w y kości łokciowej. W położeniu odwracania obie kości przedramienia są równoległe, a dłoń skierowana jest ku przodowi. W nawróceniu kość promieniowa krzyżuje się od przodu z kością łokciową, a dłoń zwraca się ku tyłowi.
95
ą Staw promieniowo-nadgarstkowy Kość promieniowa jest nieco dłuższa od łokciowej. W przedłużeniu kości łokciowej leży krążek s t a w o w y rozciągając się od przyśrodkowej krawędzi dalszej nasady kości promieniowej do wyrostka rylcowatego kości łokciowej. Dolna powierzchnia krążka wraz z powierzchnią stawową dalszej nasady kości promieniowej tworzą łącznie wklęsłą powierzchnię stawową stawu promieniowo-nadgarstkowego. W skład eliptycznej, wypukłej powierzchni tego stawu wchodzą powierzchnie stawowe bliższego szeregu kości nadgarstka z wyjątkiem kości grochowatej. Torebka stawowa stawu promieniowo-nadgarstkowego wzmocniona jest szeregiem krótkich więzadeł. S t a w promieniowo-nadgarstkowy ( a r t i c u l a t i o radiocarpea) jest eliptyczn y m stawem d w u o s i o w y m . Oś długa tego stawu ustawiona jest w płaszczyźnie ręki. Wokół niej zachodzą ruchy zginania i prostowania. Odwodzenie (w stronę promieniową) i przywodzenie odbywa się dookoła osi prostopadłej do płaszczyzny ręki (ryc. 62).
\
Ryc. 62. Zakres ruchów w stawie promieniowo-nadgarstkowym i międzynadgarstkowym.
S t a w y ręki S t a w y międzynadgarstkowe (articuiationes intercarpeae) leżą między bliższym i dalszym szeregiem kości nadgarstka, jak również między poszczególnymi kośćmi nadgarstka. Przebieg powierzchni stawowych jest dość złożony. Wszystkie te stawy mają wspólną torebkę stawową wzmocnioną krótkimi więzadłami. W stawach międzynadgarstkowych mają miejsce drobne przesunięcia wzajemne kości. Najobszerniejszy jest ruch zginania ręki. Zesztywnienie stawów międzynadgarstkowych znacznie ogranicza ruchomość ręki. 96
1
Stawy nadgarstkowo-śródręczne (articulationes carpometacarpeae) leżą na granicy dalszego szeregu kości nadgarstka i podstaw kości śródręcza od II do V. Są to stawy zwarte o małej ruchomości. Staw nadgarstkowo-śródręczny kciuka (articulatio carpometacarpea pollicis), w przeciwieństwie do pozostałych stawów śródręczno-nadgarstkowych, ma duży zakres ruchomości. Staw ten łączy siodełkowate powierzchnie kości czworobocznej większej i podstawę I kości śródręcza. Jest to staw dwuosiowy. Zachodzą w nim ruchy zginania i prostowania oraz odwodzenia i przywodzenia kciuka. Ruch złożony nosi nazwę przeciwstawiania kciuka pozostałym palcom ręki. S t a w y śródręczno-paliczkowe (articulationes metacarpophalangeae) leżą między g ł o w a m i kości śródręcza i podstawami bliższych członków palców. Ich powierzchnie stawowe zbliżone są kształtem do elipsy. Torebki stawowe wzmacniają więzadła. W stawach możliwe są ruchy zginania, prostowania oraz odwodzenia i przywodzenia palców w stosunku do palca III. Stawy międzypaliczkowe (articulationes interphalangeae) leżą między członkami palców. W kciuku występuje tylko jeden staw, w pozostałych palcach jest ich dwa - bliższy i dalszy. Wypukłą powierzchnię stawu międzypaliczkowego stanowi bloczek, w k l ę s ł ą - j e g o odbicie na podstawie dalszego członka palca. Są to stawy jednoosiowe, w których możliwe są jedynie ruchy zginania i prostowania. Ruchom bocznym zapobiegają więzadła poboczne, wzmacniające torebki stawowe. Ręka jako całość Kości nadgarstka (Dołączone więzadłami tworzą podłużną bruzdę nadgarstka, która jest zwrócona w stronę dłoniową. Bruzdę tę zamyka przebiegające poprzecznie więzadło, zwane troczkiem zginaczy. Ogranicza on od strony dłoniowej kanał nadgarstka, przez który przechodzą ścięgna zginaczy, gałęzie nerwowe i naczynia. Dzięki licznym stawom ręka jest narządem bardzo „elastycznym", mającym zdolność przystosowania się do kształtu chwytanych przedmiotów. Precyzyjny c h w y t zapewnia przeciwstawianie kciuka palcowi II (wskazicielowi) i pozostałym palcom ręki. Większe przedmioty ujmowane są w cęgi, których j e d n y m ramieniem jest kciuk, a d r u g i m pozostałe palce. Ręka umożliwia również chwytanie przedmiotów na dłoni ustawionej ku górze i palcami dostosowanymi do kształtu przedmiotu. Cztery palce ręki mogą ponadto pracować jako haki. Ma to miejsce np. podczas odsuwania szuflady stołu palcami zahaczonymi o jej dolną krawędź.
7 - Anatomia i fizjologia
97
Połączenia kończyny dolnej Połączenia obręczy kończyny dolnej Obręcz kończyny dolnej tworzą obie kości miedniczne, kość krzyżowa i kość guziczna. Kości te, łącząc się za pomocą trzech stawów oraz licznych i mocnych więzadeł, tworzą miednicę (pelviś). Kości miedniczne łączą się z kością krzyżową, stawami krzyżowo-biodrowymi, a między sobą spojeniem ł o n o w y m .
S t a w krzyżowo-biodrowy S t a w krzyżowo-biodrowy (articulatio sacroiliaca) tworzą stykające się powierzchnie uchowate kości krzyżowej i biodrowej. Powierzchnie te są nierówne i połączone śródstawowo więzadłami międzykostnymi. Staw wzmacniają od przodu więzadła krzyżowo-biodrowe brzuszne, a od tyłu - więzadła krzyżowo-biodrowe grzbietowe. Od góry wzmacnia staw więzadło biodrow o - l ę d ź w i o w e . Łączy ono wyrostek poprzeczny V kręgu lędźwiowego z tylną częścią grzebienia kości biodrowej. Niezależnie od wymienionych więzadeł kość krzyżową i biodrową łączą przebiegające w dużej odległości od stawu bardzo mocne więzadła krzyżowo-guzowe i krzyżowo-kolcowe. Więzadło krzyżowo-guzowe (lig. sacrotuberale) biegnie od bocznego brzegu kości krzyżowej i guzicznej do guza kulszowego. Więzadło krzyżowo-kolcowe (lig. sacrospinale) łączy boczny brzeg kości krzyżowej i guzicznej z kolcem kulszowym. Oba więzadła zamykają miednicę od boku i tyłu i przekształcają wcięcia kulszowe w o t w o r y kulszowe. Otwór kulszowy większy leży powyżej więzadła krzyżowo-kolcowego, otwór kulszowy mniejszy leży poniżej tego więzadła. Dolne ograniczenie otworu kulszowego mniejszego stanowi więzadło krzyżowo-guzowe. Otwór zasłoniony kości miednicznej zamyka błona zasłonowa, mająca w górnej części otwór, przez który przechodzą naczynia i nerw zasłonowy.
Spojenie ł o n o w e Obie kości miedniczne łączą się ze sobą spojeniem ł o n o w y m (symphysis pubica). Spojenie zbudowane jest z grubej warstwy chrząstki włóknistej. Połączenia wzmacniają więzadła przebiegające powyżej i poniżej spojenia. Niekiedy w spojeniu występuje jama spojenia - szczelinowata przestrzeń ustawiona strzałkowo. Stawy miednicy należą do połączeń ścisłych i ruchy w nich zachodzące mają charakter niewielkich przesunięć. 98
Miednica jako całość Miednica dzieli się na miednicę większą i miednicę mniejszą. Granicę między nimi stanowią kresa łukowata, grzebienie łonowe i przedni brzeg podstawy kości krzyżowej. Miednicę większą ograniczają od boków talerze biodrowe. Ściany miednicy mniejszej utworzone są z kości krzyżowej i guzicznej od tyłu, kości kulszowej, części kości biodrowych oraz więzadeł krzyżowo-guzowych i krzyżowo-kolcowych od boków. Od przodu miednicę mniejszą ograniczają kości łonowe i spojenia łonowe. Miednica mniejsza jest kanałem kostnym, przez który przesuwa się płód podczas porodu. Kanałten rozpoczyna się w c h o d e m do miednicy, ograniczonym kresami łukowatymi, grzebieniami ł o n o w y m i i krawędzią podstawy kości krzyżowej, kończy zaś w y c h o d e m miednicy ograniczonym kością guziczną, więzadłami krzyżowo-guzowymi oraz gałęziami kości ł o n o w e j i kulszowej. Istnieją różnice kształtu miednicy uwarunkowane płcią. Miednica kobieca jest szersza i krótsza. Talerze biodrowe są bardziej rozchylone. Owalny w c h ó d do miednicy mniejszej kobiet jest większy, jego oś długa ustawiona poprzecznie. Spojenie ł o n o w e jest niższe, kąt ł o n o w y zawarty między gałęziami dolnymi obu kości ł o n o w y c h większy. Kanał miednicy małej kobiet ma kształt cylindryczny, w przeciwieństwie do miednicy męskiej, w której jest on lejkowaty. W pionowej postawie ciała płaszczyzna wchodu tworzy z płaszczyzną poziomą kąt ok. 60°. Nazywamy go kątem pochylenia miednicy. W praktyce położniczej istnieje konieczność oceny b u d o w y miednicy na podstawie p o m i a r ó w wykonywanych na osobniku żywym. Najważniejsze z nich to: 1. Szerokość miednicy międzykolcowa (odległość między kolcami biodrow y m i przednimi g ó r n y m i wynosi przeciętnie 25 do 26 cm). 2. Największa szerokość miednicy, mierzona na najbardziej oddalonych punktach grzebieni biodrowych, wynosi średnio 28-29 cm. 3. Szerokość międzykrętarzowa - 31-32 cm. 4. Sprzężna zewnętrzna (odległość od górnego brzegu spojenia łonowego do wierzchołka wyrostka kolczystego V kręgu lędźwiowego) - średni w y m i a r 18-21 cm. 5. Sprzężna anatomiczna - odległość między przednią krawędzią podstawy kości krzyżowej lub tzw. w z g ó r k i e m (promontorium) a g ó r n y m brzegiem spojenia łonowego - średnio 11 cm. Wymienione powyżej i szereg innych pomiarów pozwalają położnikowi ocenić prawidłowość budowy miednicy, a co za t y m idzie - zapobiec nieprzewidzianym powikłaniom podczas porodu.
Połączenia kończyny dolnej wolnej Staw biodrowy Staw biodrowy ( a r t i c u l a t i o coxae) łączy obręcz kończyny dolnej z kończyną dolną wolną. W skład stawu wchodzi głęboka panewka kości miednicznej i głowa kości udowej. Powierzchnia stawowa panewki ma postać szerokiego, 99
nie zamkniętego od dołu pierścienia obejmującego zagłębienie, zwane dołem panewkowym. Na brzegu panewki przymocowuje się chrzęstny obrąbek panewkowy. Głowa kości udowej objęta jest przez panewkę poza równik. Z tego powodu ruchy w stawie biodrowym mają mniejszy zakres niż w stawie ramiennym.
Ryc. 63. Przekrój przez staw biodrowy.
Torebkę stawu biodrowego wzmacnia szereg więzadeł. Najsilniejsze z nich jest więzadło biodrowo-udowe. Biegnie ono od okolicy kolca biodrowego przedniego dolnego do kresy międzykrętarzowej. Więzadło to hamuje nadmierne prostowanie uda i ma duże znaczenie w utrzymywaniu wyprostnej postawy ciała. Więzadło łonowo-udowe ogranicza odwodzenie uda, a więzadło kulszowo-udowe zapobiega nadmiernemu przywodzeniu. Więzadło głowy kości udowej leży śródstawowo i łączy dolną część dołu panewkowego z dołkiem głowy kości udowej. U dziecka w więzadle tym przebiegają naczynia zaopatrujące w krew część głowy sąsiadującą z przyczepem więzadła. W wieku późniejszym naczynia te zanikają. Więzadło głowy kości udowej nie ma większego znaczenia mechanicznego. Staw biodrowy należy do stawów wieloosiowych. Prawidłowe zakresy ruchów w t y m stawie przedstawiono na ryc. 64. 100
S t a w kolanowy S t a w kolanowy (articulatio genus) ma złożoną budowę. W jego skład wchodzą powierzchnie stawowe trzech kości: udowej, piszczelowej i rzepki. Płaską stawową powierzchnię piszczeli uzupełniają dwie półksiężycowe chrząstki włókniste, zwane łąkotkami s t a w o w y m i (menisci articulares). Łąkotka przyśrodkowa przyczepia się rogiem przednim ku przodowi od wyniosłości kości piszczelowej, rogiem t y l n y m poza tą wyniosłością. Na obwodzie łąkotka zrasta się z torebką stawową i więzadłem pobocznym przyśrodkowym. Łąkotka boczna jest szersza od przyśrodkowej, zbliżona kształtem do nie zamkniętego koła. Łąkotka przymocowuje się przed i poza wyniosłością kości piszczelowej i zrasta się z torebką stawową. Nie łączy się ona z więzadłem pobocznym strzałkowym. Kość udową i piszczelową łączą mocne więzadła poboczne i więzadła krzyżowe. Więzadło poboczne piszczelowe biegnie od nadkłykcia przyśrodkow e g o kości udowej do kości piszczelowej. Zrasta się ono z torebką stawową i łąkotką przyśrodkową. Więzadło poboczne strzałkowe ma postać powrózka i łączy nadkłykiec boczny z głową strzałki. Więzadło nie zrasta się z torebką stawową. Więzadła poboczne są napięte przy w y p r o s t o w a n y m kolanie i w t y m położeniu uniemożliwiają ruchy boczne goleni. Więzadła krzyżowe leżą śródstawowo. Więzadło krzyżowe przednie biegnie od przyśrodkowej powierzchni kłykcia bocznego kości udowej do pola międzykłykciowego przedniego. Więzadło krzyżowe tylne łączy powierzchnię boczną kłykcia przyśrodkowego kości udowej z polem międzykłykciowym t y l n y m piszczeli. Krzyżując się ze sobą, więzadła zapobiegają w y s u w a n i u się kości piszczelowej ze stawu kolanowego do przodu (więzadło krzyżowe przednie) lub do tyłu (więzadło krzyżowe tylne). Więzadła te, owijając się dookoła siebie, hamują ruch obracania goleni do wewnątrz. Z powierzchnią rzepkową kości udowej styka się rzepka. Do rzepki przymocowuje się ścięgno mięśnia czworogłowego uda. Ścięgno to zrasta się z przednią powierzchnią rzepki i przechodzi bezpośrednio w więzadło rzepki, kończące się na guzowatości piszczeli. Torebka stawowa przymocowuje się na krawędziach kłykci piszczeli, bocznych brzegach rzepki i krawędziach kłykci kości udowej. Torebka ma szereg uchyłków, z których największy, zwany kaletką nadrzepkową, ma postać odwróconej do góry kieszonki, leżącej między tylną powierzchnią mięśnia czworogłowego a przednią powierzchnią kości udowej. Staw kolanowyjest najbardziej zwarty w pozycji wyprostowania. Stabilność tę zapewniają napięte więzadła poboczne i tylna część torebki stawowej. Ważną rolę w stabilizacji kolana odgrywa również prawidłowa czynność mięśni, a szczególnie mięśnia czworogłowego uda. Zgięcie kolana zluźnia napięte więzadła i w tej pozycji możliwe są niewielkie ruchy boczne i obrotowe piszczeli. Staw kolanowy należy do stawów jednoosiowych. Teoretycznie oś obrotu przebiega w linii łączącej oba nadkłykcie kości udowej. W rzeczywistości 101
B
Ryc. 64. Zakres ruchów w stawie biodrowym: A - z g i n a n i e , B- prostowanie, C- odwodzenie, Dobracanie na zewnątrz, E- obracanie do wewnątrz.
Więzadło krzyżowe tylne
Więzadło poboczne boczne
Ścięgno m. dwugłowego uda
Więzadła sts.. piszczelowo- strzałkowego bliższej" Błona międzykuoli.a'i
Więzadło krzyżowe przednie Łąkotka boczna Więzadło poboczne przyśrodkowe Więzadło właściwe rzepki Powierzchnia stawowa rzepki
Ryc. 65. Otwarty staw kolanowy.
A
B
Ryc. 66. Zakres ruchów w stawie kolanowym: A- zginanie, B - p r o s t o w a n i e .
położenie osi zmienia się w zależności od ustawienia goleni w stosunku do uda. W początkowej fazie zginania stawu powierzchnie stawowe przetaczają się po sobie, w następnej fazie dochodzi również do ich wzajemnego przesuwania się. Połączenia kości goleni Kość piszczelowa i strzałkowa łączą się stawem piszczelowo-strzałkowym, błoną międzykostną i więzozrostem piszczelowo-strzałkowym. 103
Więzadło przyśrodkowe (trójgraniaste)
Szpara stawu skokowo-goleniowego t Więzadło strzałkowo-skokowe
8 v
K. sześcienna
V-
K. skokowa
-Staw poprzeczny stępu Kości klinowate Staw stępowo-śródstopny
Ryc. 67. Przekrój przez stawy stopy.
S t a w piszczelowo-strzałkowy (articulatio tibiofibułaris) tworzą powierzchnie stawowe głowy strzałki i powierzchnia stawowa strzałkowa kłykcia bocznego kości piszczelowej. Torebkę stawową wzmacniają liczne więzadła. Trzony kości piszczelowej i strzałkowej łączy błona międzykostna goleni. Więzozrost piszczelowo-strzałkowy znajduje się w miejscu połączenia dalszego końca strzałki z wcięciem strzałkowym piszczeli. Więzozrost wzmacniają więzadła piszczelowo-strzałkowe przednie i tylne. Ruchy w stawie i więzozroście piszczeloWo-strzałkowym polegają na niewielkich przesunięciach strzałki w górę i w dół oraz równie niewielkich ruchach obrotowych. Staw skokowo-goieniowy S t a w s k o k o w o - g o i e n i o w y ( a r t i c u l a t i o talocruralis) łączy kości goleni z kością skokową. Połączone więzozrostem dalsze nasady piszczeli i strzałki tworzą wspólną panewkę w kształcie klamry obejmującej bloczek kości skokowej. Kostka przyśrodkowa tej klamry jest dwukrotnie krótsza od kostki bocznej, która ślizga się po powierzchni kostkowej bocznej bloczka kości skokowej. Torebka stawowa przymocowuje się na krawędziach dalszych nasad piszczeli i strzałki i obejmuje bloczek kości skokowej. Torebkę wzmacniają liczne i mocne więzadła. Więzadło przyśrodkowe (trójgraniaste) łączy kostkę przyśrodkową z kośćmi skokową, piętową i łódkowatą. Ma ono przebieg wachla104
rzowaty. Kostka boczna powiązana jest z kośćmi stopy wiązadłami biegnącymi do kości skokowej i piętowej. Staw skokowo-goieniowy należy do stawów jednoosiowych zawiasowych. Oś obrotu łączy tu szczyt kostki przyśrodkowej z połową długości kostki bocznej. Oś ta ustawiona jest skośnie na zewnątrz w stosunku do płaszczyzny czołowej. Staw skokowy jest najbardziej zwarty w pełnym wyprostowaniu stopy. W t y m ustawieniu szerszy z przodu bloczek kości skokowej wklinowuje się między kostki goleni, a silnie napięte więzadła uniemożliwiają ruchy boczne. W zgięciu podeszwowym stopy staw jest rozluźniony i w t y m ustawieniu łatwiej ulega uszkodzeniom urazowym. S t a w y stopy Staw skokowo-piętowo-łódkowy (articulatio talocalcaneonavicularis). Kość skokowa łączy się z kością piętową. Powierzchnie stawowe tylne kości skokowej i piętowej oddzielone są zatoką stępu od mniejszych powierzchni stawowych środkowych i przednich. Torebkę stawową wzmacniają liczne krótkie więzadła. Zatokę stępu wypełniają więzadła i naczynia krwionośne wchodzące do kości skokowej. Głowa kości skokowej przylega do wklęsłej powierzchni kości łódkowatej. Torebka stawowa przyczepia się w sąsiedztwie brzegów powierzchni stawowych. Obie części stawu biorą udział w ograniczonych ruchach odwracania i nawracania stopy. Odwracanie polega na unoszeniu przyśrodkowego brzegu stopy. W czasie nawracania brzeg przyśrodkowy stopy opuszcza się, a unosi brzeg boczny. S t a w poprzeczny stępu (articulatio tarsi transversa), zwany dawniej staw e m Choparta, składa się ze s t a w ó w skokowo-łódkowatego i piętowo-sześciennego. W stawie t y m odbywają się złożone ruchy odwracania i przywodzenia oraz nawracania połączonego z odwodzeniem przedniej części stopy. Kość łódkowata łączy się od przodu z kośćmi klinowatymi, a kość piętowa 105
z kością sześcienną. Ponadto istnieją stawy między kośćmi klinowatymi oj"az staw łączący kość klinowatą boczną z kością sześcienną. Torebki stawowe w y m i e n i o n y c h połączeń wzmacniają liczne więzadła. Niewielkie ruchy w tych stawach polegają na przesunięciach wzajemnych kości, co w p ł y w a na zwiększenie elastyczności stopy. S t a w y stępowo-śródstopne (articulationes tarsometatarseae) znajdują się na granicy dalszych powierzchni stawowych kości klinowatych i kości sześciennej z jednej strony a powierzchniami s t a w o w y m i bliższymi wszystkich kości śródstopia. Kości śródstopia I, II i III łączą się z kośćmi klinowatymi przyśrodkową, pośrednią i boczną. Kości śródstopia IV i V łączą się z kością sześcienną. Wszystkie stawy łącznie tworzą łamaną linię, ponieważ podstawa II kości śródstopia wklinowuje się między kość klinowatą przyśrodkową i boczną. Stawy stępowo-śródstopne określa się w chirurgii wspólną nazwą stawu Lisfranka. Ruchy w stawach stępowo-śródstopnych polegają na niewielkich wzajemnych przesunięciach kości. S t a w y śródstopno-paliczkowe (articulationes metatarsophalangeae) łączą głowy kości śródstopia z wklęsłymi powierzchniami podstaw paliczków. Torebki stawowe wzmacniają więzadła poboczne. W stawach zachodzą ruchy zginania i prostowania palców oraz w mniejszym zakresie przywodzenia i odwodzenia. S t a w y międzypaliczkowe stopy (articulationes interphalangeae pedis) łączą poszczególne członki palców. Wypukłe bloczki znajdujące się na końcach dalszych paliczków łączą się z odpowiednio ukształtowanymi podstawami paliczków. Torebkę stawową każdego stawu wzmacniają więzadła poboczne. Stawy międzypaliczkowe należą do stawów jednoosiowych, w których występują ruchy zginania i prostowania.
Stopa jako całość W rozwoju ontogenetycznym stopa przystosowała się do przenoszenia dużych i zmiennych obciążeń. Kościec stopy tworzy sklepienie podłużne i poprzeczne. Prawidłowo zbudowana stopa dotyka do podłoża guzem kości piętowej oraz g ł o w a m i I i V kości śródstopia. Opiera się ona również częściowo na bocznym brzegu stopy. Brzeg przyśrodkowy stopy jest uniesiony tworząc sklepienie podłużne. Sklepienie poprzeczne tworzy odpowiednie ułożenie kości śródstopia. Taka budowa stopy umożliwia nie tylko przenoszenie dużych obciążeń, ale przyczynia się również do amortyzacji wstrząsów. Stopę jako całość wzmacniają liczne więzadła. Ich osłabienie powoduje powstanie zniekształcenia stopy, do których należy przede wszystkim -stopa płaska. Do najważniejszych więzadeł stopy należą więzadło piętowo-łódkowe i więzadło podeszwowe długie. Więzadło p i ę t o w o - ł ó d k o w e (ligamentum calcaneonaviculare), grubości około 5 m m , łączy obie w y m i e n i o n e kości i podtrzymuje od dołu głowę kości 106
skokowej. Jego osłabienie przyczynia się do obniżenia g ł o w y kości skokowej, a co za t y m idzie - do obniżenia podłużnego sklepienia stopy. Więzadło p o d e s z w o w e długie ( l i g a m e n t u m plantare longum) rozpoczyna się na kości piętowej ku przodowi od guza piętowego i kończy na podstawach II do V kości śródstopia. Rozcięgno p o d e s z w o w e ( a p o n e u r o s i s p l a n t a r i s ) przebiega podskórnie między kością piętową a kośćmi śródstopia. Ma ono charakter cięciwy łuku, która łączy przednią i tylną część stopy. Napięcie rozcięgna jest dobrze wyczuwalne przez skórę na przyśrodkowej stronie podeszwy.
Połączenia kości czaszki Kości czaszki łączą się ze sobą szwami lub za pomocą chrząstkozrostów, a jedynie żuchwa łączy się z czaszką stawem skroniowo-żuchwowym.
Staw skroniowo-żuchwowy S t a w s k r o n i o w o - ż u c h w o w y ( a r t i c u l a t i o temporamandibularis) tworzą w y rostek kłykciowy żuchwy oraz dół żuchwy kości skroniowej. Krążek s t a w o w y dzieli jamę stawu na dwie części - górną i dolną. Staw obejmuje torebka stawowa wzmocniona więzadłem pobocznym. W czasie otwierania ust krążek stawowy przemieszcza się wraz z wyrostkiem kłykciowym ku przodowi. Ruchy boczne występujące podczas żucia polegają na naprzemiennym przesuwaniu się ku przodowi raz jednego, to znów drugiego wyrostka kłykciowego.
UKŁAD MIĘŚNIOWY - MIOLOGIA
Wstęp Mięśnie są biologicznymi silnikami wykonującymi pracę mechaniczną. Przemiany chemiczne zachodzące w mięśniach wyrażają się skurczem czynnym skróceniem długości mięśnia. Siła ta jest t y m większa, im więcej znajduje się w nim czynnych włókienek mięśniowych, innymi słowy - im mięsień ten jest grubszy, im większa jest powierzchnia przekroju prostopadłego do przebiegu włókien, tzw. przekroju fizjologicznego. 1 cm 2 przekroju fizjologicznego zawiera włókna mięśniowe zdolne rozwinąć siłę od 2 do 10 kp w zależności od rodzaju mięśnia. U człowieka występują trzy rodzaje włókien mięśniowych: 1. Mięśnie gładkie wchodzą w skład ścian naczyń i narządów wewnętrznych. Ich skurcz je§t powolny, długotrwały i niezależny od naszej woli. 2. Mięśnie poprzecznie prążkowane są p o d s t a w o w y m i „ s i l n i k a m i " wchodzącymi w skład narządu ruchu. Ich skurcz jest szybki i zależy od w o l i człowieka. 107
3. Mięsień sercowy zbudowany jest z sieci włókien mięśniowych poprzecznie prążkowanych o specyficznej budowie. Jego czynność jest automatyczna i nie podlega naszej woli. Przedmiotem zainteresowania miologii są mięśnie poprzecznie prążkowane. Praca mięśni tej grupy powoduje zmianę położenia części ciała lub całego ustroju. Mięśnie stanowią 2 / 5 ciężaru ciała i występują w licżbie ok. 400 jednostek anatomicznych.
Budowa mięśni Mięsień poprzecznie prążkowany składa się z w ł ó k i e n mięśniowych, swoistych komórek zwanych miocytami. Wewnątrz komórki znajdują się równolegle ułożone włókienka kurczliwe, tzw. miofibryle. Na końcach komórki miofibryle przechodzą w ścięgno. Włókna mięśniowe układają się w pęczki otoczone cienką błoną łącznotkankową, zwaną omięsną wewnętrzną. Grubsza błona otaczająca wiele pęczków nazywa się omięsną zewnętrzną. Cały mięsień otacza łącznotkankowa powięź. Tworzy ona elastyczną powłokę i oddziela mięsień od mięśni sąsiednich. Część czynną mięśnia nazywamy brzuścem ( venter). Brzusiec kończy się ścięgnem (tendo). Ścięgno jest zbudowane z pasm tkanki łącznej włóknistej zbitej. Mięśnie płaskie - szerokie, mają zakończenia w postaci mocnych blaszek, zwanych rozcięgnami. Mięśnie mają różnorodne kształty. Najprostszą postać stanowi mięsień w r z e c i o n o w a t y , mający obły, wydłużony brzusiec, przechodzący na zwężających się końcach w ścięgna. Mięsień może zawierać wiele brzuśców kończących się w s p ó l n y m ścięgnem. Wtakich przypadkach brzuśce otrzymują nazwę głów. Istnieje więc mięsień d w u g ł o w y , t r ó j g ł o w y i czworogłowy. Dwa brzuśce przedzielone ścięgnem pośrednim tworzą mięsień dwubrzuścowy. Mięśnie wrzecionowate zalicza się do mięśni długich. Mięśnie szerokie mają płaski brzusiec zakończony rozcięgnem. Niektóre z mięśni posiadają brzusiec podzielony na odcinki pasemkami tkanki łącznej włóknistej, zwanymi smugami ścięgnistymi. W zależności od układu mięśni i ścięgien odróżnia się rozmaite f o r m y tych połączeń. W mięśniu w r z e c i o n o w a t y m ścięgna leżą w przedłużeniu brzuśca, w innych mięśniach włókna dochodzą do ścięgna pod kątem z obu jego stron (mięsień pierzasty) lub z jednej strony (mięsień półpierzasty). Poza o m ó w i o n y m i rodzajami mięśni w ustroju występują jeszcze mięśnie krótkie, mające małe w y m i a r y i krótkie ścięgna. Mięśnie mieszane nie dają się zaliczyć do żadnej z wyżej w y m i e n i o n y c h grup (np. mięsień okrężny ust). Podczas skurczu mięsień zbliża do siebie oba przyczepy. Najczęściej są to przyczepy na kościach, w innych przypadkach mogą to być przyczepy na powięziach, torebkach stawowych lub wewnętrznej powierzchni skóry (mięśnie mimiczne). 108
Jak już w s p o m n i a n o , mięśnie mają na ogół dwa przyczepy. Odróżnić należy przyczep początkowy ( o r i g o ) i końcowy (insertio). Przyczep początkowy znajduje się bliżej płaszczyzny pośrodkowej tułowia, a na kończynach bliżej tułowia. Przyczep początkowy jest zwykle punktem stałym mięśnia, podczas gdy przyczep końcowy jest jego punktem ruchomym. W niektórych ruchach oba przyczepy mogą być ruchome albo przyczep końcowy może stać się punktem stałym.
Podstawy miomechaniki Rola mięśni sprowadza się do wykonywania pracy poprzez zbliżenie przyczepów. Największą wydajność osiąga mięsień, który od swego pełnego rozluźnienia przechodzi do pełnego skurczu. Mięsień powodujący ruch w jednym stawie nazywamy jednostawowym, mięśnie dłuższe mogą przerzucać się nad wieloma stawami (mięśnie dwu-, trój- i wielostawowe). Położenie mięśnia lub jego ścięgna w stosunku do osi obrotu stawu warunkuje charakter czynności. Mięśnie leżące ku przodowi od tej osi nazywają się zginaczami, przeciwległe - prostownikami. Po stronie przyśrodkowej leżą przywodziciele, a po bocznej odwodziciele. Mięśnie obracające mają przebieg skośny, spiralny lub poprzeczny. Skośny przebieg mięśni powoduje, iż mogą być one jednocześnie np. mięśniami zginającymi i obracającymi. Zwykle nawet prosty ruch (zginanie palca) wykonuje nie jeden, ale kilka mięśni. Noszą one nazwę mięśni współdziałających (synergistycznych). Mięśnie wykonujące ruch przeciwny noszą nazwę antagonistów. W b r e w pozorom obie grupy mięśniowe działają jednocześnie. Na przykład zginanie palca powodują synergistycznie działające zginacze, ale jednocześnie słabiej pobudzone mięśnie ahtagonistyczne działają jak włączany w miarę potrzeby hamulec. Mięśnie antagonistyczne kontrolują czynność synergistów. Przy zrównoważeniu działania synergistów i antagonistów, czyli podczas obustronnego hamowania, ruchu nie ma, a staw zostaje czynnie ustalony. Wzajemną zależność przeciwstawnych grup mięśniowych precyzyjnie kontroluje układ nerwowy. Ruch dokładny, precyzyjny, w y m a g a ustalenia stawów, które aktualnie nie biorą udziału w ruchu. Czynność tę wykonują mięśnie stabilizujące. Tak więc ruch miogenny wyraża się czynnością mięśni synergistycznych pod kontrolą antagonistów i przy współdziałaniu stabilizatorów. Wspomniane grupy mięśniowe są odpowiedzialne za koordynację ruchów, to jest dokładne i celowe ich wykonywanie. Koordynacja dotyczy siły skurczów poszczególnych g r u p mięśniowych, ich rozległości i następstwa w czasie. Koordynację ruchów zapewnia ośrodkowy układ nerwowy. Koordynacja niektórych ruchów jest przekazywana dziedzicznie (ssanie, oddychanie). Mięśnie działają na kości połączone stawami. Z mechanicznego punktu widzenia kości są dla mięśni dźwigniami. W każdej dźwigni istnieją: 1) punkt 109
podparcia - miejsce, w którym obraca się ramię dźwigni, 2) miejsce przyłożenia siły mięśniowej, 3) miejsce przyłożenia oporu, który musi być przezwyciężony siłą mięśni. 1. Punktem podparcia dźwigni kostnej jest staw, a ściślej oś obrotu, dookoła której odbywa się ruch w stawie. 2. Miejscem przyłożenia siły mięśniowej jest końcowy przyczep ścięgna. Przyczepy umiejscowione blisko stawu umożliwiają szybki ruch przy stosunkowo niewielkiej amplitudzie skurczu mięśnia. W tych przypadkach miejsce przyłożenia oporu znajduje się dalej od miejsca przyłożenia siły, a dźwignię tego typu nazywamy szybkościową. W przeciwieństwie do dźwigni szybkościowej dźwignia siłowa ma punkt przyłożenia siły daleko od stawu. W tyfeh przypadkach nawet słaby mięsień, ale działający na długiej dźwigni, potfafi pokonać duży opór. . 3. Opory występujące podczas ruchu to opory wewnętrzne, jak tarcie, i opory zewnętrzne pokonywane przez mięśnie podczas wykonywania pracy. W ustroju istnieją dwa rodzaje dźwigni. W dźwigni jednoramiennej miejsce przyłożenia siły i oporu znajduje się po jednej stronie w stosunku do punktu podparcia. Dźwignia dwuramienna ma punkt podparcia pomiędzy punktami przyłożenia siły i oporu. Nie cała siła mięśnia jest wykorzystywana do pokonania oporów. Siła ta ma postać wektora zgodnego z przebiegiem końcowego odcinka ścięgna. Rozkłada się on na dwie składowe: składową obrotową i składową stawową. Pierwsza z nich jest siłą użyteczną, składowa stawowa powoduje dociskanie powierzchni stawowych. Zauważmy, że stosunek obu składowych zależy od wielkości kąta ścięgnowo-kostnego. Im kąt ten jest większy, tym składowa obrotowa siły mięśnia jest większa. Zwiększenie odległości między ścięgnem a osią obrotu wpływa korzystnie na rozkład sił. Odległość tę zwiększają wyrośla kostne, do których przyczepiają się ścięgna. Wpływ na powiększenie składowej obrotowej ma również prawidłowe wyjściowe ustawienie kończyny przed podjęciem pracy. Na przykład unosząc ciężki przedmiot najpierw zginamy staw łokciowy. Zestawienie mięśni wraz z opisem ich przyczepów i czynności podano wtab. 4 na końcu rozdziału.
Miologia szczegółowa Mięśnie tułowia W skład mięśni tułowia wchodzą: 1) mięśnie klatki piersiowej, 2) mięśnie brzucha i 3) mięśnie grzbietu. Mięśnie klatki piersiowej Mięśnie klatki piersiowej dzielą się na dwie grupy. Grupę powierzchowną tworzą mięśnie, których zakończenia znajdują się na kościach obręczy kończyny górnej i kości ramiennej. Grupę głęboką tworzą właściwe mięśnie klatki 110
Ryc. 69. Mięśnie tułowia od strony bocznej.
piersiowej, które powodują ruchy żeber. Wraz z przeponą grupa ta tworzy zespół mięśni oddechowych. Największym z mięśni klatki piersiowej jest mięsień piersiowy większy. Przyczepia się on do przymostkowej części obojczyka, przedniej powierzchni mostka i chrząstek żebrowych od S do VI żebra. Jego włókna biegną zbieżnie w kierunku bliższego końca kości ramiennej, kończąc się krótkim ścięgnem na grzebieniu guzka większego. Mięsień przywodzi i obraca ramię do wewnątrz. Uniesione ramię silnie obniża, jak przy rąbaniu drzewa. Po unieruchomieniu przyczepu ramiennego działa jako pomocniczy mięsień wdechowy. Mięsień piersiowy mniejszy leży pod mięśniem piersiowym większym, jego włókna przebiegają pionowo. Rozpoczyna się na wyrostku kruczym łopatki, a kończy na przedniej powierzchni żeber od II do V. Mięsień obniża łopatkę, a przy ustalonej łopatce unosi żebra. Mięsień zębaty przedni przykrywa boczną powierzchnię klatki piersiowej. Poszczególne przyczepy mające kształt zębów znajdują się na żebrach. Mię111
Ryc. 70. Przepona widziana od strony klatki piersiowej.
sień kończy się na d o l n y m kącie i przyśrodkowym brzegu łopatki. Mięsień przesuwa łopatkę do przodu i obraca ją skierowując wydrążenie panewkowe ku górze. Przy ustalonej łopatce jest pomocniczym mięśniem w d e c h o w y m . Mięśnie międzyżebrowe wypełniają przestrzenie międzyżebrowe, łącząc sąsiednie żebra. Mięśnie międzyżebrowe zewnętrzne przebiegają skośnie w kierunku dolnozbieżnym i są mięśniami w d e c h o w y m i . Mięśnie międzyżebr o w e w e w n ę t r z n e mają przebieg górozbieżny; ponieważ obniżają żebra należą do mięśni wydechowych.
Przepona Przepona ( d i a p h r a g m a ) jest szerokim, cienkim i wysklepionym kopulasto ku górze mięśniem, który oddziela jamę klatki piersiowej od jamy brzusznej. W przeponie wyróżnia się trzy części, nazwane od miejsca ich przyczepu. Część lędźwiowa przepony przymocowuje się odnogami na kręgach XII piers i o w y m i II lędźwiowym. Część żebrowa, najszersza, odchodzi od wewnętrznych powierzchni VII do XII żebra. Część m o s t k o w a , najmniejsza, przymocow u j e się do wewnętrznej powierzchni mostka. Wszystkie trzy części przepony kończą się w tzw. ośrodku ścięgnistym, który jest błyszczącą błoną w kształcie liścia koniczyny. Przeponę przebija szereg o t w o r ó w , przez które przechodzą główne naczynia krwionośne i przełyk. W części lędźwiowej przepony leży rozwór przełykowy, przez który przechodzą również towarzyszące przełykowi nerwy błędne. Ku t y ł o w i od rozworu przełykowego, tuż przy kręgosłupie, leży rozwór a o r t o w y , przez który przechodzi tętnica główna i przewód piersiowy limfatyczny. Na granicy przedniego i prawego płatka ośrodka ścięgnistego przepony widnieje obszerny o t w ó r żyły g ł ó w n e j dolnej. 112
M. podłopatkowy
- Mm. międzyżebrowe wewnętrzne
M. zębaty przedni
Mm. międzyżebrowe zewnętrzne
M. najszerszy grzbietu
s-f- M. skośny brzucha wewnętrzny
M. pośladkowy średni
Ryc. 71. Mięśnie tułowia - warstwa głębsza.
Przepona jest głównym mięśniem wdechowym. W czasie skurczu przepona się spłaszcza. Powoduje to powiększenie pionowego wymiaru jamy klatki piersiowej. Jednocześnie zmniejsza się wymiar wysokościowy jamy brzusznej. Dzięki temu skurcze przepony powoduję rytmiczne zmiany ciśnienia w klatce piersiowej i jamie brzusznej. W czasie skurczu przepony ciśnienie w jamie klatki piersiowej obniża się, a w jamie brzusznej wzrasta. Różnica ciśnień umożliwia oddychanie i przyczynia się do wypierania krwi żylnej z jamy brzusznej do jamy klatki piersiowej. Z przeponą jako podstawowym mięśniem wdechowym współpracują mięśnie międzyżebrowe zewnętrzne, mięśnie piersiowe większe i mniejsze oraz mięsień zębaty przedni.
8 - Anatomia i fizjologia
113
Mięśnie brzucha Mięśnie brzucha tworzą przednią i boczne ściany j a m y brzusznej. W skład ściany przedniej wchodzą mięśnie proste, ściany boczne zbudowane są z krzyżujących się włókien mięśni skośnych brzucha zewnętrznych, skośnych brzucha wewnętrznych oraz mięśni poprzecznych brzucha i czworobocznych lędźwi. Od wyrostka mieczykowatego do spojenia ł o n o w e g o ciągnie się kresa biała, oddzielająca mięśnie brzucha prawej i lewej strony. Równolegle po obu stronach biegnie prawy i lewy mięsień prosty. Bocznie od niego leżą mięśnie skośne zewnętrzny i wewnętrzny oraz poprzeczny brzucha. Mięśnie czworoboczne lędźwi biegną równolegle do kręgosłupa. Mięsień prosty brzucha biegnie od zewnętrznej powierzchni chrząstek V-VII żpbra do spojenia łonowego. Na przebiegu włókien mięśniowych znajduje się szereg wstawek łącznotkankowych, zwanych s m u g a m i ścięgnistymi. Mięśnie proste są silnymi zginaczami tułowia. Mięsień skośny zewnętrzny brzucha jest płaskim mięśniem biegnącym od zewnętrznych powierzchni V-XII żeber ku dołowi i przyśrodkowo. Przyczep końcowy mięśnia znajduje się na grzebieniu b i o d r o w y m , więzadle pachwinow y m i kresie białej. Działając jednostronnie obraca t u ł ó w w kierunku przeciwnym, zgina i pochyla go ku bokowi. Obustronny skurcz mięśni wzmaga tłocznię brzuszną. Mięsień skośny w e w n ę t r z n y brzucha leży pod mięśniem skośnym zewnętrznym brzucha, ma dolnorozbieżny kierunek włókien. Mięsień rozciąga się od kresy białej do grzebienia biodrowego i powięzi piersiowo-lędźwiowej. Mięsień zgina, pochyla i obraca t u ł ó w ku swojej stronie. Skurcz obustronny wzmaga tłocznię brzuszną. Mięsień poprzeczny brzucha, leżący najgłębiej, przyczepia się do powięzi piersiowo-grzbietowej i biegnie do kresy białej. Poprzeczny przebieg jego włókien powoduje, że jego skurcz zwiększa tłocznię brzuszną. Mięsień czworoboczny lędźwi leży obok kręgosłupa, łącząc XII żebro z w y rostkami poprzecznymi kręgów lędźwiowych i grzebieniem b i o d r o w y m . Mięsień pochyla t u ł ó w w swoją stronę i obniża XII żebro. Rozcięgna szerokich mięśni brzucha wchodzą w skład pochewki mięśnia prostego brzucha, kresy białej i więzadła pachwinowego (p. tab. 4).
Pochewka mięśnia prostego
brzucha
Pochewka mięśnia prostego brzucha składa się z blaszki przedniej, leżącej przed mięśniem prostym, i tylnej, leżącej poza nim. W górnej części pochewki w skład blaszki przedniej wchodzą: połowa grubości rozcięgna mięśnia skośnego brzucha wewnętrznego i rozcięgno mięśnia skośnego zewnętrznego brzucha. Druga połowa rozcięgna mięśnia skośnego wewnętrznego i rozcięgno mięśnia poprzecznego brzucha tworzą tylną blaszkę pochewki. W dolnej części pochewki ścianę przednią tworzą rozcięgna wszystkich mięśni, tj. skośnych i poprzecznego brzucha. Blaszka tylna tej części pochewki jest 114
cienka, zbudowana tylko z powięzi poprzecznej i otrzewnej ściennej. Granica między górną a dolną częścią blaszki tylnej pochewki nosi nazwę kresy łukowatej. Kresa biała, biegnąca między wyrostkiem mieczykowatym mostka a spojeniem ł o n o w y m , zbudowana jest z krzyżujących się włókien rozcięgien mięśni skośnych i poprzecznych.
Więzadło
pachwinowe
Więzadło pachwinowe jest to łącznotkankowy powrózek biegnący od kolca przedniego górnego kości biodrowej do guzka łonowego. Rozcięgna mięśni brzucha wplatają się w to więzadło, tworząc ponad nim szczelinowatą przestrzeń, zwaną kanałem p a c h w i n o w y m (canalisinguinaHs). Kanał pachwinowy biegnie równolegle do więzadła pachwinowego na przestrzeni 4 - 5 cm od góry i boku, ku dołowi i przyśrodkowo, przebijając ścianę jamy brzusznej. Rozpoczyna się on w jamie brzusznej w połowie długości więzadła pachwinowego pierścieniem p a c h w i n o w y m głębokim, a kończy pierścieniem p a c h w i n o w y m powierzchownym. Ściany kanału pachwinowego tworzą: od przodu rozcięgno mięśnia skośnego brzucha zewnętrznego, od góry dolny brzeg mięśnia skośnego wewnętrznego i poprzecznego brzucha, od dołu więzadło pachwinowe, od tyłu powięź poprzeczna brzucha i otrzewna. Przez kanał pachwinowy przechodzi u mężczyzny powrózek nasienny, u kobiet - więzadło obłe macicy. Wzdłuż kanału mogą przedostawać się do moszny i pod skórę trzewa (jelita, sieć wielka), tworząc tzw. przepukliny pachwinowe (herniae inguinales).
Czynność mięśni brzucha Mięśnie brzucha jako całość spełniają trzy podstawowe czynności: 1. Są mięśniami w y d e c h o w y m i dzięki temu, że ich skurcz powoduje obniżenie żeber, a uciskając na trzewa wtłaczają przeponę do jamy klatki piersiowej. 2. Powodują zwiększenie ciśnienia w jamie brzusznej. Łącznie z przeponą i mięśniami miednicy tworzą tzw. tłocznię brzuszną. Tłocznia brzuszna powstaje w przebiegu szeregu funkcji fizjologicznych (poród, defekacja, oddawanie moczu). Prawidłowe napięcie mięśni brzucha przyczynia się do zachowania prawidłowej postawy ciała. 3. Są mięśniami ruchowymi tułowia. Powodują zginanie tułowia, łącznie z mięśniami kręgosłupa obracają górną część tułowia w stosunku do części dolnej oraz pochylają t u ł ó w w stronę boczną. Mięśnie grzbietu Mięśnie grzbietu dzielą się na trzy warstwy: 1) powierzchowną, 2) pośrednią i 3) głęboką. 1. Powierzchowne mięśnie grzbietu mają końcowe przyczepy na kościach obręczy kończyny górnej i kości ramiennej. Przyczepy początkowe znajdują się na wyrostkach kolczystych kręgów. Do tej grupy mięśni należą: 1) mięsień 115
Ryc. 72. Warstwa powierzchniowa mięśni grzbietu.
czworoboczny, 2) mięsień najszerszy grzbietu, 3) mięsień równoległoboczny i 4) mięsień dźwigacz łopatki. Mięsień czworoboczny ma kształt dużego trójkąta, jego długi bok leży w linii wyrostków kolczystych. W tym miejscu mięsień przyczepia się do kości potylicznej, więzadła karkowego oraz wyrostków kolczystych kręgów piersiowych. Włókna mięśniowe zbiegają się w kierunku łopatki i przyczepiają się do bocznej V 3 części obojczyka, wyrostka barkowego i do grzebienia łopatki. Część górna mięśnia unosi łopatkę i obraca ją wydrążeniem stawowym ku górze. Po ustaleniu przyczepu łopatkowego mięsień pochyla w bok głowę i obraca ją w stronę przeciwną. Część środkowa mięśnia zbliża łopatkę do kręgosłupa, a część dolna obniża łopatkę. Mięsień najszerszy grzbietu, największy z powierzchownych mięśni grzbietu, rozpoczyna się na wyrostkach kolczystych sześciu dolnych kręgów piersiowych, wyrostkach kolczystych kręgów lędźwiowych, kości krzyżowej i grzebieniu biodrowym. Włókna mięśnia biegną zbieżnie i przyczepiają się do grzebienia guzka mniejszego kości ramiennej. Mięsień przywodzi do tyłu i obraca ramię do wewnątrz. Po ustaleniu przyczepu ramiennego (np. podczas zwisu na rękach) podciąga tułów ku górze. Mięsień równoległoboczny i mięsień dźwigacz łopatki przebiegają między wyrostkami kolczystymi dwóch ostatnich kręgów szyjnych i czterech pierw116
Ryc. 73. Mięśnie grzbietu po usunięciu m. czworobocźnego i lewego m. najszerszego grzbietu.
szych piersiowych i do brzegu przyśrodkowego łopatki. Mięśnie zbliżają łopatkę do kręgosłupa. Dzięki skośnemu przebiegowi włókien mięsień dźwigacz łopatki unosi łopatkę ku górze. Mięsień równoległoboczny zbliża łopatkę do kręgosłupa (p. tab. 4). 2. Do w a r s t w y pośredniej mięśni grzbietu zalicza się mięśnie zębate: tylny górny i tylny dolny. Ze względu na swoje przyczepy otrzymały one nazwę mięśni kolcowo-żebrowych (p. tab. 4). 3. Do warstwy głębokiej należą mięśnie wypełniające bruzdę między wyrostkami kolczystymi a kątami żeber. Włókna tych mięśni biegną równolegle lub skośnie do kręgosłupa. Główną czynnością tych mięśni jest prostowanie tułowia, stąd nazwa - mięsień prostownik grzbietu. Mięsień jest najgrubszy w odcinku lędźwiowym i uwypukla się w tym miejscu po obu stronach wyrostków kolczystych. Pokryty jest powięzią lędźwiowó-grzbietową. Mięsień prostownik grzbietu dzieli się na leżący bocznie mięsień biodrowo-żebrowy i przyśrodkowo mięsień najdłuższy. Mięsień biodrowo-żebrowy rozpoczyna się na grzebieniu biodrowym, biegnie ku górze i przyczepia się krótkimi ścięgnami do żeber w okolicy ich kątów. Działając obustronnie, prostuje kręgosłup i obniża żebra. Podczas skurczu jednostronnego pochyla kręgosłup ku tyłowi i ku bokowi. Mięsień najdłuższy rozpoczyna się na grzebieniu biodrowym. Wyrostkach kolczystych kręgów lędźwiowych, grzebieniu pośrodkowym, kości krzyżowej 117
i na wyrostkach poprzecznych VII—XII kręgów piersiowych. Przyczepy końcowe mięśnia najdłuższego znajdują się na żebrach oraz na wyrostkach poprzecznych kręgów piersiowych dolnych i lędźwiowych. Mięsień jest bardzo silnym prostownikiem kręgosłupa. W działaniu jednostronnym prostuje i pochyla oraz obraca kręgosłup w swoją stronę.
Mm. krótkie podpotyliczne
M. płatowy szyi
M. najdłuższy^M. biodrowo- żebrowy M. skośny wewnętrzny brzucha
— M. prostownik grzbietu
Ryc. 74. Głęboka warstwa mięśni grzbietu.
W części szyjno-piersiowej kręgosłupa leżą również mięśnie przyczepiające się na czaszce. Ich czynność polega nie tylko na prostowaniu kręgosłupa, ale również w p ł y w a na ruchy g ł o w y w stawie szczytowo-potylicznym i szczytowo- o b r o t o w y m . Należą do nich mięsień płatowaty, mięsień półkolcowy i mięśnie podpotyliczne. Mięsień p ł a t o w a t y ma przyczep początkowy na wyrostkach kolczystych lll-V kręgów piersiowych, kończy się na wyrostkach sutkowatych i bocznych częściach kości potylicznej oraz na wyrostkach poprzecznych dwóch górnych 118
kręgów szyjnych. Działając obustronnie, mięsień prostuje głowę. W skurczu jednostronnym pochyla i obraca głowę w swoją stronę. Mięsień pótkolcowy składa się z licznych pęczków biegnących od wyrostków poprzecznych 5 dolnych kręgów szyjnych i w y r o s t k ó w poprzecznych kręgów piersiowych do wyrostków kolczystych 6 dolnych kręgów szyjnych i 5 górnych piersiowych. Zgina on kręgosłup w swoją stronę i obraca w stronę przeciwną. Działając obustronnie prostuje kręgosłup. Mięśnie podpotyliczne składają się z 6 jednostek mięśniowych łączących dwa górne kręgi szyjne ze sobą i z kością potyliczną. Mięśnie prostują i obracają głową. Poza w y m i e n i o n y m i dużymi mięśniami istnieję krótkie mięśnie kręgosłupa. Mięśnie międzykoIczyste, łączące wyrostki kolczyste, prostują kręgosłup. Mięśnie międzypoprzeczne - pochylają kręgosłup ku bokowi. Mięśnie łączące wyrostki poprzeczne kręgów niższych z wyrostkami kolczystymi kręgów wyższych obracają kręgosłup w stronę przeciwną (p. tab. 4). -
Powięź
piersiowo-iędźwiowa
Powięź piersiowo-iędźwiowa otacza mięsień prostownik grzbietu i wraz z kręgosłupem tworzy rodzaj pochewki powięziowo-kostnej. Blaszka tylna powięzi ciągnie się od dolnego brzegu mięśnia zębatego tylnego górnego do grzebienia kości biodrowej, przyczepiając się do w y r o s t k ó w kolczystych kręgów piersiowych, lędźwiowych i na grzebieniu krzyżowym pośrodkowym. Boczna krawędź tylnej blaszki przymocowuje się do kątów żeber, a w części lędźwiowej łączy się z blaszką przednią. Blaszka przednia powięzi piersiowo-lędźwiowej biegnie między dolną krawędzią XII żebra a grzebieniem kości biodrowej. Przyśrodkowo przyczepia się do wyrostków poprzecznych, a bocznie łączy się z blaszką tylną. Powięź piersiowo-iędźwiowa umożliwia prawidłową czynność mięśnia prostownika grzbietu, zapobiegając przemieszczaniu się jego podczas ruchów bocznych.
Czynność
mięśni grzbietu
Powierzchowna grupa mięśni grzbietu w p ł y w a na ruchy obręczy kończyny górnej i ramienia. Mięśnie te unoszą obręcz (dźwigacz łopatki, górna część mięśnia czworobocznego), obniżają ją (dolna część mięśnia czworobocznego) i współdziałają w obracaniu łopatki, co ma miejsce przy odwodzeniu ramienia powyżej kąta 90°. Mięsień najszerszy grzbietu jestsilnym przywodzicielem ramienia. W z w i s i e mięsień ten podciąga t u ł ó w ku górze. Grupa głębokich mięśni grzbietu pełni ważną rolę mięśni antygrawitacyjnych, gdyż ich napięcie zapobiega zginaniu tułowia. Asymetryczny skurcz mięśni grzbietu powoduje boczne pochylenie tułowia 119
i głowy. Mięśnie mające skośny przebieg obracają kręgosłup wokół bslz pionowej. Niektóre mięśnie grzbietu działają jako pomocnicze mięśnie oddechowe. Mięsień zębaty tylny górny unosi żebra, jest zatem mięśniem wdechowym. Włókna mięśnia prostownika grzbietu, mające przyczep na żebrach, działają jako mięśnie wydechowe. Nie dotyczy to mięśni ustalających dolne żebra. Prawidłowe czynne ich unieruchomienie stwarza dobre warunki dla pracy przepony, która, jak wiemy, jest głównym mięśniem wdechowym. Mięsień prostownik grzbietu należy do najsilniejszych mięśni ustroju. Jego największa siła znacznie przekracza 300 kp.
Mięśnie szyi i głowy Mięśnie tej grupy dzielą się na powierzchowne i głębokie mięśnie szyi, mięśnie żuchwy oraz mięśnie mimiczne. Tuż pod skórą leży symetryczny mięsień szeroki szyi. Mięsień rozpoczyna się w skórze klatki piersiowej, a kończy w okolicy
M. żwacz
M. dwubrzuścowy M. płatowy głowy
M. tarczowo-gnykowy M. gnykowo- mostkowy
M. czworoboczny
M łopatkowognykowy M. mostkowo-obojczykowo- sutkowy
Ryc. 75. Mięśnie szyi. Usunięty m. szeroki szyi.
120
dolnego brzegu żuchwy. Obniża on żuchwę i kąty ust, współdziała przy otwieraniu ust. Najmocniejszym, dobrze widocznym przez skórę, mięśniem szyi jest mięsień mostkowo-obojczykowo-sutkowy. Przyczepia się on w okolicy stawu obojczykowo-mostkowego, biegnie skośnie ku górze i ku tyłowi i kończy się na wyrostku sutkowatym kości skroniowej. Oba mięśnie, działając wspólnie, prostują, odchylają ku tyłowi i zginają głowę nieco zgiętą. Po ustaleniu górnych przyczepów unoszą mostek, są zatem pomocniczymi mięśniami wdechowymi. Skurcz jednego mięśnia powoduje obrót głowy ku stronie przeciwnej i pochylenie ku stronie własnej, tak że twarz zwraca się ku górze i ku stronie przeciwległej. Nieco głębiej leżą mięśnie pod- i nadgnykowe, łączące kość gnykową z mostkiem, łopatką, chrząstką tarczowatą i żuchwą. Mięśnie te powodują ruchy krtani i opuszczają żuchwę. Do głębokich mięśni szyi należą trzy mięśnie pochyłe, biegnące pomiędzy wyrostkami poprzecznymi kręgów szyjnych a górną powierzchnią I i II żebra, oraz mięśnie długie głowy i szyi, leżące na przedniej powierzchni trzonów kręgów szyjnych.
^ j Ę P
— M. żuchwowo-gnykowy
M. poi
t
M. pochyły środkowy Ryc. 76, Mięśnie szyi. Usunięty m. mostkowo-obojczykowo-sutkowy.
121
Poszczególne grupy mięśni szyi oddzielają łącznotkankowe blaszki noszące nazwę powięzi szyi. Rozróżnia się powięź szyi powierzchowną, przedtchawiczą i przedkręgową. Blaszka powierzchowna otacza mięsień mostkowo-sutkowo-obojczykowy, tworząc dla niego pochewkę. Powięź przymocowuje się na brzegu żuchwy, wyrostku sutkowym, wyrostkach poprzecznych kręgów szyjnych, obojczyku i mostku. Blaszka przedtchawicza ma kształt trójkąta rozpiętego między kością gnykową, mięśniami łopatkowo-gnykowymi i górnymi brzegami obojczyka i mostka. Blaszka przedkręgową pokrywa mięśnie głębokie szyi, przyczepiając się po bokach do wyrostków poprzecznych.
Czynność mięśni szyi jest złożona i różnokierunkowa. Wpływają one na ruchy g ł o w y i szyi, uczestniczą w ruchach żuchwy. Przesuwając chrząstkę tarczową, a z nią krtań, współdziałają w jej czynności głosowej. Mięśnie nadgnykowe tworzą elastyczne i ruchome dno j a m y ustnej. Mięśnie przyczepiające się do obojczyka, mostka i żeber, są ponadto pomocniczymi mięśniami wdechowymi. Mięśnie g ł o w y
Mięśnie
żwaczowe
W skład tej grupy wchodzą mięśnie, których praca związana jest z czynnością żucia. Przyczepy początkowe tych mięśni znajdują się na kościach czaszki, przyczepy końcowe leżą na żuchwie. Najobszerniejszy z nich mięsień skroniow y , wypełnia dół skroniowy czaszki, a jego przyczep końcowy znajduje się na wyrostku dziobiastym żuchwy. Mięsień unosi żuchwę i cofa ją do tyłu. Mięsień żwacz, najmocniejszy z mięśni żuchwy, rozpościera się między łukiem j a r z m o w y m a zewnętrzną powierzchnią kąta żuchwy. Unosi on żuchwę, przez co zwiera zęby. Współdziała z mięśniem żwaczem mięsień skrzydłowy przyśrodkowy, który ma przyczep końcowy na wewnętrznej powierzchni kąta żuchwy. Mięsień skrzydłowy boczny łączy okolicę podskroniową czaszki z wyrostkiem kłykciowym żuchwy. Mięsień ten wysuwa żuchwę ku przodowi (p. tab. 4).
Czynność mięśni żwaczowych Czynność mięśni żwaczowych związana jest z czynnością przewodu pokarm o w e g o i polega na rozcinaniu i miażdżeniu pokarmów wprowadzonych do j a m y ustnej. Mięśnie zwierające szczękę i żuchwę są wielokrotnie silniejsze od mięśni opuszczających żuchwę, które należą do mięśni szyi. Wyróżnia się trzy rodzaje ruchów żuchwy. Unoszenie żuchwy i zwieranie zębów wykonują mięsień skroniowy, mięsień żwacz i mięsień skrzydłowy przyśrodkowy. Wysuwanie żuchwy do przodu powodują mięśnie skrzydłowe boczne i częściowo mięśnie żwacze. Cofanie żuchwy związane jest z czynnością tylnej części mięśnia skroniowego. Ruchy obrotowe żuchwy polegają na jednostronnym przesuwaniu się wyrostka kłykciowego żuchwy z dołu żuchwowego ku przodowi. Czynność tę 122
wykonują działając naprzemiennie mięśnie wysuwające żuchwę do przodu. Kurczenie się mięśni raz po jednej, raz po drugiej stronie powoduje charakterystyczne ruchy żucia. Jak już wspomniano, z mięśniami żuchwy współdziałają mięśnie szyi, które opuszczają i cofają żuchwę do tyłu.
Mięśnie
mimiczne
Mięśnie mimiczne przyczepiają się w skórze twarzy, zmieniają jej rysy, przyczyniając się do odzwierciedlenia stanów psychicznych, takich jak: gniew, smutek lub radość. Mięśnie mimiczne powodują również zamykanie i otwieranie szpar powiekowych, szpary ustnej i w p e w n y m stopniu nozdrzy. Mięśnie zamykające mają przebieg okrężny. Należą do nich mięsień okrężny oka i mięsień okrężny ust. Mięśnie otwierające mają przebieg promienisty i dzielą się na szereg jednostek przyjmujących nazwy od czynności, które wykonują, np. mięsień marszczący brwi, mięsień śmiechowy. Jedynym mięśniem mimicznym, mającym również przyczep na kościach, jest mięsień policzkowy. Łączy on wyrostki zębodołowe szczęki i żuchwy, wchodzi w skład jamy ustnej, tworząc jej^ścianę boczną. Jego skurcz powoduje przesuwanie pokarmu
WęSmBSĘĘgMgm-. — M . czołowy
Ryc. 77. Mięśnie mimiczne.
123
między zęby, pociąga ku bokowi kąciki ust i zwiększa ciśnienie w jamie ustnej jak przy dmuchaniu. Drugi „ d u ż y " mięsień potyliczno-czołowy współdziała z czepcem ścięgriistym. Czepiec ścięgnisty ( galea aponeurotica), jest to rozległe rozcięgno pokrywające sklepienie czaszki. Jest on mocno zrośnięty ze skórą, przesuwa się natomiast swobodnie po sklepieniu czaszki. Ruchy te wykonuje w s p o m n i a n y mięsień czołowo-potyliczny. Czynność mięśni mimicznych Jak już wspomniano, mięśnie mimiczne twarzy wykonują trzy podstawowe czynności: 1) są mięśniami w y r a z o w y m i twarzy, 2) współdziałają w zamykaniu i otwieraniu szpar powiekowych i szpary jamy ustnej oraz 3) niektóre z nich (mięsień policzkowy, mięsień okrężny ust) współdziałają w procesie przyjmowania i przesuwania pokarmów między zębami. Mięśnie kończyny górnej Mięśnie kończyny górnej dzielą się na: 1) mięśnie obręczy kończyny górnej, 2) mięśnie ramienia, 3) mięśnie przedramienia i 4) mięśnie ręki. Mięśnie obręczy kończyny górnej Mięśnie obręczy kończyny górnej mają przyczep początkowy na łopatce i obojczyku; przyczep końcowy umiejscowiony jest na kości ramiennej. Największym z nich jest mięsień naramienny. Mięsień ten przerzuca się ponad stawem ramiennym, obejmując go od przodu, boku i tyłu. Przyczep bliższy mięśnia naramiennego znajduje się na bocznej części obojczyka, wyrostku barkowym i grzebieniu łopatki. Włókna mięśniowe zbiegają się stożkowato ku dołowi, kończąc się na guzowatości naramiennej kości ramiennej. Przednia część mięśnia zgina ramię w stawie ramiennym, boczna odwodzi, a tylna cofa ku tyłowi. Pod mięśniem naramiennym leży pięć krótkich mięśni rozpiętych między guzkami kości ramiennej a łopatką. Mięśnie te współuczestniczą w odwodzeniu i ruchach obrotowych w stawie ramiennym. Zwierają one staw dociskając do siebie powierzchnie stawowe i zapoczątkowują odwodzenie ramienia (tab. 4).
Dół
pachowy
Ściany dołu pachowego tworzą mięśnie: piersiowy większy i najszerszy grzbietu. Dół pachowy (fossa axillaris) uwidacznia się w czasie odwodzenia ramienia. Ma on kształt trójściennej piramidy, której wierzchołek jest skierowany ku górze. Podstawę piramidy tworzy powięź pachowa, rozpięta pomiędzy dolnymi krawędziami mięśnia piersiowego większego i najszerszego grzbietu. Powięź ta jest wciągnięta w głąb dołu pachowego przez łącznotkankowe więzadło pachowe podwieszające, przyczepiające się do wyrostka kruczego. 124
M. riaramienny M. piersiowy większy M. ramienny M. trójgłowy ramienia
M. prostownik palców-
M. dwugłowy ramienia
M. odwodziciel długi kciuka M. prostownik krótki kciuka M. prostownik długi kciuka M. przywodziciel kciuka
Ryc. 78. Mięśnie kończyny górnej.
Czynność mięśni obręczy kończyny górnej Czynność mięśni obręczy kończyny górnej pozostaje w ścisłym związku z czynnością mięśnia piersiowego większego, mięśnia najszerszego gYzbietu, jak również z czynnością mięśni ramienia przyczepiających się na kościach obręczy kończyny górnej. Mięśnie obręczy kończyny górnej są przede wszystkim silnymi odwodzicielami ramienia. Należą do nich: boczna część mięśnia naramiennego, mięsień nadgrzebieniowy i częściowo podgrzebieniowy. Dwa ostatnie mięśnie zapoczątkowują ruch i zwierają staw ramienny. Obrócenie zewnętrzne ramienia stanowi pozycję wyjściową dla szeregu czynności. W t y m położeniu łatwiejsze i obszerniejsze jest odwodzenie ramienia. Do mięśni obracających ramię na zewnątrz należą te, które przyczepiają 125
£
się do tylnej części guzka większego (mięsień obły mniejszy, mięsień podgrzebieniowy). Obracanie ramienia do wewnątrz wykonują mięśnie przyczepiające się do guzka mniejszego i jego grzebienia. Są to mięśnie: obły większy, podłopatkowy oraz najszerszy grzbietu i piersiowy większy. Mięśnie te należą również do silnych przywodzicieli ramienia. Zgina ramię część przednia mięśnia naramiennego, prostuje - część tylna. W ruchach tych współdziałają niektóre mięśnie ramienia.
Mięśnie kończyny górnej w o l n e j
Mięśnie
ramienia
Mięśnie ramienia dzielą się na dwie grupy. Grupa przednia zawiera zginacze przedramienia. Należą do nich: leżący powierzchownie mięsień d w u g ł o w y ramienia i znajdujący się pod nim mięsień ramienny. Trzeci należący do tej
Ryc. 79. Mięśnie ramienia i łopatki od przodu.
126
grupy mięsień kruczo-ramienny nie wpływa na ruchy w stawie łokciowym, natomiast przywodzi i zgina ramię w stawie ramiennym. Najmocniejszy z nich mięsień d w u g ł o w y ramienia głową długą rozpoczyna się na guzku nadpanewkowym łopatki, a głową krótką na wyrostku kruczym. Długie ścięgno początkowe głowy długiej przebiega śródstawowo pokryte torebką stawu ramiennego, po czym przechodzi przez bruzdę międzyguzkową, poniżej której obie głowy mięśnia dwugłowego łączą się ze sobą. Wspólny przyczep końcowy mięśnia znajduje się na guzowatości kości promieniowej oraz w powięzi przedramienia. Mięsień zgina i odwraca przedramię, ponadto głowa długa odwodzi ramię, a krótka przywodzi i zgina ramię. Grupę tylną mięśni ramienia tworzą głowy silnie rozwiniętego mięśnia trójgłowego ramienia. Głowa długa tego mięśnia rozpoczyna się na guzku podpanewkowym łopatki, głowa boczna ma przyczep na tylnej powierzchni kości ramiennej w górnej części trzonu, głowa przyśrodkowa przymocowuje się na dolnej części powierzchni tylnej trzonu kości ramiennej. Między obiema
M. naramienny
Głowa długa m. trójgłowego Głowa boczna m. trojgłowego
- M. dwugłowy ramienia
M. ramienny
M ramiennopromieniowy M, prostownik promieniowy długi ladgarstka
Ryc. 80. Mięśnie ramienia od strony bocznej.
127
g ł o w a m i leży nerw promieniowy przebiegający w bruździe nerwu prómieniow e g o kości ramiennej. Wszystkie trzy g ł o w y mięśnia trójgłowego kończą się w s p ó l n y m ścięgnem na wyrostku łokciowym kości łokciowej. Mięsień trójgłowy ramienia jest bardzo silnym prostownikiem stawu łokciowego. Mięśnie ramienia otacza powięź ramienia, od której odchodzą dwie przegrody międzymięśniowe przyczepiające się do kości ramiennej. Przegroda międzymięśniowa przyśrodkowa i boczna oddziela mięśnie zginacze od prostowników (p. tab. 4).
Mięśnie
przedramienia
Mięśnie przedramienia w większości mają kształt wrzecionowaty. Na o b w o dzie przechodzą w długie ścięgna poruszające ręką i palcami. Mięśnie przedramienia wykonują również ruchy odwracania i nawracania przedramienia. Większość z nich to mięśnie wielostawowe, stąd złożoność ich mechaniki. Mięśnie przedramienia dzieli się na trzy grupy: przednią, boczną i tylną. Mięśnie grupy przedniej działają w większości jako zginacze stawów łokciow e g o i promieniowo-nadgarstkowego oraz jako zginacze palców. Biorą one
jfiSl M. ramienno -promieniow
M. nawrotny obły
M. zginacz promieniowy nadgarstka M. zginacz łokciowy nadgarstka M. dłoniowy długi
Ryc. 81. Mięśnie przedramienia.
128
początek od nadkłykcia przyśrodkowego kości ramiennej. Przyczep dalszy mięśnia nawrotnego obłego znajduje się na trzonie kości promieniowej. Zginacz nadgarstka promieniowy i zginacz nadgarstka łokciowy przymocowują się do dłoniowej powierzchni podstaw kości śródręcza i do kości nadgarstka. Największy z tej grupy mięśni - zginacz powierzchowny palców - kończy się na podstawach paliczków środkowych II, III, IV, V palca. W drodze do palców ścięgna przechodzą przez kanał nadgarstka pod łącznotkankowym troczkiem zginaczy. Na wysokości bliższego paliczka każde ze ścięgien rozdwaja się tworząc szczelinę, przez którą przechodzi ścięgno zginacza głębokiego. Zginacz palców powierzchowny zgina stawy śródręczno-palcowe i międzypaliczkowe bliższe. Warstwa głęboka przedniej grupy mięśni przedramienia składa się z mięś-
M. ramienny
\
M. ramienno-promieniowy
M. zginacz łokciowy nadgarstka M. zginacz długi kciuka M. zginacz głęboki palców
J r Ryc. 82. Warstwa głęboka mięśni przedramienia. 9 - Anatomia i fizjologia
129
nia zginacza kciuka długiego, zginacza głębokiego palców i mięśnia nawrotnego czworobocznego. Początkowy przyczep mięśni warstwy głębokiej znajduje się na trzonach kości przedramienia i na łączącej je błonie międzykostnej. Przyczep końcowy zginacza długiego kciuka leży na podstawie dalszego paliczka kciuka. Zginacz palców głęboki ma cztery ścięgna końcowe dla palców II, III, IV i V. Ścięgna przechodzą przez kanał nadgarstka, po czym każde z nich zmierza do podstawy dalszego paliczka odpowiednich palców, przechodząc przez szczelinę w ścięgnie zginacza powierzchownego palców. Mięsień nawrotny czworoboczny przebiega prawie poprzecznie łącząc przednie powierzchnie dalszych odcinków trzonów kości przedramienia. Do grupy bocznej zalicza się mięśnie pokrywające boczny brzeg kości promieniowej i rozpoczynające się na nadkłykciu bocznym i dalszym końcu kości ramiennej. Do grupy tej należą: mięsień ramienno-promieniowy, pros-
M. ramienno-promieniowy M. prostownik promieniowy długi nadgarstka M. prostownik promieniowy krótki nadgarstka M. prostownik palców
M. odwodziciel długi kciuka M. prostownik krótki kciuka M. prostownik długi kciuka
Ryc. 83. Mięśnie przedramienia od strony bocznej.
130
towniki nadgarstka p r o m i e n i o w y długi i krótki oraz mięsień odwracacz przedramienia. Mięsień ramienno-promieniowy kończy się na dalszej części trzonu kości promieniowej. Mięsień zgina łokieć, odwraca przedramię ustawione w pronacji i odwrotnie - nawraca przedramię ustawione w odwróceniu. . Prostowniki nadgarstka przyczepiają się do grzbietowej powierzchni podstaw II i III kości śródręcza. Mięśnie zginają przedramię w stawie łokciowym, prostują i odwodzą rękę w stronę promieniową. Mięsień odwracacz kończy się na bocznej i przedniej powierzchni kości promieniowej, odwraca on przedramię współdziałając z mięśniem d w u g ł o w y m ramienia. Mięśnie grupy tylnej układają się w dwie warstwy - powierzchowną i głęboką. Mięśnie warstwy powierzchownej rozpoczynają się w s p ó l n y m brzuścem na nadkłykciu bocznym kości ramiennej. Najmocniejszym mięśniem tej grupy jest p r o s t o w n i k palców. Brzusiec mięśnia przechodzi w cztery ścięgna biegnące wspólnie przez pochewkę troczka prostowników na grzbiet ręki, gdzie biegną do rozcięgien grzbietowych palców. Ścięgna prostownika III, IV i V palca łączą więzadła poprzeczne, które utrudniają prostowanie poszczególnych palców. Mięsień p r o s t o w n i k nadgarstka łokciowy kończy się na grzbietowej powierzchni podstawy V śródręcza. Mięsień prostuje nadgarstek i przywodzi go w stronę łokciową. Pozostałe dwa mięśnie tej grupy - to krótki mięsień łokciowy, prostujący przedramię i współdziałający ż mięśniem t r ó j g ł o w y m ramienia, oraz prostownik palca małego. Warstwę głęboką tworzą mięśnie wpływające na ruchy kciuka i palca wskazującego. Mięśnie odwodziciel i p r o s t o w n i k kciuka krótki przyczepiają się na kości promieniowej i do błony międzykostnej. Ścięgna obu tych mięśni biegną skośnie ku dołowi, ograniczając od strony dłoniowej zagłębienie, zwane tabakierką anatomiczną. Mięsień odwodziciel kciuka długi kończy się na podstawie I kości śródręcza. Ścięgno prostownika kciuka krótkiego kończy się na grzbietowej powierzchni paliczka bliższego kciuka. Mięsień p r o s t o w n i k kciuka długi przyczepia się na kości łokciowej i na błonie międzykostnej. Jego ścięgno biegnie skośnie ku d o ł o w i i przyczepia się do podstawy dalszego paliczka kciuka. Ścięgno prostownika kciuka długiego ogranicza tabakierkę anatomiczną od strony grzbietowej. Mięsień prostownik wskaziciela towarzyszy odpowiedniemu ścięgnu mięśnia prostownika palców, kończąc się wraz z nim w rozcięgnie grzbietowym wskaziciela.
Mięśnie ręki Mięśnie ręki leżą na jej powierzchni dłoniowej, powierzchnia grzbietowa zawiera tylko ścięgna mięśni przedramienia. Mięśnie ręki dzielą się na trzy grupy: 1) grupa boczna tworzy tzw. kłąb kciuka - wyniosłość zawierającą mięśnie poruszające kciukiem i I kością śródręcza, 2) grupa przyśrodkowa należy do palca małego i tworzy wyniosłość zwaną kłąbkiem, 3) grupa środkowa zawiera mięśnie właściwe dłoni. 131
M. przywodziciel kciuka M. odwodziciel krótki kciuka M. przeciwstawiacz kciuka
Ryc. 84. Mięśnie ręki od strony dłoniowej.
Mięśnie kłębu współdziałają z mięśniami przedramienia w ruchach kciuka. Należą do nich: m ięsień odwodziciel krótki kciuka, przeciwstawiacz kciuka, zginacz krótki i przywodziciel kciuka. Przyczepy początkowe tych mięśni znajdują się na kościach nadgarstka, przyczepy końcowe na I kości śródręcza i paliczku bliższym kciuka. Mięsień odwodziciel krótki kciuka kończy się na bocznej powierzchni paliczka bliższego. Przeciwstawiacz przyczepia się do bocznego brzegu I kości śródręcza. Przebieg włókien tego mięśnia jest skośny w stosunku do kości. Zginacz krótki działa na podstawę bliższego paliczka. Przywodziciel kciuka ma poprzeczny przebieg włókien, przyczepia się po przyśrodkowej stronie podstawy bliższego paliczka kciuka. Kłębik składa się z szeregu drobnych mięśni ułożonych równolegle do V kości śródręcza. Mięśnie rozpoczynają się na kościach nadgarstka. Odwodziciel palca małego i zginacz krótki palca małego kończą się na podstawie bliższego paliczka. Przeciwstawiacz palca małego kończy się na brzegu łokciowym V kości śródręcza. Czynności mięśni są zgodne z ich nazwami (p. tab. 4). Do mięśni środkowych zalicza się mięśnie giistowate o długich i cienkich brzuścach, oraz mięśnie międzykostne, leżące w przestrzeniach między kośćmi śródręcza. Mięśnie giistowate, w liczbie czterech, odchodzą od ścięgien zginacza głębokiego palców
132
w okolicy nadgarstka i biegną do rozcięgien grzbietowych palców II, III i V, po drodze owijając się dookoła promieniowej strony paliczka bliższego. Mięśnie zginają palce w stawach śródręczno-palcowych, prostują w stawach międzypaliczkowych.
Ryc. 85. Mięśnie ręki od strony grzbietowej.
Mięśnie międzykostne dzieli się na dłoniowe i grzbietowe. Przyczepiają się one do kości śródręcza w okolicy ich podstaw, po czym przechodzą na grzbiet palców przyczepiając się na podstawach paliczków bliższych. Cztery mięśnie międzykostne grzbietowe odwodzą palce II i IV od palca III. Trzy mięśnie międzykostne dłoniowe przywodzą palce II, IV i V do palca III. Jednoczesny skurcz wszystkich mięśni powoduje zginanie bliższych paliczków. 133
Pochewki ścięgien
ręki
Długie ścięgna zginaczy palców otoczone są p o c h e w k a m i w ł ó k n i s t y m i w y słanymi od wewnątrz błoną maziową zapewniającą ślizganie się ścięgien. Obszerna pochewka ścięgien zginacza powierzchownego i głębokiego palców obejmuje ścięgna na wysokości kanału nadgarstka. Łączy się ona z pochewką włóknistą V palca. Pochewki włókniste palców II, III i IV nie łączą się bezpośrednio z pochewką ścięgien zginaczy. Pochewka włóknista długiego zginacza kciuka dłuższa od pozostałych, przechodzi przez kanał nadgarstka, nie łączy się jednak z pochewką ścięgien zginaczy powierzchownych i głębokich. Taki układ anatomiczny sprawia, iż zakażenia pochewek ścięgnistych palców I i V łatwiej szerzą się w kierunku nadgarstka i przedramienia. W okolicy nadgarstka ścięgna przytrzymywane są przez przebiegające poprzecznie włókniste pasma tkanki łącznej, zwane troczkami. Troczek prost o w n i k ó w składa się z sześciu przegród, między którymi przechodzi dziesięć ścięgien prostowników. Troczek zginaczy rozpina się między kością łódeczkowatą i czworoboczną z jednej strony, a kością grochowatą i haczykowatą z drugiej. Zamyka on od strony dłoniowej kanał nadgarstka. Po stronie dłoniowej ręki mięśnie własne i ścięgna zginaczy pokrywa rozcięgno dłoniowe.
Czynność
mięśni kończyny górnej wolnej
Mięśnie zginacze spełniają podstawową rolę w czynności kończyny górnej i są mocniejsze od prostowników. Z prostownikami współdziała na ogół siła ciężkości o b w o d o w y c h odcinków kończyny. Zginacze ramienia działają na przedramię jako na dźwignię szybkościową, natomiast zginacze przedramienia, przyczepiające się do nadkłykcia przyśrodkowego kości ramiennej, zginają staw łokciowy działając na przedramię w charakterze mięśni siłowych na długiej dźwigni. Przy ustaleniu o b w o d o w e g o odcinka kończyny charakter dźwigni ulega odwróceniu. Mięśnie przedramienia stają się szybkościowe, a ramienia siłowe. Wstępne ustawienie przedramienia w zgięciu 30° poprawia warunki czynności zarówno mięśni zginających, jak prostowników. Najsilniejszym zginaczem przedramienia jest mięsień d w u g ł o w y ramienia, jest on jednocześnie najsilniejszym mięśniem odwracającym przedramię. W tej czynności współdziałają z nim mięśnie mające skośny przebieg w stosunku do osi przedramienia. W większości czynności przedramię ustawione jest w zgięciu i lekkim nawróceniu, ręka ustawia się w zgięciu grzbietowym 30° i niewielkim odchyleniu łokciowym, kciuk w przeciwstawieniu do lekko zgiętych palców. Jest to tzw. czynnościowe ustawienie kończyny górnej. Podstaw o w e znaczenie w czynności ręki odgrywa przeciwstawianie kciuka pozostał y m palcom. Na ten ruch składają się: 1) odwodzenie I kości śródręcza (wykonuje mięsień odwodziciel kciuka), 2) przeciwstawianie I kości śródręcza połączone z obrotem kciuka w stawie śródręczno-palcowym (mięsień przeciw134
stawiacz), 3) zginanie kciuka w stawie śródręczno-palcowym i międzypaliczkow y m (mięsień zginacz długi i zginacz krótki kciuka). Budowa ręki pozwala na różnego rodzaju chwyty, wyróżniamy: a) precyzyjny chwyt opuszkami kciuka i palca wskazującego, b) chwyt kciuka z boczną powierzchnią wskaziciela, c) chwyt trzema palcami, jak przy pisaniu ołówkiem, d) chwyt całą ręką, jak przy posługiwaniu się młotkiem, e) c h w y t czterema palcami bez udziału kciuka, jak hakiem, i szereg innych. Duże możliwości c h w y t n e w połączeniu z ruchami o b r o t o w y m i przedramienia i dużym zakresem ruchów w stawie ramiennym i łokciowym tworzą z ręki doskonałe i uniwersalne narzędzie pracy, nie mające sobie równego w przyrodzie.
Mięśnie kończyny dolnej Mięśnie obręczy kończyny dolnej Mięśnie obręczy kończyny dolnej łączą kości miednicy z kością udową i dzielą się na trzy grupy: grupę przednią, grupę tylną (mięśni pośladkowych) i grupę mięśni głębokich.
Grupa przednia mięśni obręczy kończyny dolnej Grupa przednia obejmuje mięśnie zginające staw biodrowy. Najmocniejszy z nich jest mięsień b i o d r o w o - l ę d ź w i o w y . Składa się on z dwóch części: mięsień b i o d r o w o - l ę d ź w i o w y większy rozpoczyna się na trzonach kręgów lędźwiowych, a mięsień b i o d r o w y na całej wewnętrznej powierzchni talerza biodrowego. Oba mięśnie łącząc się ze sobą przechodzą pod więzadłem p a c h w i n o w y m i kończą się m o c n y m ścięgnem na krętarzu mniejszym kości udowej. Mięsień zgina udo aż do jego oparcia o przednią powierzchnię brzucha. Po unieruchomieniu przyczepu udowego pochyla miednicę ku przodowi, co ma miejsce podczas siadania z pozycji leżącej.
Grupa tylna mięśni obręczy kończyny dolnej (mięśnie pośladkowe) Mięśnie pośladkowe są silnymi prostownikami i odwodzicielami uda. Ich czynność umożliwia prawidłową postawę i chód. Mięsień pośladkowy w i e l k i rozpoczyna śię na tylnej części zewnętrznej powierzchni talerza biodrowego, na kości krzyżowej i więzadle krzyżowo-guzowym, kończy na guzowatości pośladkowej trzonu kości udowej. Mięsień prostuje udo w stawie biodrowym, obraca je na zewnątrz i odwodzi. Mięśnie pośladkowe średni i mały łączą zewnętrzną powierzchnię talerza biodrowego z krętarzem większym. Mięśnie odwodzą udo w stawie b i o d r o w y m . Podczas stania na jednej nodze zapobiegają opadaniu miednicy po stronie nie obciążonej, przez co współdziałają w utrzymaniu r ó w n o w a g i ciała. 135
M. lędźwiowy większy M. biodrowy M. naprężacz powięzi szerokiej
M. grzebieniowy M. przywodziciel wielki M. krawiecki M. prosty uda M. obszerny przyśrodkowy M. obszerny boczny
Ryc. 86. Mięśnie obręczy kończyny dolnej i uda od przodu.
Z mięśniami pośladkowymi średnim i m a ł y m współdziała naprężacz powięzi szerokiej (p. Mięśnie uda).
Mięśnie
głębokie
miednicy
Mięśnie głębokie miednicy przyczepiają się na przedniej powierzchni kości krzyżowej, błonie zasłonowej i kości kulszowej w okolicy wcięcia kulszowego mniejszego. Przyczepy końcowe znajdują się na przyśrodkowej powierzchni krętarza większego. Dwa inne mięśnie tej grupy rozpoczynają się na zewnętrznej powierzchni błony zasłonowej i na kości kulszowej, a kończą wspólnie z poprzednimi mięśniami na przyśrodkowej powierzchni krętarza większego. Wszystkie mięśnie tej grupy obracają udo na zewnątrz (p. tab. 4). 136
Czynność
mięśni obręczy kończyny dolnej
Mięśnie miedniczno-udowe ustalają czynnie kość udową względem miednicy. Silny prostownik stawu biodrowego, jakim jest mięsień pośladkowy wielki, zaliczamy do mięśni antygrawitacyjnych. Jego napięcie jest niezbędne dla utrzymania pionowej postawy ciała, ponieważ rzyt środka ciężkości ciała leży ku przodowi od osi obrotu stawów biodrowych. Mięśnie odwodzące udo, a przede wszystkim mięsień pośladkowy średni, zapewniają równowagę ciała podczas stania na jednej nodze i naprzemiennie w czasie chodu. W staniu na jednej nodze przyczep krętarzowy mięśnia pośladkowego średniego i małego jest ustalony, a skurcz mięśni pociąga miednicę w swoją stronę, jednocześnie przemieszcza się rzut środka ciężkości w stronę kończyny obciążanej, padając na jej płaszczyznę podparcia. Osłabienie mięśni pośladkowych średniego i małego powoduje opadanie miednicy po stronie nie obciążonej. Takie zjawisko nosi nazwę objawu Trendelenburga. U osób, u których istnieje objaw Trendelenburga, chód jest utykający, kołyszący, tzw. chód kaczkowaty. Większość mięśni miedniczno-udowych, a w szczególności mięśnie głębokie, obracają udo na zewnątrz. Do wewnątrz obraca udo przednia część mięśnia pośladkowego średniego i mięsień pośladkowy mały. Duża przewaga mięśni obracających udo na zewnątrz powoduje, iż u osób długo leżących ustawienie to utrwala się.
Mięśnie kończyny dolnej w o l n e j
Mięśnie uda Mięśnie uda można podzielić na trzy grupy: przednią, tylną i przyśrodkową. Z mechanicznego punktu widzenia są to mięśnie działające na staw biodrowy, staw kolanowy, albo też na oba stawy równocześnie (mięśnie dwustawowe). Grupa przednia mięśni uda Grupa przednia zawiera mięśnie zginające staw biodrowy. Są to mięsień krawiecki i mięsień czworogłowy uda. Mięsień krawiecki leży powierzchownie, biegnie skośnie od kolca biodrow e g o przedniego górnego ku dołowi i przyśrodkowo. Jego ścięgno przechodzi poża nadkłykciem przyśrodkowym kości udowej i przyczepia się nieco przyśrodkowo od guzowatości kości piszczelowej. Mięsień zgina udo, zgina kolano i obraca udo na zewnątrz. Jest to ruch zakładania nogi na nogę z oparciem goleni na udzie strony przeciwnej. Największym mięśniem kończyny dolnej jest mięsień c z w o r o g ł o w y uda. W jego skład wchodzą: mięsień prosty uda, biegnący od kolca biodrowego przedniego dolnego, oraz mięśnie obszerne - boczny, pośredni i przyśrodkowy - przyczepiające się na trzonie kości udowej. Wymienione cztery mięśnie (stąd mięsień czworogłowy) kończą się w s p ó l n y m ścięgnem na rzepce i d a l e j 137
poprzez więzadło właściwe rzepki - na guzowatości kości piszczelonej. Mięsień prosty uda zgina staw biodrowy. Mięsień czworogłowy jako całość prostuje staw kolanowy. Do mięśni grupy przedniej zalicza się również mięsień naprężacz powięzi szerokiej. Zaczyna się on na grzebieniu biodrowym w sąsiedztwie kolca biodrowego przedniego górnego i za pośrednictwem pasma biodrowo-piszczelowego, biegnącego na bocznej powierzchni uda, zmierza do kości piszczelowej. Mięsień zgina, odwodzi i obraca na zewnątrz udo, zgina kolano wyprostowane i prostuje zgięte.
M. dwugłowy uda
Ryc. 87. Mięśnie obręczy kończyny dolnej i mięśni uda od boku.
Grupa tylna mięśni uda Tylna grupa mięśni uda obejmuje mięśnie prostujące staw biodrowy i zginające staw kolanowy. Należą do nich mięśnie: dwugłowy uda, półbłoniasty, półścięgnisty i smukły. 138
„
M. pośladkowy wielki
M. dwugłowy uda
* — M. półbłoniasty
— M . półścięgnisty
M smukły
Ryc. 88. Mięśnie obręczy kończyny dolnej i mięśni uda od tyłu.
Wymienione mięśnie rozpoczynają się na guzie kulszowym, biegną ku dołowi, po czym rozchodzą się, ograniczając bocznie i przyśrodkowo dół podkolanowy. Mięsień d w u g ł o w y uda ogranicza dół podkolanowy od boku i przyczepia się do głowy strzałki. Głowa krótka tego mięśnia rozpoczyna się na trzonie kości udowej, głowa długa, jak już wspomniano, na guzie kulszow y m . Pozostałe mięśnie (półbłoniasty, półścięgnisty, smukły) zmierzają w kierunku przyśrodkowym, okrążają od tyłu nadkłykiec przyśrodkowy kości udowej i przyczepiają się w sąsiedztwie guzowatości kości piszczelowej. Wraz z przyczepami mięśnia krawieckiego i smukłego tworzą układ ścięgien ( zwany „gęsią stopą". Grupa przyśrodkowa mięśni uda Grupa przyśrodkowa zawiera mięśnie przywodzące udo. Rozpoczynają się one na kości łonowej i kulszowej, a kończą na całej długości trzonu kości udowej. Najdłuższy z nich mięsień smukły biegnie od dolnej gałęzi kości 139
M- pośladkowy mały
Mm. miedniczno- udowe krótkie
M. przywodziciel wielki M. przywodziciel długi
M. obszerny boczny M. obszerny przyśrodkowy
Ryc. 89. Głęboka warstwa mięśni obręczy kończyny dolnej i uda od tyłu.
ł o n o w e j przez całą długość przyśrodkowej powierzchni uda, okrąża od tyłu nadkłykiec przyśrodkowy kości udowej i przymocowuje się do guzowatości piszczeli. Pozostałe cztery mięśnie wypełniają trójkątną przestrzeń między kością łonową, kulszową a kością udową. Posuwając się od góry, napotykamy następujące mięśnie: grzebieniowy, przywodziciel krótki, przywodziciel długi i przywodziciel wielki. Przyczepy dalsze tych mięśni znajdują się na kresie chropawej trzonu kości udowej (p. tab. 4).
Powięzie uda Mięśnie uda otacza powięź szeroka. Boczna część tej powięzi wzmocniona jest przebiegającymi podłużnie w ł ó k n a m i pasma biodrowo-piszczelowego. Pasmo to łączy się od góry z grzebieniem b i o d r o w y m , a na obwodzie przymo140
cowuje się do kłykcia bocznego kości piszczelowej. 2 pasmem biodrowo-piszczelowym łączy się mięsień naprężacz powięzi szerokiej. Poszczególne grupy mięśni oddzielają przegrody międzymięśniowe. Przegroda międzymięśniowa boczna oddziela zginacze od prostowników uda i rozpina się między powięzią szeroką a kresą chropawą kości udowej. Przegroda międzymięśniowa przyśrodkowa oddzielą prostowniki od mięśni przywodzących. Między mięśniem obszernym przyśrodkowym a mięśniami przywodzącymi leży kanał przywodzicieli, zamknięty od strony przyśrodkowej przez blaszkę ścięgnistą. Kanał przywodzicieli łączy okolicę pachwinową z dołem podkolanowym. Przez ten kanał przechodzą główne pnie naczyniowe. Wchodzą one do kanału po przejściu pod więzadłem pachwinowym. Więzadło pachwinowe z kością łonową łączy pasmo ścięgniste, zwane łukiem b i o d r o w o - ł o n o w y m . Dzieli ono przestrzeń zawartą między więzadłem p a c h w i n o w y m a kośćmi miednicy na rozstęp naczyniowy, leżący przyśrodkowo i rozstęp m i ę ś n i o w y , położony od boku. Przez rozstęp mięśniowy przechodzi mięsień lędźwiowo-biodrowy i nerw udowy, przez rozstęp naczyniowy tętnica i żyła udowa. Poniżej więzadła pachwinowego leży trójkąt udowy. Jego granice stanowią więzadło pachwinowe, mięsień krawiecki i mięsień przywodziciel wielki.
Czynność mięśni uda Podstawową rolę w postawie i w chodzie odgrywa mięsień czworogłowy uda. Silnie prostując staw kolanowy należy on do mięśni antygrawitacyjnych. Mięśnie zginające .goleń są w większości prostownikami stawu biodrowego. Grupa przyśrodkowa mięśni przywodzi udo. Mięśnie mające przebieg skośny w stosunku do kości udowej (np. mięsień krawiecki) obracają staw biodrowy.
Mięśnie
goleni
Mięśnie goleni dzielą się na trzy grupy: a) grupę przednią - mięśnie prostowniki stopy, b) grupę t y l n ą - z g i n a c z y i c) grupę boczną, w skład której wchodzą mięśnie strzałkowe. Grupa przednia mięśni goleni Grupa p r o s t o w n i k ó w obejmuje mięśnie przymocowujące się do bocznej powierzchni piszczeli, błony międzykostnej i sąsiadującej z nią powierzchni strzałki. Długie ścięgna tych mięśni przechodzą pod troczkami prostowników na grzbiet stopy. Ścięgno mięśnia piszczelowego przedniego kończy się na kości łódkowatej. Ścięgna prostownika długiego palców kończą się w rozcięgnach grzbietowych palców od II do V. Mięsień prostownik długi palucha przyczepia się do grzbietowej powierzchni podstawy paliczka dalszego palucha. 141
Grupa tylna mięśni goleni
O
Grupa zginaczy zawiera najsilniejszy z mięśni goleni - mięsień trójgłowy łydki oraz głębiej położone mięśnie: piszczelowy tylny, mięsień zginacz palców długi i mięsień zginacz palucha długi. sin* tsBaS
W'
- M. strzałkowy długi - M . piszczelowy przedni -M. prostownik długi palców
-M. strzałkowy krótki -M. prostownik palucha
14 Ryc. 90. Mięśnie goleni od przodu.
Mięsień trójgłowy łydki składa się z mięśnia brzuchatego łydki i mięśnia płaszczkowatego. Mięsień brzuchaty łydki rozpoczyna się dwiema głowami boczną i przyśrodkową - powyżej odpowiednich kłykci kości udowej. Mięsień płaszczkowaty przyczepia się do bliższych końców piszczeli i strzałki. Oba mięśnie kończą się wspólnym ścięgnem piętowym (Achillesa), przymocowującym się do guza piętowego. Mięsień trójgłowy łydki jest bardzo silnym zginaczem podeszwowym stopy. Mięsień brzuchaty zgina ponadto kolano. 142
Zginacze głębokie rozpoczynają się na piszczeli, tylnej powierzchni błony międzykostnej i na strzałce. Ścięgno mięśnia piszczelowego tylnego okrąża od dołu kostkę przyśrodkową i przymocowuje się na kości łódkowatej i kościach klinowatych. Mięsień zgina podeszwowo i odwraca (supinuje) stopę.
M. podkolanowy
M. płaszczkowaty
Głowa przyśrodkowa m. brzuchatego łydki
M. strzałkowy długi
Ryc. 91. Mięśnie goleni od tyłu.
Mięsień zginacza palców długi przebiega poza kostką przyśrodkową piszczeli, jego cztery ścięgna kończą się na dalszych paliczkach II - V palców. Ścięgno zginacza długiego palucha okrąża od tyłu i dołu kostkę przyśrodkową i kończy się na dalszym paliczku palucha. Oba ostatnie mięśnie zginają palce oraz zginają podeszwowo i odwracają stopę.
143
M. podkolanowy
M. piszczelowy tylny
M. strzałkowy długi
M. zginacz długi palucha
M. zginacz długi palców
v \Y Ryc. 92. Głęboka warstwa mięśni goleni od tyłu.
Grupa boczna mięśni goleni Grupa boczna zawiera mięśnie nawracające (pronującę) stopę. Należą do nich mięśnie strzałkowe długi i krótki. Mięśnie rozpoczynają się na kości strzałkowej. Ich ścięgna przechodzą poza kostką boczną. Ścięgno mięśnia strzałkowego długiego zagina się przyśrodkowo na podeszwową stronę stopy, krzyżuje sklepienie poprzeczne i przyczepia się na kości klinowatej przyśrodkowej i podstawie I kości śródstopia. Ścięgno mięśnia strzałkęwego krótkiego przyczepia się do guzowatości V kości śródstopia. Oba mięśnie nawracają i zginają podeszwowo stopę (p. tab. 4).
Powięzie
goleni
Powięź goleni otacza mięśnie i łączy się z o d p o w i e d n i m i powięziami uda i stopy. Odchodzące od powięzi przegrody oddzielają mięśnie strzałkowe od prostowników i zginaczy. 144
M. brzuchaty łydki
Ryc. 93. Mięśnie goleni od boku.
Mięśnie
stopy
Na grzbiecie stopy znajdują się jedynie dwa cienkie mięśnie. Prostownik krótki palców i prostownik krótki palucha rozpoczynają się na kościach stępu. Ich ścięgna wplatają swoje włókna w rozcięgna grzbietowe palców i palucha. Mięśnie podeszwowe stopy dzielą się na trzy grupy: przyśrodkowo leżącą grupę mięśni kłębu palucha, boczną mięśni kłębu palca V i grupę środkową. W skład mięśni kłębu palucha wchodzi mięsień odwodziciel palucha. Leży on po stronie przyśrodkowej stopy i odwodzi paiuch od palca II. Mięsień zginacz krótki palucha przymocowuje się do paliczka podstawowego. Zgina paluch w stawie śródstopno-palcowym. Mięsień przywodziciel palucha ma 10 - Anatomia i fizjologia
i
145
głowę poprzeczną i skośną. Jako całość przywodzi paluch do palca II. Głowa poprzeczna współdziała w czynnym kształtowaniu poprzecznego sklepienia stopy Mięśnie kłębu palca V są znacznie słabsze od mięśni kłębu palucha. Należą do nich odwodziciel, zginacz i przeciwstawiacz palca V.
Mm. międzykostne grzbietowe
Ryc. 94. Mięśnie grzbietowe stopy.
Do grupy mięśni środkowych należy zginacz krótki palców. Biegnie on od guza piętowego, a kończy czterema rozczepionymi ścięgnami na podstawach środkowych paliczków II — V palców. Mięsień zgina palce i wzmacnia sklepienie podłużne stopy. Współdziała z nim mięsień czworoboczny podeszwowy, biegnący od kości piętowej do bocznego brzegu ścięgna długiego zginacza palców. Pozostałe mięśnie grupy środkowej - to mięśnie międzykostne i mięśnie giistowate. Są one słabiej rozwinięte niż w kończynie górnej i spełniają podobną do nich czynność (p. tab. 4). 146
M.zginacz krótki palucha M. czworoboczny podeszwy M. zginacz krótki palca małego M. zginacz krótki palqów M. odwodziciel palca małego M. odwodziciel palucha
Ryc. 95. Mięśnie podeszwowe stopy. Ryc. 96. Warstwa głęboka mięśni podeszwowych stopy.
Czynność mięśni goleni i stopy Mięśnie goleni i stopy przystosowały się do pracy statycznej podczas utrzymywania postawy stojącej i do pracy dynamicznej w chodzie. Do najważniejszych mięśni antygrawitacyjnych goleni należy mięsień trójg ł o w y łydki. Zgina on staw kolanowy i jest bardzo silnym zginaczem podeszwowym stopy. Jego czynność w czasie chodu przejawia się w fazie odbicia stopy od podłoża. Mięśnie grupy przedniej goleni oraz mięśnie odwracające i nawracające stopę regulują jej położenie względem podłoża. 147
i
Głównym zadaniem podeszwowych mięśni stopy jako całości jest wzmacnianie podłużnego i poprzecznego sklepienia stopy. Współdziałają z nimi mięśnie goleni, a przede wszystkim mięsień piszczelowy tylny i w pewnej mierze przedni. Bierne wzmocnienie łuku podłużnego stopy tworzy silne rozcięgno podeszwowe, rozpięte między guzem piętowym i podstawami palców. Tabela 4 MIĘŚNIE Mięśnie klatki piersiowej Mięsień
Przyczep początkowy
M. piersiowy większy przymostkowa część (m. pectoralis major) obojczyka, przednia powierzchnia mostka, chrząstki żebrowe żeber I—VI.
Przyczep końcowy
grzebień guzka więk- przywodzi ramię, obszego kości ramiennej raca ramię ku w e w nątrz, opuszcza podniesione ramię
M. piersiowy mniejszy przednia powierzchnia wyrostek (m. pectoralis minor) II do V żebra patki
kruczy ło-
M. zębaty przedni (m. serratus anterior)
boczna powierzchnia klatki piersiowej od I do IX żebra
kąt d o l n y i brzeg przyś r o d k o w y łopatki
M m . międzyżebrowe zewnętrzne
dolny brzeg żebra wyższego
(mm. intercostales externi) Mm. międzyżebrowe wewnętrzne (mm. intercostales interni)
g ó r n y brzeg żebra niższego (kierunek przebiegu dolnozbieżny)
dolny brzeg wyższego
g ó r n y brzeg żebra niższego (kierunek dolnozbieżny)
żebra
Czynność
obniża łopatkę, przy ustalonej łopatce unosi żebra cz. górna mięśnia przesuwa łopatkę do przodu, cz. dolna obraca łopatkę pociągając do przodu jej kąt dolny unoszą żebra - mięśnie wdechowe
opuszczają żebra mięśnie wydechowe
Mięśnie brzucha M.
prosty brzucha (m. rectus abdominis) M. skośny zewnętrzny brzucha (m. obliquus abdominis externus) M. skośny w e w n ę t r z n y brzucha (m. obliquus abdominis intern us) M. poprzeczny brzucha (m. transversus abdominis)
148
chrząstki V—VII żebra
kość ł o n o w a i spojenie zgina t u ł ó w , obniża łonowe żebra, unosi miednicę
powierzchnia zewnętrzna V—XII żebra
grzebień więzadło
biodrowy, pachwino-
obraca t u ł ó w w stronę przeciwną, zgina i po-
w e , kresa biała
chyla ku bokowi
X - X I I żebro pochewka m. prostego brzucha, kresa biała
powięź l ę d ź w i o w o -biodrowa, grzebień k. b i o d r o w e j
obraca, zgina i pochylą t u ł ó w w swoją stronę, obniża żebra
powięź wewnętrzna VII—XII żebra, powięź piersiowo-iędźwiowa
więzadło pachwinow e , pochewka m. prostego brzucha kresa biała
zwiększa ciśnienie jamie brzusznej
Mięsień
Przyczep początkowy
M. czworoboczny lędźwi (m. quadratus lumborum)
XII żebro, wyrostki po- grzebień b i o d r o w y przeczne kręgów lędźwiowych
Przyczep k o ń c o w y
Czynność obniża XII żebro, zgina bocznie t u ł ó w
Mięśnie grzbietu M. Czworoboczny (m. trapezius)
kość potyliczna, wię- boczna '/ 3 obojczyka, zadło karkowe, w y r o - wyrostek barkowy i stki kolczyste kręgów grzebień łopatki szyjnych i piersiowych
unosi łopatkę, zbliża łopatkę do kręgosłupa, obraca łopatkę przy unoszeniu ramienia powyżej poziomu, pochyla g ł o w ę w bok i skręca w stronę przeciwną, do dołu grzebień guzka mniej- przywodzi szego kości ramien- i obraca ramię do wewnątrz nej
M. najszerszy grzbietu (m. latissimus dorsi)
wyrostki kolczyste 6 dolnych kręgów piersiowych, kręgów lędźwiowych, kość krzyżowa, grzebień biodrowy
wyrostki kolczyste kręgów szyjnych VI i VII oraz piersiowych I i II wyrostki poprzeczne M. dźwigacz łopatki (m. levator scapulae) kręgów szyjnych od I do IV
brzeg przyśrodkowy łopatki
zbliża łopatkę do kręgosłupa
górna część brzegu przyśrodkowego łopatki
M. zębaty tylny g ó r n y (m. serratus posterior superior)
w y r o s t k i kolczyste VI i VII kręgów szyjnych i I-II kręgów piersiowych
powierzchnia zewnętrzna żeber od II do V
unosi i zbliża łopatkę do kręgosłupa, przy ustalonej łopatce pochyla g ł o w ę ku tyłowi i b o k o w i unosi żebra, jest mięśniem w d e c h o w y m
M. zębaty tylny dolny (m. serratus posterior inferior)
powięź lędźwiowo-piersiowa na wysokości w y r o s t k ó w kolczystych kręgów piersiowych XI i XII
żebra od IX do XII
M.
równoległoboczny (m. rhomboideus)
prostuje kręgosłup przy skurczu obustronn y m , zgina bocznie w czasie skurczu jednostronnego
M. prostownik grzbietu (m. errector spinae)
a) M. biodrowo-żebrowy grzebień kości biodro-
(m. iliocostaiis) M. najdłuższy (m. lortgissimus)
ustala żebra dolne pomocniczy mięsień wydechowy
kąty żeber
wej grzebień kości biodro- żebra i wyrostki popw e j , wyrostki kolczys- rzeczne kręgów pierte od I lędźwiowego siowych i szyjnych do IV krzyżowego, w y rostki poprzeczne kręg ó w piersiowych o d VII do XII,
prostuje kręgosłup w działaniu obustronnym, jednostronnie zgina bocznie kręgosłup
149
Mięsień
Przyczep początkowy
Przyczep k o ń c o w y
Czynność
b) M. płatowaty . (m. splenius)
wyrostki kolczyste 5 górnych kręgów piersiowych i 5 dolnych szyjnych
wyrostki sutkowate i boczne części kości potylicznej, wyrostki poprzeczne 2 górnych kręgów szyjnych
działając obustronnie prostuje g ł o w ę , jednostronnie obraca i pochyla g ł o w ę w swoją stronę •
c) M. półkolcowy (m. semispinalis)
wyrostki poprzeczne kręgów piersiowych i 5 dolnych szyjnych
wyrostki kolczyste 5 górnych kręgów piersiowych, 6 dolnych szyjnych
zgina kręgosłup i głowę w swoją stronę i obraca w stronę przeciwną, obustronnie prostuje g ł o w ę i kręgosłup
d) M m . podpotyliczne (mm. suboccipitales)
guzek tylny I, w y r o s tek kolczysty II kręgu szyjnego, wyrostki poprzeczne I kręgu szyjnego
wyrostki poprzeczne I kręgu szyjnego, kość potyliczna
zespół krótkich mięśni - prostują i obracają głowę
Mięśnie szyi M. szeroki szyi (m. plastysma)
skóra szyi na pograni- skóra w okolicy żuchczu klatki piersiowej w y
obniża ust
M. mostkowo-obojczykowo-sutkowy (m. sternocleidomastoideus)
m o s t k o w y koniec obojczyka i rękojeść mostka
wyrostek sutkowaty k. skroniowej
pochyla g ł o w ę w swoją stronę i skręca głowę w stronę przeciwną
k. gnykowa
obniża k. gnykową
chrząstka tarczowata
obniża krtań
M. mostkowo-gnykowy (m. sternohyoideus)
żuchwę
i
kąt
rękojeść mostka
M. mostkowo-tarczowy (m. sternothyreoideus) M. dwubrzuśćcowy (m. digastricus)
Wyrostek sutkowaty k. skroniowej
k. gnykowata trzon żuchwy
unosi k. g n y k o w ą i krtań, obniża żuchwę
M. r y l c o w o - g n y k o w y (m. stylohyoideus)
wyrostek rylcowaty k. skroniowej
k. gnykowa
unosi k. gnykową
M. żuchwowo-gnykowy (m. mylohyoideus)
wewnętrzna powierzchnia trzonu żuchwy
k. gnykowa
unosi k. gnykową, obniża żuchwę
M. b r ó d k o w o - g n y k o w y
kolec b r ó d k o w y żuch-
unosi k. gnykową, ob-
wy
niża żuchwę
(m. geniohyoideus) M m . pochyłe
(mm.
150
scaleni)
wyrostki poprzeczne kręgów szyjnych
żebra I i I
unoszą żebra I i II (pomocnicze mięśnie wdechowe), zginają i pochylają bocznie szyję
Mięsień
Przyczep początkowy
Przyczep k o ń c o w y
Czynność
Mięsnie g ł o w y
Mięśnie
żwaczowe
M. s k r o n i o w y (m. temporalis)
powierzchnia dołu skroniowego, powięź skroniowa, łuk jarzmowy
wyrostek żuchwy
M. żwacz (m. masseter)
d o l n y brzeg łuku jarzmowego
boczna powierzchnia kąta żuchwy na guzowatości żwaczowej
unosi żuchwę, dociska żuchwę do szczęki, nieco przesuwa ją do przodu
M. skrzydłowy przyśr o d k o w y (m. pterygoideus medialis)
dół skrzydłowy wyrośtka skrzydłowego kości klinowej
powierzchnia przyśrodkowa kąta żuchwy
unosząc żuchwę współdziała z skroniowym i czem
M. skrzydłowy boczny (m. pterygoideus lateralis)
powierzchnia podskroniowa podstawy czaszki, boczna blaszka wyrostka skrzydłowatego
wyrostek kłykciowy żuchwy
działając obustronnie w y s u w a żuchwę ku przodowi, jednostronnie obraca żuchwę ku stronie przeciwnej
Mięśnie
dziobiasty
unosi żuchwę, tylna mięśnia żuchwę
część cofa
mm: żwa-
mimiczne
M. policzkowy (m. buccinator)
wyrostek zębodołowy szczęki i część zęb o d o ł o w a żuchwy
M. obniżający w a r g ę dolną (m. depressor labii inferioris)
żuchwa szyi
M. obniżający kąt ust Im. depressor anguli orisj
przednia i boczna część żuchwy
M. potyliczno-czołowy (m. occipitofrontalis)
brzusiec potyliczny do kości potylicznej brzusiec czołowy w czepiec ścięgnisty pokrywający czaszkę skórze b r w i
wygładza zmarszczki czoła, przesuwając czepiec ścięgnisty do tyłu marszczy czoło, unosi brwi
część przyśrodkowa skóry brwi
marszczy czoło, unosi brwi
m. szeroki
M_m. marszćzący b r w i k. czołowa ponad (m. corrugator su- szwem czołowopercilii) szczękowym
kąt ust
pociąga kąt ust ku bokowi, przesuwa pokarm między zęby, zwiększa ciśnienie w jamie ustnej
warga dolna
obniża w a r g ę dolną
kąt ust
obniża kąt ust
151
Przyczep początkowy
Mięsień M. okrężny oka (m. orbicularis oculi)
M. okrężny ust
(m. orbicularis oris)
M m . j a r z m o w e większy i mniejszy (mm. zygomatici major et minor) M. ś m i e c h o w y
(m. risorius)
Przyczep k o ń c o w y
Czynność
otacza szparę powiekową, dzieli się na część powiekową i o b w o d o w o leżącą część oczodołową
cz. powiekowa opuszcza powiekę, cały mięsień szczelnie zamyka szparę powiekową i zwęża wejście do oczodołu
otacza szparę ustną, dzieli się na część brzeż-
cz. brzeżna zwiera w a r gi i przyciska je do zęb ó w , cz. w a r g o w a w y suwa zwarte w a r g i ku przodowi
ną i część w a r g o w ą
k. jarzmowa
kąt ust i warga górna
unosi kąt ust i w a r g ę górną
powięź
kąt ust
pociąga kąt ust ku bokowi
przyusznicza
skóra policzka Mięśnie kończyny g ó r n e j
Mięśnie
obręczy
M. naramienny (m. deltoideus)
boczny odcinek obojczyka, wyrostek barkowy, grzebień łopatki
M. nadgrzebieniowy
dół nadgrzebieniowy łopatki
(m.
supraspinatus)
kończyny górnej guzowatość naramienna k. ramiennej
zgina ramię, odwodzi ramię, prostuje i cofa ramię zwiera staw ramienny, zapoczątkowuje
od-
wodzenie ramienia M. podgrzebieniowy ^ (m. infraspinatus)
dół podgrzebieniowy łopatki
guzek większy k. ramiennej
wspomaga nie
odwodze-
ramienia,
obraca
ramię na zewnątrz M. obły mniejszy
brzeg boczny łopatki
• (m. teres minor) M. obły większy (m. teres major)
dolny kąt łopatki
grzebień guzka mniejszego
M. p o d ł o p a t k o w y
dół podłopatkowy
guzek mniejszy k. ramiennej, torebka sta-
Im.
subscapularis)
w u ramiennego
Mięśnie
kończyny
górnej
obraca ramię na zewnątrz, przywodzi ramię przywodzi ramię i obraca je do w e w n ą t r z przywodzi, obraca ram i ę do wewnątrz, napina torebkę stawu ramiennego
wolnej
Mięśnie ramienia M. d w u g ł o w y ramienia (m. biceps barchii)
152
głowa długa - guzek nadgrzebieniowy łopatki, głowa krótka wyrostek kruczy łopatki
guzowatość k. promieniowej, powięź przedramienia poprzez rozcięgno m. dwugłowego
zgina i odwraca przedramię głowa długa o d w o dzi ramię, krótka przywodzi i zgina ramię
Mięsień
Przyczep początkowy
M. ramienny (m. brachialis)
dolna część guzowatość k. łokcioprzedniej powierzchni w e j k. ramiennej
M. kruczo-ramienny wyrostek (m. coracobrachialis) patki
M. t r ó j g ł o w y ramienia (m. triceps brachii)
kruczy ło-
Przyczep k o ń c o w y
powierzchnia środkowa k. ramiennej
przyśtrzonu
głowa długa - guzek p o d p a n e w k o w y łopatki
Czynność zgina przedramię
zgina i przywodzi ramię
prostuje ramię w stawie łokciowym
g ł o w a b o c z n a - g ó r n a wyrostek ł o k c i o w y część tylnej powierz- k. łokciowej chni trzonu k. ramiennej g ł o w a przyśrodkowa dolna część tylnej powierzchni trzonu k. ramiennej
głowa długa dzi ramię
przywo-
Mięśnie przedramienia boczna powierzchnia nawraca przedramię, 1/ cz. k. zgina przedramię w 3 środkowej promieniowej stawie ł o k c i o w y m
M. n a w r o t n y obły (m. pronator teres)
M. zginacz promieniowy nadgarstka (m. flexor carpi radialis)
nadkłykiec przyśrodk o w y k. ramiennej
M. d ł o n i o w y długi (m. palmaris longus)
podstawa śródręcza
II i
III k.
rozcięgno d ł o n i o w e
zgina przedramię w stawie łokciowym, zgina rękę, napina rozcięgno d ł o n i o w e
k. grochowata, haczykowata i podstawa V
zgina i przywodzi rękę w stawie promienio-
k. śródręcza
wo-nadgarstkowym
nadkłykiec przyśrodkowy k. ramiennej M. zginacz nadgarstka ł o k c i o w y (m. flexor
carpi
ulnaris)
oraz wyrostek dziobiasty k. łokciowej
M. zginacz powierzchow n y palców (m. flexor digitorum superficialis)
nadkłykiec przyśrod- podstawy paliczków k o w y k. ramiennej, środkowych palców wyrostek dziobiasty k, II—V łokciowej, przednia powierzchnia k. promieniowej
M. zginacz długi kciuka
środkowa '/ 3 część trzonu k. promieniowej
(m. flexor pollicis longus)
zgina i nawraca przedramię, zgina i o d w o d z i rękę
podstawa dalszego paliczka kciuka
zgina rękę w stawie nadgarstkowym, zgina' palce w stawach śród ręczno-pa Icow y c h i międzypaliczkowych bliższych zgina kciuk
153
Mięsień
Przyczep początkowy
Przyczep k o ń c o w y
Czynność
M. zginacz głęboki palców (m. flexor digitorum profundus)
górna połowa trzonu k. łokciowej
podstawy dalszych paliczków I -V palców
zgina palce w stawach śródręczno-palićzkow y c h i międzypaliczkowych, zgina rękę w stawie p r o m i e n i o w o -nadgarstkowym
M. nawrotny czworoboczny (m. pronator ąuadratus)
dolna V 4 cz. trzonu k. łokciowej
dolna część przedniej powierzchni k. promieniowej
nawraca przedramię
wyrostek rylcowaty k. promieniowej
zgina przedramię w stawie ł o k c i o w y m , odwraca lub nawraca przedramię do położenia pośredniego z położeń krańcowych
grzbietowa powierzchnia podstawy II k. śródręcza
prostuje i o d w o d z i rękę w stawie promieniowo-nadgarstkowym
grzbietowa powierzchnia III k. śródręcza
odwodzi rękę w stronę promieniową, prostuje rękę
boczna i przednia powierzchnia k. promieniowej w górnej jej części
odwraca przedramię
M. prostownik palców (m. extensor digitorum)
rozcięgno grzbietowe II—V palców ręki
prostuje palce, prostuje rękę w stawie promieniowo-nadgarstkowym
M.
rozcięgno grzbietowe palca V
prostuje palec V, prostuje rękę w stawie pro-
M.
ramienno-promien i o w y (m. brachioradialis)
boczny brzeg dalszej przynasady k. ramiennej
M. prostownik nadgarstka promieniowy długi (m. extensor carpi radialis longus) M. prostownik nadgarstka promieniowy krótki (m. extensor carpi radialis brevis)
nadkłykiec boczny k. ramiennej
M. odwracacz
(m.
supinator)
prostownik piątego
palca
(m. extensor ęuinti)
digiti
mieniowo-nadgarstkowym
M. prostownik łokciowy nadkłykiec boczny k. nadgarstka ramiennej (m. extensor carpi ulnaris)
grzbietowa
chnia podstawy V k. śródręcza
rękę w stawie promieniowo-nadgarstkowym
M. ł o k c i o w y
wyrostek ł o k c i o w y
prostuje przedramię w stawie ł o k c i o w y m
(m.
154
anconeus)
powierz-
prostuje
i
przywodzi
Przyczep początkowy
Mięsień M. odwodziciel długi kciuka (m. abductor
pollicis
longus)
powierzchnia tylna błony międzykostnej, trzon k. p r o m i e n i o w e j
Przyczep k o ń c o w y podstawa ręczą
I k, śród-
Czynność prostuje
i
odwodzi
kciuk w stawie nadgarstkowo-śród ręcznym
i trzon k. łokciowej M.
podstawa paliczka bliższego kciuka
prostuje i odwodzi kciuk w stawie śródręczno-paliczkowym
M. prostownik długi powierzchnia tylna kciuka (m. extensor błony międzykostnej pollicis longus) i trzon k. łokciowej
podstawa paliczka dalszego kciuka
prostuje i nieco przywodzi kciuk
M. prostownik wskaziciela (m. extensor indicis)
rozcięgno grzbietowe palca wskazującego
prostuje palec wskazujący
prostownik krótki kciuka (m. exteńsor pollicis brevis)
Mięśnie ręki M. odwodziciel krótki kciuka (m. abductor brevis pollicis)
k. łódeczkowata
boczna strona bliższego paliczka kciuka
M. przeciwstawiacz kciuka (m. opponens pollicis)
k. czworoboczna większa
brzeg boczny I k. śród- przeciwstawia kciuk ręczą pozostałym palcom
M. zginacz krótki kciuka (m. flexor pollicis brevis) M. przywodziciel kciuka (m. adductor poilicis)
podstawa bliższego paliczka kciuka
głowa
poprzeczna -
powierzchnia dłoniowa III k. śródręcza, gło-
podstawa
bliższego
odwodzi kciuk
'zgina kciuk w stawie śródręczno-palcowym
przywodzi
paliczka kciuka
palca II
kciuk
do
wa skośna - k. główkowata M. odwodziciel palca małego (m. abductor digiti minimi)
k. grochowata
podstawa bliższegopaliczka palca małego
odwodzi palec mały od palca IV
M. zginacz krótki palca małego (m. flexor digiti minimi)
k. haczykowata
podstawa
M. przeciwstawiacz palca małego (m. opponens digiti minimi)
bliższego
zgina palec w stawie
paliczka palca małego
śródręczno-palcowym
brzeg ł o k c i o w y V k. śródręcza
przybliża palec mały i przeciwstawia go kciukowi
155
Mięsień M. d ł o n i o w y krótki (m. palmaris brevis)
Przyczep początkowy rozcięgno d ł o n i o w e
Przyczep k o ń c o w y
Czynność
skóra ponad kłębikiem marszczy skórę
M m . giistowate (mm. lumbricales)
ścięgna zginacza głębokiego palców
rozcięgna grzbietowa- zgina palce w stawach te palców II—V śródręczno-paliczkow y c h , prostuje w staw a c h międzypaliczkowych
M m . międzykostne grzbietowe (mm. interosse'. dorsales)
d w i e m a g ł o w a m i na d w ó c h sąsiednich kk. śródręcza
rozcięgna grzbietowe palców II—IV
odwodzi palce II i IV od palca III, odwodzi i przywodzi palec III zgina palce w stawach śródręczno-paliczkow y c h i prostuje w staw a c h międzypaliczkowych
rozcięgna grzbietowe palców II, IV i V
przywodzi palce II, IV i V do palca III, zgina stawy śródręczno-paliczkowe i prostuje międzypaliczkowe
M m . międzykostne dło- kk. śródręcza II, IV, V ni o w e (mm. interossei palmares)
Mięśnie kończyny dolnej
Mięśnie
obręczy kończyny
M. biodrowo-lędźwiowy (m. iliopsoas)
składa się z m. lędźw i o w e g o większego i m. b i o d r o w e g o
M. lędźwiowy większy
trzony i wyrostki poprzeczne od I do IV kręg ó w lędźwiowych
(m. psoas major) M. b i o d r o w y (m. iliacus)
krętarz udowej
dolnej mniejszy
k.
krętarz mniejszy kości u d o w e j
„
przy ustalonym udzie pochyla miednicę ku przodowi. zgina i obraca na zewnątrz udo
dół b i o d r o w y (wewnętrzna powierzchnia talerza biodrowego)
M. lędźwiowy mniejszy trzon XII kręgu piersio- powięź biodrowa, (m. psoas minor) w e g o i I lędźwiowego wyniosłość biodrowo-łonowa
zgina kręgosłup w odcinku l ę d ź w i o w y m
M. pośladkowy wielki (m. gluteus maximus)
tylna część talerza bio- guzowatość pośladd r o w e g o , k. krzyżowa, kowa k. udowej, prze-
prostuje, o d w o d z i i obraca na zewnątrz udo
więzadło -guzowe
w stawie b i o d r o w y m
krzyżowo-
M. pośladkowy średni (m. gluteus medius)
środkowa cz. zewnętrznej powierzchni talerza b i o d r o w e g o
M. pośladkowy mały (m. gluteus minimus)
przednia cz. zewnętrznej powierzchni talerza b i o d r o w e g o
156
groda międzymięśniowa boczna krętarz większy kości udowej
odwodzi, obraca na zewnątrz udo, cz. przednia mięśnia obraca udo do w e w n ą t r z
Mięsień
Przyczep początkowy
Przyczep k o ń c o w y
Czynność
M. gruszkowaty (m. piriformis)
powierzchnia miedniczna k. krzyżowej
przyśrodkowa powierzchnia krętarza większego
obraca udo nątrz
M.
zasłaniacz w e w nętrzny (m. obtura-
torius
internus)
M m . bliźniacze gemelli)
(mm.
M. czworoboczny uda (m. quadratus femoris)
na
zew-
powierzchnia w e w n ę trzna błony zasłonowej kolec kulszowy, guz kulszowy krętarz większy i grzebień międzykrętarzowy
guz kulszowy
M. zasłaniacz zewnęt- zewnętrzna powierzrzny (m. obturatorius chnia błony zasłonoexternus) wej
Mięśnie
dół krętarzowy, przyśrodkowa strona krętarza większego
kończyny
dolnej
wolnej
Mięśnie uda M. krawiecki (m. sartorius)
M. c z w o r o g ł o w y uda (m. quadriceps femoris) a) M. prosty uda (m. rectus femoris) b) M. obszerny boczny (m. vastus
kolec b i o d r o w y przedni górny
k. piszczelowa przyśrodkowo od guzowatości piszczeli
zgina, odwodzi i obraca na zewnątrz udo w stawie b i o d r o w y m
rzepka i za pośrednict w e m więzadła właś-
prostują staw kolanow y , ponadto m. prosty
c i w e g o rzepki do guzowatości k. piszcze-
zgina udo
kolec b i o d r o w y przedni dolny
kresa ch ropowa na k. udowej
lateralis)
lowej c) M. obszerny pośredni (m. vastus intermedius)
przednia powierzchnia trzonu k. udowej
d) M. obszerny przy- kresa chropawa na k. ś r o d k o w y (m. va- u d o w e j stus medialisj
M.
dwugłowy uda (m. biceps femoris)
a) głowa krótka kresa chropawa
g ł o w a strzałki
są s i l n y m i zginaczami s t a w u kolanowego.
157
Przyczep początkowy
Mięsień
Przyczep k o ń c o w y
b) głowa długa - guz kulszowy
Czynność przyczepiając
się
do
guza kulszowego prostują również staw biodrowy
guz kulszowy
k. piszczelowa poniżej guzowatości k. piszczelowej
M. smukły (m. gracilis)
dolna gałąź k. u d o w e j
przyśrodkowa powierzchnia piszczeli (gęsia stopa)
M. grzebieniowy
grzebień k. ł o n o w e j
górna część kresy
M. półścięgnisty (m. semitendinosus M. półbłoniasty (m. semimembranosus)
(m.
chropawej
pectineus)
M. przywodziciel krótki (m. adductor brevis)
przednia powierzchnia górnej gałęzi k. łonowej
środkowa cz. kresy chropawej
M. przywodziciel długi k. ł o n o w a poniżej guz(m. adductor longus) ka ł o n o w e g o M. przywodziciel wielki (m. adductor magnus)
przywodzi udo, zgina staw kolanowy •
przywodzą udo
dolna gałąź k. ł o n o w e j dolna cz. kresy chropawej, kłykieć przyśi k. kulszowej r o d k o w y k. u d o w e j Mięśnie goleni
M. piszczelowy przedni boczna powierzchnia (m. tibialis anterior) piszczeli, błona międzykostna, powięź goleni M.
prostownik długi kłykieć boczny piszpalców (m. extensor czeli, błona międzykodigitorum longus) stna, strzałka
M. strzałkowy trzeci (m. fibularis tertius)
m. strzałkowy trzeci - cz. prostownika długiego palców, kłykieć boczny piszczeli M. prostownik długi pa- błona międzykostna lucha (m. extensor i strzałka hallucis longus)
k. klinowata przyśrodkowa, podstawa I k. śródstopia
rozcięgna grzbietowe palców II—V
zgina grzbietowo, nawraca stopę, prostuje II—V palce
grzbietowa powierzchnia podstawy V k. śródstopia
zgina grzbietowo i nawraca stopę
podstawa dalszego paliczka palucha
prostuje paluch
guz piętowy poprzez ścięgno piętowe
zgina staw kolanowy, zgina podeszwowo
M. t r ó j g ł o w y łydki (m. triceps surae)
d w i e m a g ł o w a m i na
M. brzuchaty łydki (m. gastrocnemius)
tylnej powierzchni kły- (Achillesa) kci k. u d o w e j
158
zgina grzbietowo i odWraca supinuje (stopę)
i odwraca stopę
Mięsień
Przyczep początkowy
Przyczep k o ń c o w y
M. płaszczykowaty (m. soleus)
bliższe końce piszczeli i strzałki
M. podeszwowy (m. plantaris)
kłykieć boczny k. udowej
wraz ze ścięgnem pięt o w y m do guza piętowego
zgina stopę
M. p o d k o l a n o w y (m. popliteus)
kłykieć boczny k, udowej
k. piszczelowa p o w y żej przyczepu m.
zgina staw kolanowy, obraca piszczel do w e wnątrz przy zgiętym kolanie
zgina podeszwowo odwraca stopę *
płaszczkowatego
M. piszczelowy t y l n y (m. tibialis posterior)
Czynność
trzon piszczeli, strzałki k, łódkowata, kk. kiii błona międzykostna nowate, podstawy I I V kk. śródstopia
podeszwowo
zgina podeszwowo stopę, przywodzi przednią część stopy, odwraca stopę
M. zginacz długi palców
torum
(m. flexor digi-
longus)
tylna
powierzchnia
trzonu kości piszcze-
podstawy dalszych paliczków ii—V palców
lowej
zgina podeszwowo i odwraca stopę, zgina palce II—V
M. zginacz długi palucha (m. flexor hallucis longus)
strzałka, błona międzykostna, piszczel
podstawa paliczka dalszego palucha
zgina podeszwowo stopę, zgina paluch
M.
głowa i boczna powierzchnia strzałki
ścięgno przebiega skośnie pod podeszwą do k. klinowatej przyśrodkowej i I k. śródstopia
zgina
guzowatość V k. śródstopia
zgina podeszwowo i nawraca stopę
strzałkowy długi (m. peroneus longus)
M. strzałkowy krótki (m. peroneus brevis)
środkowa i dolna cz. strzałki
podeszwowo
i nawraca stopę, o d w o dzi przednią część sto-
PY
Mięśnie stopy M. prostownik krótki palucha
grzbietowa powierzchnia kości piętowej
(m. extensor hallucis brevis)
w okolicy wejścia do
M. prostownik krótki palców (m. extensor digitorum brevis)
grzbietowa powierzchnia k. piętowej
rozcięgno grzbietowe
prostuje paluch
palucha
zatoki stępu
M. odwodziciel palucha guz piętowy (m. abductor hallucis)
rozcięgna grzbietowe palców fl—V
prostuje palce II—V
trzeszczka przyśrodkowa i podstawa bliższego paliczka palucha
odwodzi paluch od palca II, wzmacnia sklepienie podłużne stopy
159
Mięsień
Przyczep początkowy
M. zginacz krótki
k. sześcienna, więzad-
Obie trzeszczki palu-
zgina paluch, wzmac-
ło podeszwowe dłu-
cha i podstawa bliż-
nia podłużne sklepie-
gie
szego paliczka palu-
nie stopy
palucha
(m. flexor brevis)
hallucis
Przyczep k o ń c o w y
Czynność
cha
M. przywodziciel palucha (m. adductor hallucis)
głowa skośna II—IV k. śródstopia, k. klinowata boczna, głowa poprzeczna - torebki s t a w ó w śródstopno-paliczkowych ll-V palca
trzeszczka boczna
M. odwodziciel palca małego (m. abductor digiti minimi)
k. piętowa i rozcięgno podeszwowe
boczna powierzchnia podstawy paliczka bliższego palca małego
o d w o d z i p f l c e V od palców IV, zgina palec m a ł y
M. zginacz krótki palca małego (m. flexor digiti minimi)
podstawa V k. śródstopią, więzadło podeszwowe długie
podstawa paliczka bliższego palca małego
zgina i odwodzi palec mały od palca IV
M. zginacz krótki palc ó w (m. flexor digitorum brevis)
guz p i ę t o w y i rozcięgno podeszwowe
rozczepione ścięgna na podstawie środkow e g o paliczka II—V palca
zgina palce, wzmacnia sklepienie podłużne stopy
M. czworoboczny podeszwy (m. quadratus plantae)
k. piętowa
boczny brzeg ścięgien zgina palce i wzmacnia podłużne zginacza długiego sklepienie stopy palców
M m . giistowate (mm. lumbricales)
ścięgna zginacza długiego palców
podstawa bliższego paliczka II—V palca
zginają palce w staw a c h śródstopno-paliczkowych, przywodzą palce do palucha
M m . międzykostne grzbietowe (mm. interossei dorsales)
sąsiadujące ze sobą kk. śródstopia
grzbietowe rozcięgna palców II—V
zginają stawy śródstopno-paliczkowe, prostują międzypaliczkowe i przywodzą palce do palucha
M m . międzykostne po-
kk. śródstopia III, IV i V podstawy bliższych paliczków palców II, III, IV i V
zginają stawy śródstopno-paliczkowe, prostują międzypaliczkowe i odwodzą palce od palucha
deszwowe (mm. interossei plantares)
160
wzmacnia poprzeczne sklepienie stopy, przywodzi paluch do palca
II
Fizjologia mięśni Dane ogólne Jak w i e m y , mięsień zbudowany jest z cienkich, długich komórek, zwanych włóknami mięśniowymi. Włókno mięśniowe ma długość od 1 mm do kilkunastu centymetrów, a jego grubość nie przekracza 100 urn. Włókno otacza błona k o m ó r k o w a ( s a r c o l e m m a ) . W dużym powiększeniu mikroskopowym widoczne jest poprzeczne prążkowanie włókna składające się z naprzemiennie ułożonych jasnych i ciemnych prążków. W obrębie cytoplazmy włókna mięśniowego, tzw. sarkoplazmy, znajdują się ułożone w osi długiej komórki włókienka mięśniowe (miofibryle), w których w mikroskopie elektronowym widoczne są jasne i ciemne prążki. Ciemne, podwójnie załamujące światło, prążki nazywamy a n i z o t r o p o w y m i (A). Prążki jasne noszą nazwę izotropowych i oznacza się je literą I. Dokładne badania struktury prążków mięśniowych pozwoliły stwierdzić, iż w środku prążków A znajduje się jaśniejsze pole H, a w środku prążka I leży ciemniejsza wstawka Z. Odcinek włókienka zawarty między d w o m a wstawkami Z tworzy jednostkę kurczliwą, zwaną sarkomerem. Podczas skurczu włókienka wstawki Z zbliżają się do siebie, a prążki I i H stają się węższe. Szerokość prążka A pozostaje nie zmieniona. Taki obraz staje się zrozumiały po prześledzeniu budowy włókienka mięśniowego.
Ryc. 97. Budowa mięśnia poprzecznie prążkowanego: 7 - s a r k o m e r , A - p r ą ż e k a n i z o t r o p o w y , /prążek izotropowy, Z- wstawka Z, H - pole H. 11 - Anatomia i fizjologia
161
Szczegółowe i żmudne badania wykazały, iż włókienko mięśniowe składa się z dwóch rodzajów białek - aktyny i miozyny. Białka mają postać długich nici ułożonych naprzemiennie - nić miozyny sąsiaduje z nicią aktyny i odwrotnie. Miozyna rozciąga się w obrębie prążka A, nici aktyny biegną od wstawki Z do granicy prążka (ryc. 98). Jak wynika z ryciny, podczas skurczu nici aktyny przesuwają się w stosunku do miozyny, przy czym dochodzi do skrócenia sarkomeru (odcinek Z - Z).
Przemiana materii i energii w mięśniach
0
Złożone produkty pokarmowe wprowadzone do ustroju ulegają przemianom na mniejsze cząsteczki na drodze hydrolizy i fosfolizy. Hydroliza polega na podzieleniu dLfżych cząsteczek na mniejsze z przyłączeniem do każdej z nich wodoru (H) lub grupy OH. Ten rodzaj zmian zachodzi w przewodzie pokarmowym (p. str. 208). W komórkach rozpad jest częściej związany z procesem fosfolizy. Cząsteczki dzielą siętu na mniejsze przyłączając kwas fosforowy (H3P04) w postaci H2PO3. „Paliwem" mięśni szkieletowych, wykorzystywanym jako czynnik energetyczny, jest glikogen. Mięśnie szkieletowe zawierają około 400 g glikogenu. Glikogen zużywa się podczas pracy mięśni i odtwarza z glukozy znajdującej się we krwi. Glikogen mięśniowy ulega fosfolizie do fosforanu glukozy, a następnie do dwóch cząsteczek kwasu pirogronowego (CH3CO COOH) z jednoczesnym wydzielaniem wysokoenergetycznych cząsteczek kwasu adenozynotrójfosforowego (ATP) (p. str. 224). Wwarunkach beztlenowych kwas pirogronowy nie rozpada się na C0 2 i H 2 0. Dzieje się tak, ponieważ koenzym niezbędny do dalszych przemian zostaje unieczynniony w zredukowanej formie po przyłączeniu dwóch atomów wodoru. W tym układzie kwas pirogronowy przechodzi w kwas mlekowy, odłączając jednocześnie od koenzymu wodór według schematu:
CH:,CO COOH kwas pirogronowy
•
koenzym
2H
-
CH.,CH/OH/COOH kwas mlekowy
1
koenzym
Nadmierne gromadzenie się kwasu mlekowego w mięśniach prowadzi do ich zakwaszenia, wyrażającego się bólem i zmęczeniem mięśni. Wwarunkach prawidłowych we krwi znajduje się 10 mg% kwasu mlekowego. Po ciężkiej pracy fizycznej jego poziom może wzrosnąć do 200 mg%. Zwiększony w czasie wypoczynku lub. wskutek nasilonego oddychania (hiperwentylacji) dopływ tlenu powoduje, że około V 5 kwasu mlekowego utlenia się do dwutlenku węgla i wody. Otrzymana z tej reakcji energia służy do przekształcenia pozostałej części kwasu mlekowego z powrotem w glukozę. 162
Przy dostatecznej podaży dochodzi do przemiany tlenowej. W jej przebiegu kwas pirogronowy przekształca się w dwutlenek węgla i wodę, z jednoczesnym uwolnieniem dużej ilości energii zawartej w ATP i fosforanie kreatyny (cykl kwasów trójkarboksylowych, czyli cykl Krebsa). Przemiany chemiczne dostarczające energii mięśniom obrazuje schemat: Glikogen mięśniowy
Kwas pirogronowy Przemiana tlenowa
Przemiana beztlenowa Kwas mlekowy
C0 2 + H 2 0
ATP
Energia
Energia
Jak wynika ze schematu, do skurczu mięśnia potrzebny jest wysokokaloryczny związek powstający w wyniku przemiany, a jest nim przede wszystkim ATP. Wiemy również, że czynnymi kurczącymi się białkami mięśni są miozyna i aktyna, z których zbudowane są miofibryle. Zachodzące reakcje można przedstawić następująco: : . > Aktynomiozyna aktyna + miozyna ^ aktyna + + miozyna + ATP — A D P + fosforany —^ADP + + fosforan kreatyny — kreatyna + ATP
Energia dostarczana w wyniku beztlenowej i tlenowej przemiany węglowodanów w mięśniach w 75% zamienia się w ciepło, a tylko w 25% może być przekształcona w energię mechaniczną, zatem wydajność „silnika mięśnioweg o " jest stosunkowo mała. Skurcz mięśnia Skurcz jest wyrazem stanu czynnościowego mięśnia. Mięsień kurcząc się zbliża z pewną siłą swe przyczepy i jeżeli jeden lub oba przyczepy mięśnia są ruchome, dochodzi do wzajemnego ich przemieszczania. Taki skurcz mięśnia nosi nazwę skurczu izotonicznego. Oznacza to, że napięcie mięśnia podczas skurczu izotonicznego nie zmienia się, natomiast zmienia się jego długość początkowa. Pobudzony mięsień, którego oba przyczepy s3 unieruchomione, zwiększa swoje napięcie, a jego długość nie ulega zmianie. Ten typ czynności mięśnia nazywamy skurczem izometrycznym. W ruchach dowolnych istnieje współdziałanie obu rodzajów czynności mięśni. Podniesienie dużego ciężaru zapoczątkowuje skurcz izometryczny i dopiero po przezwyciężeniu oporu dochodzi do skurczu izotonicznego. 163
Niekiedy możemy się również spotkać z innymi stanami czynnościowymi mięśni. Należą do nich: napięcie mięśniowe, skurcztężcowy (toniczny), skurcz kloniczny i inne. Napięciem mięśniowym nazywa się pewien stan pogotowia mięśnia do skurczu. Odróżnia się napięcie mięśni prawidłowe, wzmożone lub obniżone. Wzmożone napięcie mięśni występuje w niektórych chorobach ośrodkowego układu nerwowego, obniżone napięcie spotyka się u ludzi w wieku podeszłym prowadzących leżący lub siedzący tryb życia. Skurcz tężcowy, zwany niekiedy skurczem tonicznym, charakteryzuje się silnym i długotrwałym napięciem mięśnia; w swoim działaniu przypomina skurcz izometryczny. Skurcz kloniczny polega na serii szybkich skurczów mięśnia lub poszczególnych jego części. Przykładem skurczów klonicznych są np. dreszcze.
Pobudzenie mięśnia W warunkach doświadczalnych mięsień można pobudzić do skurczu czynnikami mechanicznymi, chemicznymi i za pomocą prądu elektrycznego. W ustroju pobudzenie mięśni następuje za pośrednictwem nerwu ruchowego, kończącego się na włóknie mięśniowym tzw. płytką końcową. Zwykle jedno włókno nerwowe, pochodzące od komórki ruchowej rogów przednich rdzenia, kończy się na kilku komórkach mięśniowych. Czynnościowa całość złożona
—- Włókno odśrodkowe (neuryt)
V
•Włókna mięśniowe
Ryc. 98. Schemat jednostki ruchowej.
164
z komórki ruchowej (neurytu) włókna nerwowego i unerwionych przez nią włókien mięśniowych nazywa się jednostką ruchową (ryc. 98). Impuls biegnący nerwem ruchowym uwalnia w jego zakończeniu - płytce ruchowej - acetylocholinę, która przenosi pobudzenie z płytki końcowej na włókno mięśniowe. W ten sposób acetylocholina tworzy pomost między nerwem a mięśniem. Zapoczątkowuje ona depolaryzację elektryczną włókna mięśniowego, która poprzedza jego skurcz. Depolaryzacja wyraża się zmianą potencjału elektrycznego na powierzchni i wewnątrz komórki mięśniowej. W spoczynku wnętrze komórki jest ujemne w stosunku do środowiska zewnętrznego, komórka jest zatem spolaryzowana poprzecznie. Pobudzenie włókna mięśniowego do skurczu przez acetylocholinę powoduje zwiększenie przepuszczalności błony komórkowej przede wszystkim dla jonów potasu i sodu. Dochodzi do wyrównania potencjałów, czyli następuje depolaryzacja włókna mięśniowego. Powrót do stanu wyjściowego nazywa się repolaryzacją (p. str. 167). Odpowiedź mięśnia na stymulację elektryczną nerwu można zbadać na preparacie nerwowo-mięśniowym. Najprostszą jego postacią jest mięsień trójgłowy łydki żaby wraz z odcinkiem unerwiającego go nerwu ruchowego. Mięsień zamocowujemy jednym końcem w uchwycie, drugi łączymy z dźwignią kimografu. Nerw pobudzamy za pomocą elektrod włączonych we wtórny obwód cewki lub też przy użyciu prądu z baterii (ryc. 99). Badanie rozpoczynamy od bardzo słabych impulsów nerwowych. Nie powodują one skurczu mięśnia i nazywamy je bodźcami podprogowymi. Dalsze powiększanie siły bodźca doprowadza po pewnym czasie (zwanym
Ryc. 99. Schemat skurczu mięśnia pobudzonego prądem elektrycznym.
165
okresem utajonego pobudzenia) do drgnięcia mięśnia. Bodziec wywołujący taki skurcz nazywa się bodźcem p r o g o w y m (tzw. reobaza). Pobudzenie mięśnia bodźcem progowym powoduje skurcz tylko niektórych jego włókien. Dalsze wzmacnianie siły prądu powoduje kurczenie się większej liczby włókien mięśniowych należących do różnych jednostek motorycznych. Powodują to bodźce nadprogowe. Przy odpowiednio dużej sile bodźca wszystkie włókna badanego mięśnia kurczą się i dalsze zwiększanie siły prądu pozostaje bez wpływu na siłę rozwijaną przez kurczący się mięsień. Dzieje się to przy zastosowaniu bodźców supramaksymalnych. Ciekawe zjawiska możemy zauważyć w przypadku stosowania dwóch impulsów elektrycznych następujących po sobie. Efekt zależy od czasu, który upływa pomiędzy pierwszym a drugim impulsem. Jeżeli okres między kolejnymi pobudzeniami jest dostatecznie długi, obserwujemy dwa identyczne skurcze mięśni. Z kolei jeżeli czas między pobudzeniami jest bardzo krótki (rzędu kilku milisekund), drugi skurcz w ogóle nie wystąpi, ponieważ bodziec trafił na okres niewrażliwości-tzw. okres refrakcji mięśnia. Pomiędzy dwoma przedstawionymi tu krańcowymi sytuacjami drugi bodziec sumuje się z pierwszym, w różnym stopniu powodując wzmożenie skurczu. Po zastosowaniu serii bodźców o umiarkowanej częstotliwości występuje skurcz kloniczny. Zwiększenie częstotliwości bodźców powoduje zwiększenie siły skurczu mięśnia i wywołuje skurcz tężcowy (toniczny). Współzależności między siłą bodźca, czasem jego trwania i czasem, j a k i upływa od chwili zadziałania bodźca do wystąpienia skurczu, są wykorzystywane w chronaksjometrycznym badaniu mięśni. Czasem chronaksji mięśni nazywa się okres utajonego pobudzenia po zastosowaniu bodźca o sile podwójnej wartości bodźca progowego (podwójnej reobazy) (ryc. 99).
Zmęczenie Po wielokrotnie powtarzanym pobudzaniu preparat nerwowo-mięśniowy ulega zmęczeniu. Okres utajonego pobudzenia staje się dłuższy, skurcz słabszy, rozkurcz trwa długo, przy czym mięsień trudno odzyskuje swoją długość początkową. W preparacie nerwowo-mięśniowym wszystkie włókna mięśniowe są pobudzane synchronicznie, co nie odpowiada ściśle bodźcom wysyłanym przez komórki nerwowe rdzenia i dlatego wyniki tych doświadczeń mają charakter orientacyjny w poznaniu czynności mięśni. W zmęczonym mięśniu zwiększa się zawartość wody, mięsień jest obrzmiały i skrócony. W mięśniu takim podwyższony jest poziom kwasu mlekowego i CO2, a obniżony poziom glikogenu. Systematyczny trening przyczynia się do lepszego przystosowania mięśni do ciężkiej pracy fizycznej. Najbardziej typowym przejawem treningu jest zwiększanie się masy mięśnia. Znacznie bardziej niż objętość zwiększa się wytrzymałość ćwiczonych mięśni. Przerwa w treningu powoduje obniżenie 166
masy mięśniowej i wydolności mięśni, ubytek ten jest jednak stosunkowo powolny. Obserwowano, iż po roku od zaprzestania ćwiczeń można jeszcze stwierdzić wydolność mięśni większą niż wyjściową.
Zjawiska elektryczne w mięśniu Pobudzenie włókna mięśniowego przez acetylocholinę, wytwarzającą się w płytce końcowej, powoduje zaburzenie równowagi elektrycznej włókna. Jak wiemy, komórka mięśniowa, podobnie jak inne komórki ustroju, jest spolaryzowana. Wnętrze komórki jest ujemne w stosunku do powierzchni zewnętrznej błony komórkowej, która jest dodatnia. W cytoplazmie komórki znajdują się białka zawierające potas, które rozpadają się na dodatni jon potasu K + i ujemne jony reszty białkowej. Dodatnie jony potasu mogą przechodzić przez błonę komórkową na zewnątrz, podczas gdy ujemne wielkocząsteczkowe jony białka pozostają w komórce. W płynie pozakomórkowym znajduje się zdysocjowany chlorek sodowy, przy czym zarówno jon Na + , jak i ujemny Cl" mogą przenikać przez barierę błony komórkowej do wewnątrz komórki i na zewnątrz. W wari^nkach równowagi spoczynkowej duże stężenie jonów K + w komórce powoduje, iż jony gromadzą się przy błonie komórkowej i przechodzą przez nią na zewnątrz. Wskutek utraty jonów K + wnętrze komórki staje się ujemne w stosunku do płynu pozakomórkowego. Równowagę tę można ująć następująco: K+ Stężenie w komórce wysokie
+
Cl" Stężenie w komórce niskie
K+ Stężenie poza komórką niskie Innymi słowy:
K + poza komórką K+ w komórce
+
cr Stężenie poza komórką wysokie
Cl" w komórce Cl~ poza komórką
Pobudzenie włókna mięśniowego polega na zwiększeniu przepuszczalności błony komórkowej dla jonów K + i Na + . W miejscu pobudzenia jony K + wydostają się na zewnątrz, zaś jony Na + wnikają do wnętrza komórki. Dochodzi do zniknięcia układu polaryzacyjnego, przy czym ładunki na zewnątrz i wewnątrz komórki wyrównują się. Powrót do warunków wyjściowych zróżnicowania ładunków elektrycznych - nazywa się repolaryzacją. Wahnięcia potencjałów mogą być zarejestrowane - ich zapis nosi nazwę elektromiogramu. Elektromiogram pojedynczego włókna składa się z krzywej ilustrującej depolaryzację i repolaryzację. Układ ramion krzywej m ó w i o prawidłowej lub nieprawidłowej czynności bioelektrycznej włókna. W badaniach klinicznych wykonuje się elektromiogramy pochodzące od wielu włókien mięśniowych, stąd też zapis takiego elektromiogramu jest wysoce złożony. 167
UKŁAD T R A W I E N N Y
Przewód pokarmowy ma za zadanie dostarczyć organizmowi materiału do budowy i odnowy własnych tkanek oraz substancji energetycznych dla podtrzymania wszelkich procesów życiowych. Podstawowe substancje doprowadzane w postaci pokarmu pod względem chemicznym dzielą się na trzy duże grupy - węglowodany, tłuszcze i białka. Ponadto z pokarmem dostarczane są witaminy, sole mineralne i woda. Węglowodany i tłuszcze są głównym źródłem energii. Białka są przede wszystkim źródłem aminokwasów - materiału niezbędnego do budowy tkanek i narządów. Przy nadmiarze pokarmu białkowego lub przy braku węglowodanów i tłuszczów aminokwasy ulegają dezaminacji i mogą być zużyte dla celów energetycznych. Energia zawarta w pokarmach pochodzi w końcowym efekcie od roślin zielonych obdarzonych zdolnością zatrzymywania i gromadzenia fotonów promieniowania słonecznego w procesie fotosyntezy - pierwotnym źródłem wszelkiej energii organizmów żywych na ziemi jest zatem słońce. Pokarm pobierany przez człowieka z otoczenia ma przeważnie postać nie pozwalającą na bezpośrednie wykorzystanie go przez ustrój. Zespół pfbcesów, którym zostaje on poddany w przewodzie pokarmowym, nazywamy trawieniem. Trawienie składa się z procesów mechanicznych i chemicznych.
Ryc. 100. Budowa i funkcja przewodu pokarmowego (wg Vandera).
168
Do procesów mechanicznych należą żucie, potykanie, ruchy robaczkowe przełyku, ruchy żołądka i jelit, oddawanie stolca. Trawienie chemiczne polega na przekształcaniu dużych cząsteczek złożonych substancji organicznych w związki prostsze, które mogą przenikać przez błony komórkowe i które organizm może użyć do budowy własnych tkanek oraz do podtrzymania procesów życiowych związanych z wydatkowaniem energii. W wyniku tych przemian węglowodany zostają rozłożone na cukry proste,
Ślinianka podżuchwowa
—Żołądek Wątroba Pęcherzyk żółciowy Dwunastnica
Trzustka Okrężnica poprzeczna Jelito czcze Okrężnica .- itępująca
Okrężnica wstępująca
Jelito ślepe Wyrostek — robaczkowy
Esica Odbytnica Ryc. 101. Budowa przewodu pokarmowego.
169
głównie glukozę i fruktozę; tłuszcze na glicerol i kwasy tłuszczowe, białka - na aminokwasy. W żywym ustroju, procesy te zachodzą w temperaturze ciała dzięki swoistym katalizatorom produkowanym przez gruczoły trawienne. Noszą one nazwę e n z y m ó w . Trawienie odbywa się w obrębie przewodu pokarmowego. Ma on kształt długiej cewy, rozpoczynającej się szparą ustną i kończącej się odbytem. Do układu tego należą: jama ustna, gardło, przełyk, żołądek, jelito cienkie, jelito grube oraz wielkie gruczoły trawienne - wątroba i trzustka.
BUDOWA ŚCIAN PRZEWODU POKARMOWEGO Ścianę przewodu pokarmowego tworzą trzy zasadnicze warstwy: 1) błona śluzowa, 2) błona mięśniowa-i 3) błona surowicza. 1. Błona śluzowa (tunica mucosa) stanowi warstwę wyściełającą powierzchnię wewnętrzną przewodu pokarmowego. Pod błoną śluzową leży warstwa łącznotkankowa, zwana tkanką podśluzową (telasubmucpsa). Obecność tkanki podśluzowej sprawia, że błona jest w stanie układać się w fałdy i wygładzać zależnie od rozciągania lub kurczenia się ściany narządu. Na granicy błony śluzowej i tkanki podśluzowej występuje warstwa mięśni gładkich, zwana blaszką mięśniową błony śluzowej. 2. Błona mięśniowa (tunica muscularis) jest następną warstwą przewodu pokarmowego. Składa się zwykle z dwóch warstw włókien mięśniowych: wewnętrznej - okrężnej i zewnętrznej - podłużnej. Są to włókna mięśniowe gładkie. Pod względem czynnościowym różnią się one od mięśni poprzecznie prążkowanych t y m , że: 1) skurcz ich odbywa się znacznie wolniej i trwa dłużej; stan napięcia (tonus) mięśni gładkich utrzymuje się nawet po oddzieleniu ich od ośrodkowego układu nerwowego, 2) mają większą rozciągliwość, 3) obdarzone są zdolnością rytmicznych skurczów, 4) są bardziej wrażliwe na bodźce termiczne, chemiczne i rozciąganie, a mniej na bodźce elektryczne. 3. Błona surowicza (tunica serosa) jest cienką, gładką, białawą błoną, która pokrywa powierzchnię z e w n ę t r z ^ narządów, zwróconą do wnętrza jamy brzusznej. Nazywamy ją otrzewną (peritoneum).
J A M A USTNA Jama ustna (cavum oris) jest początkowym odcinkiem przewodu pokarmowego. Ograniczają ją: od góry - podniebienie, po bokach - policzki, od dołu dwa mięśnie żuchwowo-gnykowe, tworzące tzw. przeponę jamy ustnej; ścianę przednią jamy ustnej tworzą wargi, ograniczające szparę ustną; w tyle jama ustna łączy się z gardłem otworem zwanym cieśnią gardzieli. Łuki zębowe górny i dolny dzielą jamę ustną na dwie części, mające kształt szpary: na przedsionek j a m y ustnej, zawarty między łukami zębowymi a wargami i policzkami, oraz na znacznie większą jamę ustną właściwą. . 170
Zatoka czołowa Przegroda nosowa
Zatoka klinowa
Podniebienie twarde Podniebienie miękkie Język Migdałek językowy
Żuchwa
Nagłośnia K gnykowa Chrząstką tdrczowdtrf
Tchawica—
- —Chrząstka } pierścieniowata Przełyk
Ryc. 102. Przekrój strzałkowy przez jamę ustną, gardło, krtań i przełyk.
Szparę ustną ograniczają w a r g i : górna (labium superius) i dolna (labium inferius), łączące się ze sobą w kącie ust. Zrąb warg stanowi mięsień okrężny ust (m. orbicularis oris). Zrębóm policzka (bucca) jest mięsień Doliczkowy (m.buccinator), pokryty od zewnątrz skórą, od wewnątrz - błoną śluzową. Napięcie mięśni policzkowych utrzymuje policzki przy łukach zębowych. Z w a r g i policzków błona śluzowa przechodzi pod kątem ostrym na wyrostki zębodołowe, gdzie zrasta się z okostną, otacza szyjki zębowe oraz pokrywa przestrzenie międzyzębowe tworząc dziąsła (gingivae).
PODNIEBIENIE Podniebienie (palatum) ma kształt łukowatego sklepienia, wypukłego ku górze, oddzielającego jamę ustną od jamy nosowej. Część przednią podniebienia, utworzoną przez wyrostki podniebienne szczęk i blaszki pbziome kości 171
podniebiennych wraz z błoną śluzową i zawartymi w niej t w o r a m i , nazywamy podniebieniem t w a r d y m (palatum durum); część tylna, składająca sięzblasz"kilącznotkankowej, mięśni i pokrywającej je śluzówki, nosi nazwę podniebienia miękkiego (palatum molle). Zwisająca ku tyłowi część podniebienia miękkiego jest wyciągnięta w linii środkowej w postaci wyrostka zwanego języczkiem podniebiennym (uyula). W kierunku bocznym i ku dołowi odchodzą od podniebienia miękkiego dwa łuki podniebienne: przedni - łuk podniebienno-językowy-zdążą od bocznego brzegu języka, t y l n y - ł u k p o d n i e b i e n n o - g a r d ł o w y - biegnie do błony mięśniowej gardła. Pomiędzy łukiem przednim a t y l n y m każdej strony znajduje się duże zagłębienie - zatoka migdałkowa (sinus tonsillaris), w której znajduje się duże skupienie tkanki limfatycznej, zwane migdałkiem podniebiennym (tohsilla palatina). Migdałek ten o kształcie jajowatym ma wielkość zmienną - może wystawać spoza łuku podnjebienno-językowego, bądź też być schowany za nim prawie całkowicie.
ZĘBY Zadaniem uzębienia jest chwytanie, rozdrabnianie i miażdżenie pokarmu. U człowieka występują dwa pokolenia zębów - zęby mleczne (dentes decidui) i zęby stałe (dentes permanentes). W skład uzębienia stałego wchodzą 32 zęby, ułożone w dwa łuki.zębowe - górny i dolny po 16 zębów w każdym. W każdym zębie rozróżniamy koronę (corona dentis), szyjkę (collum dentis) i korzeń (radix dentis). Korona wystaje do jamy ustnej i jest właściwym elementem mechanicznym zęba. Korzeń jest połączony z zębodołem za pomocą silnej błony łącznotkankowej, zwanej ozębną (periodontium). Pomiędzy koroną a korzeniem znajduje się szyjka zęba. Korona zęba jest bardzo twarda i ma zabarwienie mlecznobiałe. Zabarwienie oraz twardość zawdzięcza w a r s t w i e okrywającej ją substancji, zwanej szkliwem (enamelum). Korzeń pokrywa warstwa o zabarwieniu lekkożółtaw y m , zwana k o s t n i w e m (cementum). Szyjkę osłania wspomniane wyżej dziąsła Na koronie każdego zęba rozróżniamy powierzchnię zgryzu, bądź też brzeg sieczny, powierzchnię przedsionkową lub twarzową, powierzchnię językową i dwie powierzchnie styczne, zwrócone do sąsiednich zębów przednią i tylną, inaczej boczną i przyśrodkową. Wnętrze korony zęba zajmuje komora zęba (cavum dentis), która przechodzi w kanał korzenia (canalis radicis), kończący się o t w o r e m szczytowym zęba (foramen apicis dentis). Komorę zębową i kanał korzenia wypełnia miazga zębowa (pulpa dentis). Trzon każdego zęba zbudowany jest z zębiny (dentinum), pod względem b u d o w y odmiennej od tkanki kostnej. Uzębienie człowieka jest zróżnicowane - zęby przednie, służące tylko do odgryzania pokarmów, mają kształt inny niż przystosowane do przeżuwania pokarmów zęby tylne, w obrębie których ciśnienie wywierane przez łuki 172
Guzek policzkowy
— Guzek językowy
- Kanał korzenia zęba
Korzeń
Otwór szczytowy zęba Ryc. 103.
Szlif
podłużny zęba.
Kolec nosowy przedni Powierzchnia policzkowa
Powierzchnia wargowa Powierzchnia boczna Powierzchnia policzkowa
— Ząb mądrości
Zęby sieczne Kieł
Zęby przedtrzonowe Otwór bródkowy
Zęby trzonowe
Ryc. 104. Lewa połowa uzębienia stałego (wg Toldta). 173
zębowe jest znacznie większe. W szeregu g ó r n y m i dolnym rozróżniamy z każdej strony następujące rodzaje zębów: 2 zęby sieczne (dentes incisM), ! kieł (dens caninus), 2 zęby przedtrzonowe (dentes premolares) i 3 t r z o n o w e (dentes molares), co schematycznie przedstawiamy za pomocą tzw. wzoru zębowego: M P C I 3 2 1 2
I C P M 2 1 2 3
3 2 1 2
2 1 2 3
W praktyce dentystycznej zęby oznacza się kolejnymi liczbami, poczynając od linii pośrodkowej, i zapisuje następująco: ząb przedtrzonowy górny drugi prawy 51 kieł górny Iewvl3 siekacz przyśrodkowy dolny prawy Tl ząb trzonowy dolny lewy trzecifśfitd. Zęby sieczne (dentes incisivi) mają koronę w kształcie dłuta; jest ona silnie spłaszczona od przodu ku tyłowi, na skutek czego powierzchnia żucia przybiera postać wąskiej i ostrej krawędzi siecznej. Powierzchnia w a r g o w a jest wypukła, powierzchnia językowa wklęsła. Korzeń siekaczy jest prawie dwa razy dłuższy niż korona. Siekacz górny przyśrodkowy jest największy; siekacz dolny przyśrodkowy - najmniejszy. Kieł (dens caninus) jest zębem najdłuższym; korona jego ma kształt stożkowaty, nie ma więc na niej krawędzi siecznej, lecz tylko punktsieczny. Korzeń kła jest blisko trzy razy dłuższy od jego korony. Korona zębów przedtrzonowych albo d w u g u z k o w y c h (dentes premolares) wykazuje na powierzchni żującej rowek przebiegający strzałkowo, który oddziela dwa guzki. Zęby te mają jeden korzeń spłaszczony od przodu ku t y ł o w i ; na korzeniu widnieje podłużny rowek. W szczęce mają niekiedy dwa korzenie.
Zęby sieczne przyśrodkowy
Kieł
I II Zęby przedtrzonowe
II III Zęby trzonowe
Ryc. 105. Zęby stałe. Widok z boku lub od tyłu (wg Todta).
174
Zęby t r z o n o w e (dentes molares) na powierzchni korony mają po" kilka guzków, toteż bywają nazywane zębami wieloguzkowymi. Najmasywniejszy jest ząb trzonowy pierwszy, zarówno górny, jak i dolny. Korona pierwszego zęba trzonowego górnego ma kształt okrągławy i wykazuje na powierzchni żucia cztery guzki. Drugi ząb trzonowy górny ma cztery, a często tylko trzy guzki. Trzeci ząb trzonowy górny (ząb mądrości) jest bardzo zmienny osobniczo zarówno pod względem kształtu, jak i wielkości. Zęby trzonowe dolne są cztero- lub pięcioguzkowe. Trzeci ząb trzonowy dolny, zwany również zębem mądrości, podobnie jak górny, jest bardzo zmienny. Zęby trzonowe górne mają po trzy korzenie: dwa po stronie policzkowej, jeden po stronie językowej; dolne - po dwa korzenie: przedni i tylny. Krzywizny obu łuków zębowych nie są jednakowe. Łuk górny ma kształt połowy elipsy, dolny - paraboli.
przyśrodkowy i boczny Ryc. 106. Zęby mleczne.
Powstająca w ten sposób różnica krzywizn zostaje częściowo wyrównana dzięki temu, że zęby górne są odchylone nieco ku przodowi, zęby dolne - ku tyłowi. Korony zębowe obu łuków są przesunięte względem siebie w ten sposób, że każdy ząb łuku górnego styka się z d w o m a zębami łuku dolnego i odwrotnie. Wyjątek stanowią dolne zęby sieczne przyśrodkowe r trzeci trzonowy górny, mające tylko po jednym antagoniście. Antagonistami nazywają się stykające się ze sobą zęby obu łuków. W skład uzębienia mlecznego wchodzi 20 zębów: dwa sieczne, jeden kieł i dwa trzonowe w każdej połówce obu łuków. Schematycznie przedstawiamy to następującym wzorem: 2 1 2 2 1 2
2 1 2 2 1 2 175
Poszczególne zęby mleczne mają kształty zbliżone do zębów stałych; różnią się nieco od nich zabarwieniem, które jest niebieskawobiałe. Przeciętne terminy wyrzynania się poszczególnych zębów przedstawia tab. 5. Należy przy t y m zaznaczyć, że od tych przeciętnych t e r m i n ó w mogą się zdarzać rozmaite odchylenia w obie strony, jak również, że zęby mogą wyrzynać się w innej kolejności.
Tabela 5 Okresy wyrzynania się zębów Zęby mleczne
Zęby stałe
Zęby sieczne przyśrodkowe
6 - 8 mies.
Zęby sieczne boczne Zęby trzonowe pierwsze (przednie)
7-12 mies. 12-16 mies.
Kły
15-20 mies.
Zęby trzonowe drugie (tylne)
20-30 mies.
zęby trzonowe pierwsze (przednie) zęby sieczne przyśrodkowe zęby sieczne boczne
zęby przedtrzonowe pierwsze (przednie) kły zęby przedtrzonowe drugie (tylne) zęby trzonowe drugie (tylne) zęby trzonowe trzecie (zęby mądrości)
6- 7 lat 7- 8 lat 8- 9 lat
9-11 lat 11-13 lat 11-13 lat 12-14 lat 17-25 lat
JĘZYK Język n y m na linguae). widnieje
(lingua) jest w a ł e m m i ę ś n i o w y m pokrytym swoistą śluzówką, położodnie jamy ustnej. Główna masa języka stanowi jego trzon (corpus Na powierzchni górnej, stanowiącej grzbiet języka (dorsum linguae), bruzda pośrodkowa.
Błona śluzowa języka Błonę śluzową grzbietu języka pokrywają liczne brodawki językowe, które sprawiają, że powierzchnia górna języka jest szorstka i ma w y g l ą d aksamitu. Błona śluzowa dolnej powierzchni języka jest cienka, gła"dka, delikatna, bez brodawek. Brodawki językowe (papillae linguales) są to wyrostki tkanki łącznej, które wystają ponad powierzchnię błony śluzowej i są pokryte nabłonkiem wielowar s t w o w y m płaskim. U człowieka rozróżniamy cztery rodzaje brodawek: 1) nitkowate, 2) grzybowate, 3) okolone i 4) liściaste. 176
Koniec języka
Ryc. 107. Język.
Mięśnie języka Zrąb języka utworzony jest przez mięśnie, prążkowane. Dzielimy je na dwie grupy: 1) mięśnie zewnętrzne, łączące się z otoczeniem, które rozpoczynają się poza językiem i kończą w jego trzonie, oraz 2) mięśnie własne języka. Do mięśni zewnętrznych należą: 1. Mięsień bródkowo-językowy (m. genioglossus)-mięsień, który pociąga język ku przodowi i dołowi. 2. Mięsień rylcowo-językowy (m. styloglossus) - jest antagonistą mięśnia poprzedniego - pociąga język ku tyłowi i ku górze. 3. Mięsień g n y k o w o - j ę z y k o w y (m. hyoglossus) - pociąga język ku dołowi i do tyłu. Wśród mięśni własnych języka rozróżniamy: dwa mięśnie podłużne języka - górny i dolny (m. longitudinalis linguae superior et interior), mięsień poprzeczny języka (m. transversus linguae) i mięsień p i o n o w y języka (m. verticalis linguae). Pęczki tych mięśni przebiegają w masie języka w trzech prostopadłych do siebie kierunkach. Skurcze mięśni własnych języka zmieniają jego kształt i w y m i a r y .
12 - Anatomia i fizjologia
177
GRUCZOŁY ŚLINOWE W jamie ustnej znajduje się szereg gruczołów ślinowych. Rozróżniamy gruczoły ślinowe małe i duże. Do dużych, zwanych także śliniankami, należą: ślinianka przyuszna (glandula parotis), ślinianka p o d ż u c h w o w a (gl. submandibularis) i ślinianka podjęzykowa (gl. sublingualis). Małe gruczołki ślinowe są rozsiane w ścianach jamy ustnej i, zależnie od okolicy, w jakiej występują, rozróżniamy w ś r ó d nich: gruczoły w a r g o w e , policzkowe, językowe, podniebienne.
Ślinianka przyuszna Ślinianka przyuszna (glandula parotis) jest największym gruczołem ślinowym (ciężar 20-30 g), konsystencji miękkiej, barwy szaroróżowej. Leży na zewnętrznej powierzchni mięśnia żwacza pod powięzią przyuszniczą. Zwężona część ślinianki,' zwana częścią głęboką, wciska się w głąb dołu leżącego za gałęzią żuchwy. W całości ślinianka przyuszna ma kształt trójściennego klina o trzech powierzchniach: bocznej, przedniej i tylnej. Przewód ślinianki przyusznej (ductus parotideus) biegnie ku przodowi i uchodzi do przedsionka jamy ustnej, kończąc się m a ł y m o t w o r e m na wzniesieniu błony śluzowej, zwanym brodawką przyuszniczą (papilla parotidea), leżącym na wysokości drugiego górnego zęba trzonowego. Długość przewodu wynosi 40-60 m m , średnica 3 - 4 m m .
Ślinianka podżuchwowa Ślinianka p o d ż u c h w o w a (glandula submandibularis), mniejsza od poprzedniej (ciężar 10-15 g), ma kształt spłaszczonego jaja. Leży w okolicy podżuchwowej, częściowo pokryta żuchwą. Dolna część gruczołu zajmuje okolicę pomiędzy żuchwą a kością gnykową. Przewód ślinianki podżuchwowej, długości około 5 cm, biegnie ku przodowi otwiera się na mięsku podjęzykowym bocznie od wędzidełka języka.
Ślinianka podjęzykowa Ślinianka podjęzykowa (glandula sublingualis), najmniejsza spośród wszystkich gruczołów ślinowych (ciężar 3 - 5 g), leży pod błoną śluzową jamy ustnej, przylegając do dołka językowego żuchwy. Składa się z kilku zrazików i ma kilka (5-8) przewodów mniejszych, które otwierają się wzdłuż fałdu podjęzykowego, oraz jeden główny przewód większy długości 37-45 m m , otwierający się na mięśniu podjęzykowym, najczęściej razem z przewodem ślinianki podżuchwowej. 178
Komórki surowicze
Komórki — koszyczkowe
Ryc. 108. Budowa ślinianki.
GARDŁO Gardło (pharynx) ma kształt lejkowatej mięśniowo-włóknistej spłaszczonej cewy, położonej w tyle za jamą nosową, jamą ustną i krtanią, a przed trzonami kręgów szyjnych. Rozciąga się od podstawy czaszki do dolnego brzegu krtani, tj. do poziomu VI kręgu szyjnego, gdzie przechodzi w przełyk. Długość gardła wynosi 12-14 cm. Włóknista*warstwa ściany gardła jest mocno przytwierdzo na do podstawy czaszki. Górna część gardła ma kształt lejka, który ku dołowi stopniowo przybiera postać spłaszczonego cylindra, przy czym ściana przednia i tylna zbliżają $ię ku sobie, tak że na poziomie wejścia do krtani stykają się one ze sobą, tworząc wąską szparę, która rozszerza się w czasie przełykania pokarmu. Jamę gardła (cavum pharyngis) dzielimy na trzy części: a) górną - nosową (zwaną też jamą nosowo-gardłową), b) część środkową - ustną i c) część dolną -krtaniową. Gardło ma szereg o t w o r ó w , które łączą jamę gardła z jamami sąsiednimi. Z jamą nosową łączą je dwa nozdrza tylne, z jamą bębenkową z każdej strony symetryczne ujścia gardłowe trąbek słuchowych, z jamą u s t n ą - c i e ś ń gardzieli, z krtanią - wejście do krtani. Ku dołowi gardło przechodzi w przełyk (esophagus).
Część nosowa gardła Część nosowa gardła (pars nasalis pharyngis) czynnościowo należy do układu oddechowego. Ściany tej części są przymocowane nieruchomo do kości czaszki i dlatego jest ona zawsze otwarta. Na sklepieniu gardła, przy przejściu ściany górnej w tylną, znajduje się skupienie tkanki limfatycznej, 179
tworzące nieparzysty migdałek g a r d ł o w y (tonsilla pharyngea). Migdałek ten u dzieci czasem przerasta, utrudniając przepływ powietrza z jamy nosowej. W bocznej ścianie części nosowej gardła znajduje się z każdej strony owalny lub trójkątny otwór - ujście gardłowe trąbki słuchowej. Otwór ten leży nieco ponad poziomem podniebienia twardego, na wysokości tylnego końca małżow i n y nosowej dolnej. Ujście to ograniczają od tyłu i od góry dwa fałdy, z których większy, tylny, zwany jest w a ł e m t r ą b k o w y m . Bezpośrednio za w a ł e m t r ą b k o w y m znajduje się wąski, pionowo ustawiony, zachyłek gardłow y . Skupienie tkanki adenoidalnej w wale t r ą b k o w y m nosi nazwę migdałka t r ą b k o w e g o (tonsilla tubaria).
Część ustna gardła Część ustna gardła (pars oralis pharyngis) leży bezpośrednio za jamą ustną, z którą łączy ją cieśń gardzieli (isthmus faucium). Jak już w s p o m n i a n o , ściana boczna cieśni gardzieli ma kształt trójkątnego pola, ograniczonego od przodu łukiem podniebienno-językowym, a od tyłu łukiem podniebienno-gardłowym. Pole to nosi nazwę zatoki migdałkowej, gdyż leży tu migdałek podniebienny, o którym w s p o m n i a n o przy omawianiu j a m y ustnej. Ściana tylna jamy ustnej gardła jest widoczna i dostępna badaniu od strony ustnej, przy rozwartych szczękach. Migdałek podniebienny (tonsilla palatina) ma kształt owalny, o osi długiej, ustawionej pionowo. Wielkość migdałka jest zmienna. Długość jego w y n o s i przeciętnie około 20-22 m m , szerokość 18-20 mm i grubość 12-15 m m ; może on już to wystawać spoza łuku podniebienno-językowego do j a m y gardła, już to bywa za t y m łukiem schowany. Powierzchnia przyśrodkowa migdałka pokryta jest nabłonkiem płaskim. Na powierzchni tej znajdują się wąskie, głębokie wręby, w które wchodzi błona śluzowa. Migdałki łatwo ulegają powiększeniu, zwłaszcza u dzieci. Przy stanach zapalnych mogą one znacznie zwężać cieśń gardzieli.
Część krtaniowa gardła Część krtaniowa gardła (pars laryngea pharyngis) położona jest w tyle za krtanią, mając od przodu chrząstkę nagłośniową i wejście do krtani. W ścianie bocznej tej części gardła znajduje się dosyć głęboki zachyłek groszkowaty (recessus piriformis). Część krtaniowa, dosyć szeroka u góry, ku dołowi nagle się zwęża i na poziomie górnego brzegu krtani przechodzi w przełyk.
Budowa ściany gardła Ściana ga rdła składa się z czterech warstw. Licząc od zewnątrz są t o : 1) błona zewnętrzna, 2) błona mięśniowa, 3) błona włóknjsta i 4) błona śluzowa. 180
1. Błona zewnętrzna (tunica adventitia) stanowi przedłużenie powięzi policzkowo-gardłowej. Pokrywa ona od zewnątrz mięśnie gardła oraz łączy je z narządami sąsiednimi. 2. Błona mięśniowa (tunica muscularis) składa się z mięśni poprzecznie prążkowanych, przebiegających w dwóch kierunkach: okrężnym (zwieracze gardła) i podłużnym (dźwigacze gardła). 3. Błona w ł ó k n i s t a gardła (membrana fibrosa), położona pod warstwą mięśniową, jest mocną blaszką łącznotkankową, która u góry przyczepia się do podstawy czaszki. 4. Błona śluzowa gardła (tunica mucosa) pokryta jest w części ustnej i krtaniowej gardła nabłonkiem w i e l o w a r s t w o w y m płaskim. W części nosowej gardła występuje nabłonek walcowaty migawkowy. W części ustnej gardła następuje skrzyżowanie drogi pokarmowej i oddechowej. Przy zaburzeniach w przełykaniu powietrze może trafić do przełyku (aerophagia) i dalej do żołądka, albo też cząstki pokarmów mogą trafić do krtani i tchawicy (zakrztuszenie się, zachłyśnięcie się). Część krtaniowa gardła, położona poniżej wejścia do krtani, należy wyłącznie do układu pokarmowego, część nosowa - wyłącznie do układu oddechowego.
TRAWIENIE POKARMU W JAMIE USTNEJ W jamie ustnej następuje dokładna ocena pokarmów, które się tu dostają. Skład chemiczny pokarmu w y w o ł u j e uczucie smaku. Rozróżniamy cztery podstawowe rodzaje smaku: słodki, gorzki, słony i kwaśny. Czuciem dotyku rozpoznajemy konsystencję i stopień rozdrobnienia pokarmu, które dokonuje się w trakcie żucia. Temperatura pokarmu w y w o ł u j e czucie zimna i ciepła. Duże znaczenie dla oceny przydatności pokarmu ma zmysł węchu. Wszystkie te wrażenia decydują o przyjęciu pokarmu do dalszej przeróbki lub wydaleniu go na zewnątrz. Podczas żucia wydziela się do jamy ustnej duża ilość śliny. Ślina jest produktem czynności gruczołów ślinowych. W skład gruczołów ślinowych wchodzą dwa rodzaje komórek: śluzowe i surowicze. Pierwsze produkują gęstą, ciągnącą się wydzielinę, drugie - ślinę rzadką, wodnistą. Z komórek surowiczych zbudowana jest ślinianka przyuszna i niektóre gruczoły języka. Gruczoły śluzowe występują na nasadzie języka, na podniebieniu miękkim i t w a r d y m . W skład ślinianek podżuchwowej i podjęzykowej wchodzą obydwa rodzaje komórek.- Mieszane gruczoły, głównie śluzowe, występują również w błonie śluzowej policzków, warg i końca języka. Ślina (saliva) jest cieczą wodojasną, w skład której wchodzi duża ilość w o d y (99,5%), pewna ilość białka, mucyny, sole mineralne oraz enzym trawiący węglowodany - amylaza ślinowa, zwana też ptialiną. Ciężar właściwy śliny w y n o s i 1,005. Odczyn jej jest prawie obojętny (pH 6,4-7,0). Ilość wydzielanej na dobę śliny ulega dużym wahaniom, przeciętnie wynosi 1-1,5 I. 181
Zadanie śliny polega na: 1) zmiękczeniu i zwilżeniu pokarmu w czasie żucia i połykania, 2) pokryciu kęsa pokarmu śIuzem w celu zmniejszenia tarcia w czasie jego wędrówki przez gardło i przełyk, 3) rozpuszczeniu suchych składników pokarmowych, które tylko w roztworze mogą pobudzać zakończenia smakowe w jamie ustnej, 4) wstępnym trawieniu skrobi, 5) współdziałaniu w utrzymaniu odpowiedniej ilości wody w ustroju. Jeżeli zawartość wody w ustroju spadnie poniżej pewnej granicy, następuje zahamowanie wydzielania śliny i zmniejszenie wilgotności błony śluzowej jamy ustnej i gardła, co wywołuje uczucie pragnienia. Amylaza ślinowa jest enzymem, który przyśpiesza zamianę skrobi na dekstryny i maltozę. Rozkład skrobi pod wpływem amylazy ślinowej zachodzi najlepiej w temperaturze ciała w odczynie obojętnym. Skrobia gotowana ulega tym przemianom znacznie łatwiej niż surowa. Pokarm przebywa w jamie ustnej zbyt krótko, aby rozkład skrobi mógł zajść dość daleko, zostaje on tu tylko zapoczątkowany. Jeżeli przeżuty i zmieszany ze śliną pokarm złożony ze skrobi potrzymamy przez dłuższy czas w uętach, nabiera on smaku słodkawego.
POŁYKANIE Połykanie jest złożonym aktem odruchowym, w którym odróżniamy trzy fazy związane z trzema początkowymi odcinkami przewodu pokarmowego jamą ustną, gardłem i przełykiem. W fazie pje rwszej po karm przeżuty i wymieszany ze śliną, dzięki ruchom języka i policzków, zostaje uformowany w kęs. Połykanie rozpoczyna się przez przyciśnięcie końca języka do podniebienia, a następnie przesuwanie kęsa ku tyłowi i przepchnięcie go przez cieśń gardzieli do jamy^ardła. W tym czasie kęs zostaje obficie zwilżony wydzieliną śluzową gruczołów leżących w okolicy icieśni gardzieli. Od tej chwili rozpoczyna się druga faza połykania, niezależna od wolj, złożona z serii szybkich i skomplikowanych ruchów, koordynowanych przez ośrodki w rdzeniu przedłużonym. W chwili gdy kęs wchodzi do gardła, następuje odruchowy skurcz mięśni unoszących podniebienie miękkie, które przylega do tylnej ściany gardła i uniemożliwia przedostanie się połykanego pokarmu do jamy nosa. Powrotna droga do jamy ustnej jest zamknięta uniesioną do góry nasadą języka i przylegającymi do niej szczelnie łukami podniebiennymi. Odruchowy skurcz mięśni zwieraczy gardła przesuwa przełykany kęs ku dołowi. Równocześnie krtań zostaje podciągnięta ku górze i przodowi i ukryta pod nasadą języka, przy czym zostaje otwarte wejście do przełyku, do którego kęs zostaje przepchnięty. Wszystkie ruchy wfazie drugiej powstają na drodze odruchowej. Odruchy te są wyzwolone przez podrażnienie zakończeń nerwów czuciowych na bodźce dotykowe. Znieczulenie tych okolic uniemożliwia połykanie. W fazie trzeciej kęs pokarmowy wędruje przez przełyk, przesuwany skurcza182
rii mięśniówki okrężnej. W dolnej części przełyku kęs posuwa się znacznie wolniej niż w 1 / 3 górnej, gdzie są mięśnie poprzecznie prążkowane. Kęs miękki i dobrze zwilżony dochodzi do wpustu w ciągu 5 s, suchy wędruje w o l n i e j (ok. 4 cm/s); powstają przy t y m wtórne fazy skurczów, które przepychają kęs do żołądka, odczuwane jako nieprzyjemne drapanie wzdłuż przełyku. Płyny, dzięki pchnięciu nadanemu przy połykaniu i sile ciężkości, wyprzedzają falę skurczu i dochodzą do żołądka w ciągu sekundy. Wpust otwiera się w chwili, gdy dochodzi doń fala perystaltyczna przełyku.
PRZEŁYK Przełyk (esophagus) '\est najwęższą częścią przewodu pokarmowego. Ma on kształt długiego wąskiego cylindra, ciągnącego się od dolnej granicy gardła, tj. od poziomu VI kręgu szyjnego, aż do wpustu żołądka położonego na wysokości X - X I kręgu piersiowego. Długość przełyku wynosi 25-30 cm, średnica przełyku pustego około 13 m m , w stanie rozciągniętym może osiągać ponad 25 m m . Odpowiednio do okolicy, w jakiej przebiega, rozróżniamy w przełyku: część szyjną, piersiową i brzuszną. Krótka część szyjna przełyku leży bezpośrednio za tchawicą, iuźno łącząc się z jej ścianą tylną. Najdłuższa - część piersiowa, biegnie w śródpiersiu t y l n y m w kierunku ku dołowi. Najkrótsza - część brzuszna, długości 2 - 5 cm, rozpoczyna się po przejściu przez otwór przełykowy przepony i kończy się w p u s t e m żołądka. Przełyk na sWym przebiegu wykazuje trzy zwężenia: pierwsze znajduje się na granicy z gardłem, drugie - w miejscu skrzyżowania z l e w y m oskrzelem g ł ó w n y m , trzecie - w obrębie przepony. W zwężeniach tych mogą się zatrzymywać przełknięte ciała obce. Ściana przełyku składa się z trzech warstw: błony śluzowej, mięśniowej i zewnętrznej.
ŻOŁĄDEK Żołądek (ventriculus, gaster)j>tanowi workowate rozszerzenie przewodu pokarmowego, które ma do spełnienia dwa zadania: 1) stanowi zbiornik pokarmu, 2) jest narządem wydzielniczym wytwarzającym kwas solny i enzymy. Pod w p ł y w e m tych enzymów pokarm zostaje zmieniony na płynną miazgę (chymus). Żołądek leży ukośnie od góry i strony lewej ku dołowi i stronie prawej w okolicy nadbrzusznej i podżebrzu lewym. Kształt i położenie żołądka u człowieka żywego ustawicznie się zmieniają i zależą od jego wypełnienia, okresu'trawienia, stanu mięśniówki, jak również od stanu narządów sąsiednich. Liekko wypełniony żołądek ma kształt haka. W początkowym okresie trawienia żołądek wygląda jak złożony z dwóch części: lewej - rozszerzonej 183
balonowato, i prawej - wąskiej, cylindrycznej. Przy skurczonej mięśniówce żołądek przybiera kształt pończochy. Gdy mięśniówka traci całkowicie swe napięcie, co u człowieka żywego zdarza się niezwykle rzadko, żołądek przybiera kształt retorty lub gruszki, taki jaki znajdujemy na zwłokach i jaki zwykle bywa opisywany w podręcznikach anatomii. W żołądku rozróżniamy dwie ściany: przednią, zwróconą ku przodowi, dołowi i ku stronie lewej, oraz ścianę tylną, skierowaną ku tyłowi, górze i ku stronie prawej. Ściany te łączą się ze sobą u góry wzdłuż tzw. krzywizny
Pęcherzyk żółciowy
Więzadło obłe wątroby
Dno
^żołądka
Przewód wątrobowy
-Przepona
Wątroba - — płat prawy
_ - Aorta Żyła wrotna
Przęwód pęcherzykowy
• Żołądek Przewód żółciowy wspólny
Dwunastnica
Trzustkd
Ryc. 109. Wątroba, żołądek, dwunastnica i trzustka. Schemat ilustrujący wzajemne stosunki tych narządów.
mniejszej, u dołu - wzdłuż krzywizny większej. W g ó r n y m końcu krzywizny mniejszej żołądek łączy się z przełykiem. Otwór stanowiący ujście przełyku do żołądka nazywa się w p u s t e m (cardia). Ujście żołądka do dwunastnicy nazywa się odźwiernikiem (pylorus). Odźw.iernik zaopatrzony jest w gruby pierścień mięśniówki okrężnej, tworzący mięsień zwieracz odźwiernika, którego skurcz oddziela światło żołądka od dalszego odcinka przewodu pokarmowego. Zaokrąglona, obszerna część żołądka położona na lewo i ku górze od wpustu, nazywa się d n e m żołądka. Dno umieszczone jest pod lewą kopułą przepony. Część żołądka sąsiadująca z odźwiernikiem nosi nazwę części 184
odźwiernikowej. Pomiędzy częścią odźwiernikową a dnem znajduje się trzon żołądka. Część odźwiernikową u człowieka żywego bardzo często oddziela od trzonu lekkie przewężenie. o
Budowa ściany żołądka ©
Ściana żołądka składa się z czterech warstw: 1) błony surowiczej, 2) błony mięśniowej, 3) tkanki podśluzowej i 4) błony śluzowej. 1. Błona surowicza jest częścią otrzewnej, która pokrywa cały żołądek z wyjątkiem wąskiego pasa wzdłuż obu jego krzywizn. 2. Błona mięśniowa, zbudowana z pęczków mięśni gładkich, składa się z trzech warstw. Warstwa zewnętrzna zawiera pęczki mięśniowe ułożone podłużnie, najgęściej skupione na krzywiźnie mniejszej i większej. Środkowa warstwa składa się z pęczków o przebiegu okrężnym. W ścianie odźwiernika mięśniówka okrężna grubieje bardzo znacznie, tworząc tam mięsień zwieracz odźwiernika. Niekompletną warstwę wewnętrzną stanowią włókna skośne, które wychodzą z warstwy okrężnej mięśniówki przełyku i rozchodzą się wachlarzowato na obie ściany żołądka. 3. Tkanka podśluzowa jest dość grubą warstwą tkanki łącznej wiotkiej, zawierającej liczne naczynia krwionośne i nerwy. 4. Błona śluzowa stanowi wewnętrzną warstwę żołądka, skierowaną ku jego światłu. Jest ona miękka, dosyć gruba, barwy szaroróżowej, najgrubsza w okolicy odźwiernikowej, najcieńsza w obrębie dna. W pustym żołądku błona śluzowa jest silnie pofałdowana. Fałdy te krzyżują się ze sobą w różnych kierunkach i jedynie w obrębie krzywizny mniejszej występują trzy lub cztery stałe fałdy podłużne, ciągnące się od wpustu do odźwiernika. Powierzchnię błony śluzowej żołądka pokrywa nabłonek jednowarstwowy walcowaty. Na powierzchni tej przy niewielkim powiększeniu można dostrzec liczne punkcikowate ujścia gruczołów żołądkowych, tzw. dołeczki żołądkowe. W żołądku rozróżniamy dwa rodzaje gruczołów: gmczoły żołądkowe właściwe [gruczoły odźwiernikowe. Gruczoły żołądkowe właściwe występują w wielkiej liczbie (ok. 100 na 1 mm 2 ) w okolicy dna i trzonu żołądka. Są to gruczoły cewkowe proste, zawierające dwa rodzaje komórek nabłonkowych: kornórki główne, wyściełające światło gruczołu, oraz komórki okładzinowe. Pierwsze wytwarzają pepsynogen, drugie - kwas solny. Pepsynogen w obecności kwasu solnego zmienia się w pepsynę, która jest enzymem proteolitycznym żołądka. Gruczoły odźwiernikowe występują w części odźwiernikowej żołądka. Są to proste lub złożone gruczoły cewkowo-pęcherzykowe, zbudowane z komórek nabłonka walcowatego. Wydzielają one prawi e wyłącznie śluz. Śluz ten tworzy na powierzchni błony śluzowej warstwę grubości ok. 1 mm, chroniąc ją przed strawieniem przez enzymy żołądkowe oraz zmniejszając tarcie przy przechodzeniu pokarmu. 185
Trawienie pokarmu w żołądku Pusty żołądek ma światło niewiele większe od jelita cienkiego. Jego pojemność wynosi ok. 50 ml, a błona śluzowa jest silnie pofałdowana. W miarę nadchodzenia pokarmu ściany żołądka rozciągają się. Dzięki napięciu mięśniówki pojemność jest zawsze dopasowana do jego zawartości. Przybywający pokarm układa się w ten sposób, że pierwsze jego porcje zajmują miejsce przy ścianach i w części odźwiemikowej, następne układają się "koncentrycznymi warstwami w środku. W ciągu ok. pół godziny po spożyciu pokarmu aktywność ruchowa żołądka pozostaje bardzo słaba, następnie czynność jego stopniowo się wzmaga, najpierw w postaci skurczów słabych, a później coraz mocniejszych, które rozpoczynają się w środkowej jego części i przesuwają w kierunku odźwiernika. Dzięki tym ruchom pokarm zostaje wymieszany z sokiem żołądkowym
r
Ryc. 110. Schemat ruchów żołądka.
86
i zamieniony na płynną miazgę pokarmową (chymus). W miarę wzrostu kwasoty treści żołądkowej skurcze żołądka stają się coraz silniejsze. Gdy znajdujący się w żołądku pokarm uzyska odpowiednią konsystencję, zstępująca fala perystaltyczna przepycha go do dwunastnicy. Odźwiernik pozostaje przez większy okres czasu otwarty i kurczy się w c h w i l i , gdy przechodzi przezeń fala perystaltyczna. Część żołądka sąsiadująca z dnem nie wykazuje tak silnej aktywności ruchowej. Zgromadzony tu pokarm pozostaje przez dłuższy czas we względnym spokoju, nie zmieszany z sokiem żołądkowym, co pozwala na dalsze działanie amylazy ślinowej. W miarę przechodzenia miazgi pokarmowej do dwunastnicy pokarm z okolicy dna żołądka przesuwa się do części odźwiernikowej. Czas trawienia w żołądku jest różny i zależy od właściwości spożytego pokarmu. Pokarmy płynne słabo zakwaszone przechodzą przez żołądek bez zatrzymania. Niezbyt obfite posiłki przebywają w żołądku 1 - 2 h, w i ę k s z e - 5 - 7 h.
Sok żołądkowy M e t o d y otrzymywania soku żołądkowego W żołądku pokarm ulega dosyć g r u n t o w n y m przemianom chemicznym p o d w p ł y w e m soku żołądkowego. Sok żołądkowyJest produktem czynności wydzielniczej gruczołów żołądkowych. U człowieka otrzymano po raz pierwszy sok żołądkowy od chorego z przewlekłą przetoką żołądkową. W praktyce klinicznej do celów diagnostycznych otrzymuje się sok żołądkowy przez cienki zgłębnik długości 50 cm, który zostaje przełknięty przez pacjenta i wprowadzony do żołądka. Uprzednio żołądek przepłukuje się wodą, a następnie stosuje się jakiś bodziec wydzielniczy, np. wstrzyknięcie histaminy, insuliny lub pentagastryny, który powoduje obfite wydzielanie soku żołądkowego.
Skład soku żołądkowego Sok żołądkowy jest płynem bezbarwnym, wodojasnym, o charakterystycznym zapachu i odczynie silnie kwaśnym (pH - 1 ) . Ilość wydzielanego na dobę soku żołądkowego zależna jest od ilości i rodzaju spożywanego pokarmu i waha się w granicach 1,5-2,5 I. W skład soku żołądkowego_wchodzi: kwas solny 0,4-0,5%, sole mineralne, głównie NaCI i KCI, w ilości 0,3-0,5%, oraz niewielka ilość białka. Sok żołądkowy zawiera również enzymy trawienne: pepsynę, chymozynę ijipazę. Pepsyna jest enzymem proteolitycznym (proteazą) wydzielanym w postaci nieczynnej, jako tzw. pepsynogen, przez komórki główne gruczołów żołądkowych. Pepsynogen pod w p ł y w e m kwasu solnego zamienia się w pepsynę. 187
Pepsynajest enzymem silnie działającym w środowisku kwaśnym. Powoduje ona rozpadłiydrolityczny białek przez kilka etapów przejściowych doalbumoz i peptonów. W ten sposób białka zostają przygotowane do całkowjtego rozbicia przez enzymy trzustki - trypsynę i erepsynę, które rozkładają peptony najiminokwasy. Podpuszczka (chymozyna) jest wydzielana w postaci enzymu również przez komorki główne gruczołów żołądkowych. Rozkłada ona tylko rozpuszczalne białko mleka - kazeinę, którą zamienia na parakazeinę, ta zaś w obecności wapnia wytrąca się tworząc sernik, dalej trawiony przez pepsynę i trypsynę. Najwięcej podpuszczki zawierają żołądki noworodków karmionych mlekiem matki. Lipaza żołądkowa występuje w soku żołądkowym w niewielkiej ilości. Działa ona tylko na te tłuszcze, które nadchodzą do żołądka w postaci zemulgowanej, jak np. tłuszcze mleka, śmietany, jaj itp. Kwas solny: 1) aktywuje pepsyno gen, zamieniając go w pepsynę, 2) wytwarza środowisko kwaśne, które jest nieodzowne dla działania pepsyny, 3) powoduje pęcznienie włókien białkowych, które w-tym stanie łatwiej ulegają działaniu pepsyny, 4) działa jako środek antyseptyczny, zabijając liczne drobnoustroje, które dostają się do żołądka z pokarmem, 5) bierze udziałw regulacji otwierania i zamykania odźwiernika.
Regulacja wydzielania soku żołądkowego Wydzielanie soku żołądkowego jest procesem, który rozpoczyna się jeszcze zanim pokarm znajdzie się w żołądku, pod wpływem takich bodźców, jak: widok, zapach lub nawet sama myśl o pokarmie. Bodźce te powstają w korze mózgowej i są przekazywane do żołądka przez włókna nerwu błędnego. Jest to tzw. faza nerwowa wydzielania żołądkowego. Utrzymuje się ona 10-20 min od chwili zadziałania bodźca i przyczynia się do ok. V 3 całej sekrecji żołądkowej związanej ze spożyciem pokarmu. Natomiast podrażnienie układu współczulnego hamuje czynność gruczołów trawiennych. Po fazie nerwowej następuje faza żołądkowa wydzielania, która rozpoczyna się z chwilą gdy spożyty pokarm znajdzie się w żołądku. Rozciąganie żołądka przez pokarm jest bodźcem do wydzielania przez błonę śluzową części odźwiernikowej swoistego hormonu zwanego gastryną. Gastryna dochodząc do gruczołów żołądkowych pobudza je do wydzielania soków trawiennych, szczególnie - kwasu solnego. W fazie żołądkowej wydzielanie nie jest tak obfite, trwa jednak znacznie dłużej - do kilku godzin. Pó fazie żołądkowej następuje trzecia - faza jelitowa wydzielania żołądkowego. Udział jej w ogólnym wydzielaniu żołądkowym jest niewielki (ok. 5%).
188
WYMIOTY W warunkach prawidłowych pokarm nie wraca do przełyku, jednakże silne skurcze spastyczne mięśni brzucha, przepony i ścian żołądka mogą wyrzucić zawartość żołądka przez przełyk i jamę ustną na zewnątrz, co nazywamy w y m i o t a m i . W y m i o t y są odruchem obronnym, dzięki któremu ustrój pozbywa się szkodliwych substancji dostających się do żołądka. W y m i o t y poprzedzane są zwykle uczuciem nudności i o d r u c h o w y m wydzielaniem śliny. W y m i o t y mogą być w y w o ł a n e przez podrażnienie odpowiednich n e r w ó w czuciowych, np. przez drażnienie podstawy języka, ściany gardła, otrzewnej itp., albo automatycznie przez podrażnienie odpowiednich ośrodków w wyniku wprowadzenia do krwi pewnych trucizn, jak np. apomorfiny.
JELITO Jelitem (intestinum) nazywamy najdłuższą część przewodu pokarmowego, rozpoczynającą się od żołądka i kończącą odbytem. Jelito dzieli się neTdwle odmienne części: jelito cienkiej jelito grube.
Jelito cienkie Budowa jelita cienkiego Jelito cienkie (intestinum tenue) rozpoczyna się od odźwiernika żołądka i kończy ujściem do jelita grubego, położonym w p r a w y m dole b i o d r o w y m . Długość jelita cienkiego po wyjęciu ze zwłok wynosi 4—5 m, średnica - 45-25 m m ; zażycia jest ono krótsze. Odległość od odźwiernika do zastawki krętriiczo-kątniczej mierzona zgłębnikiem u człówieka żywego w y n o s i ok. 3 m. Rozróżniamy w n i m trzy odcinki: 1) dwunastnicę (duodenum), 2) jelito czcze (jejunum) i 3) jelito kręte ( i l e u m ) . Dwunastnica stanowi początkowy odcinek jelita cienkiego pozbawiony krezki. Jelito czcze przechodzi bez wyraźnej granicy w jelito kręte. Ściana jelita cienkiego składa się z czterech w a r s t w : 1 ) błony surowiczej, 2) błony mięśniowej, 3) tkanki podśluzowej i 4) błony śluzowej. 1. Błona surowicza jest częścią otrzewnej, która ze ścian jamy brzusznej przechodzi na jelito" za pośrednictwem krezki grzbietowej. Błona ta stanowi warstwę zewnętrzną, która pokrywa jelito czcze i kręte w całości. Natomiast dwunastnica ma pokrywę surowiczą tylko od przodu. 2. Błona mięśniowa utworzona jest z pęczków mięśni gładkich, ułożonych w dwóch warstwach. Warstwę zewnętrzną tworzy mięśniówka podłużna, warstwę wewnętrzną - okrężna. Warstwa okrężna jest 2 - 3 razy grubsza od podłużnej. 189
Fałdy jelita cienkiego
%J>, Wycinek -powiększony, na ryc. B
A Wycinek powiększony na ryc. A,
Kosmek jelitowy
Fałd jelita pokryty kosmkami
Wycinak powierzchni kosmka w powiększeniu
Ryc. 111. Budowa ściany jelita cienkiego: , 4 - f a ł d jelitowy,/A;-powiększony wycinek, B - k o s m e k jelitowy, C- mikrokosmki pokrywające powierzchnię nabłonka jelitowego.
3. Tkanka podśluzowa, łącząca błonę śluzową z błoną mięśniową, przedstawia się w postaci dobrze rozwiniętej w a r s t w y tkanki łącznej wiotkiej, dzięki której położona nad nią błona śluzowa ma dużą ruchomość. 4. Błona śluzowa jest gruba i obficie unaczyniona. Pokrywa ją nabłonek jednowarstwowy walcowaty, który tworzy liczne gruczoły jelitowe. Błona śluzowa jelita cienkiego tworzy liczne fałdy okrężne. W odróżnieniu od fałdów 190
śluzówki żołądka fałdy okrężne jelita nie znikają przy rozciąganiu jelit. Fałdy okrężne rozpoczynają się w odległości 2,5 - 5 cm za odźwiernikiem i są najlepiej rozwinięte w dwunastnicy i początkowej części jelita czczego; w dalszym odcinku jelita stają się coraz mniejsze, nieregularne i w końcu jelita cienkiego zanikają prawie zupełnie. Pofałdowanie zwiększa powierzchnię jelita, przez którą odbywa się wydzielanie soków trawiennych i wchłanianie pokarmu. Powierzchnię błony śluzowej całego jelita cienkiego pokrywają poza t y m liczne drobne kosmki jelitowe, które nadają jej wygląd aksamitu. Liczba kosmków w jelicie człowieka sięga kilku milionów. Długość kosmka jelitowego wynosi 0,5-1,5 m m , grubość ich jest znikoma. W dwunastnicy kosmki mogą być rozgałęzione, w pozostałych jelitach mają kształt nitkowaty. Kosmki są narządami, za pomocą których odbywa się wchłanianie strawionego pokarmu. Pod powierzchnią każdego kosmka znajduje się sieć naczyń włosowatych krwionośnych, do których przenikają z jelita produkty trawienia w ę g l o w o d a n ó w i białek. Zawiesina drobnych kropelek tłuszczu natomiast zostaje wchłonięta do naczynia chłonnego, położonego centralnie w osi kosmka, skąd zostaje doprowadzona do układu naczyń chłonnych (limfatycznych), nadając mlecznobiałe zabarwienie limfie (chłonce) odpływającej z jelit. W ścisłym związku z naczyniami limfatycznymi, przebiegającymi w ścianie jelita cienkiego, pozostają skupienia tkanki limfatycznej w kształcie okrągłych t w o r ó w wielkości główki szpilki, zwane grudkami limfatycznymi samotnymi. Większe skupienia grudek limfatycznych noszą nazwę grudek skupionych.
Gruczoły jelita cienkiego W błonie śluzowej jelita cienkiego występują liczne gruczoły jelitowe. Gruczoły jelitowe są to proste cewkowate zagłębienia w błonie śluzowej, wysłane nabłonkiem walcowatym. Ujścia tych gruczołów mają postać okrągłych o t w o r k ó w położonych pomiędzy kosmkami jelitowymi. Gruczoły te występują na całej długości jeljta cienkjego. Gruczoły dwunastnicze są to złożone gruczoły cewkowo-pęcherzykowe, występujące w błonie podśluzowej dwunastnicy. Gruczoły jelitowe i dwunastnicze wydzielają sok jelitowy (succus entericus) przeznaczony do trawienia pokarmu.
Dwunastnica Dwunastnica (duodenum) jest początkowym odcinkiem jelita cienkiego, długości ok. 25 cm, średnicy 3,5-5 cm, który łączy się z odźwiernikiem żołądka. Nie ma ona krezki i jest przytwierdzona do tylnej ściany jamy brzusznej, leżąc pozaotrzewnowo. Uchodzą do niej przewody wyprowadzające dwóch najwi kszych gruczołów trawiennych - wątroby i trzustki. 191
Dwunastnica ma kształt podkowy otwartej ku stronie lewej i ku dołowi. Rozróżniamy w niej cztery części: 1) część górną, ciągnącą się od odźwiernika (na wysokości I kręgu lędźwiowego) ukośnie ku tyłowi, nieznacznie na prawo i lekko ku górze; 2) część zstępującą, biegnącą prawie pionowo w dół do dolnego brzegu III kręgu lędźwiowego; 3) część poziomą, krzyżującą poziomo kręgosłup i 4) część wstępującą, biegnącą ukośnie ku górze po lewej stronie aorty brzusznej do poziomu II kręgu lędźwiowego, gdzie zgina się ku przodowi i dołowi, by przejść w jelito czcze. W początkowym odcinku dwunastnicy, zwanym przez radiologów opuszką dwunastnicy, błona śluzowa nie ma fałdów. Mniej więcej w połowie części zstępującej błona śluzowa tworzy fałd podłużny dwunastnicy, kończący się wzniesieniem, zwanym brodawką większą dwunastnicy. Na brodawce tej znajduje się ujście przewodu żółciowego wspólnego i przewód u trzustkowego. W pobliżu tej brodawki znajduje się czasem brodawka mniejsza, na której wierzchołku otwiera się przewód trzustkowy dodatkowy. Błona śluzowa dwunastnicy.jest grubsza niż w pozostałej części jelita cienkiego. Pokrywają ją krótkie, dość grube kosmki. Błona podśluzowa wyróżnia się tym, że zawiera, głównie w górnej połowie dwunastnicy, gruczoły dwunastnicze.
Jelito cienkie krezkowe (jelito czcze i jelito kręte) Jelito czcze i jelito kręte są przymocowane do tylnej ściany jjamy brzusznej za pomocą krezki, są przeto także nazywane jelitem cienkim krezkowym. Krezka jelita cienkiego jest podwójnym fałdem otrzewnej, który schodzi z tylnej ściany jamy brzusznej i w swym w o l n y m brzegu zawiera jelito. Brzeg wolny krezki ma długość równą długości jelita, natomiast brzeg przeciwległy, przytwierdzony do ściany jamy brzusznej, zwany korzeniem krezki, jest krótki (długość jego wynosi tylko ok. 15 cm) i rozciąga się skośnie od lewej powierzchni II kręgu lędźwiowego do okolicy prawego stawu biodrowo-krzyżowego. Tak wielka różnica długości obu brzegów krezki może być wyrównana tylko przez silne pofałdowanie brzegu wolnego, co czyni ją podobną do krezy i stąd nazwa. Wobec tego jelito cienkie układa się w liczne pętle, które wypełniają większą część jamy brzusznej i sięgają także do miednicy małej. Pętle te mogą przesuwać się i zajmować różne położenia w jamie brzusznej; dlatego też położenie poszczególnych części jelita cienkiego jest bardzo zmienne. Ogólnie można powiedzieć, że jelito czcze zajmuje lewą część jamy brzusznej poniżej żołądka, zaś pętle jelita krętego układają się w prawej i dolnej części jamy brzusznej. Pomiędzy dwiema blaszkami krezki przebiegają naczynia krwionośne i limfatyczne oraz nerwy zdążające do jelita. W krezce znajdują się poza tym liczne węzły chłonne oraz pewna ilość tkanki tłuszczowej. 192
Jelito czcze przechodzi w jelito kręte bez wyraźnej granicy. Mniej więcej 2 / 5 górne części jelita krezkowego stanowi jelito czcze, 3 / 5 dolne - jelito kręte. Różnice pomiędzy jelitem czczym i krętym podaje tab. 6. Tabela 6 Jelito czcze
Jelito kręte
Szersze - średnica 31-39 mm Ściana jest grubsza, barwy ciemniejszej, obficiej unaczyniona Fałdy okrężne dobrze rozwinięte Kosmki krótkie i szerokie Grudki limfatyczne samotne mniej liczne, małe Grudek limfatycznych skupionych brak Mniej tkanki tłuszczowej w krezce Przezroczyste pólka w krezce w pobliżu jelita
węższe - średnica 25-31 mm ściana cieńsza i jaśniejsza, bledsza i słabiej unaczyniona fałdy okrężne niskie, krótsze, mniej liczne kosmki cienkie, nitkowate grudki limfatyczne samotne duże i liczne grudki limfatyczne skupione występują skupienie tkanki tłuszczowej w krezce krezka nieprzezroczysta
Jelito grube Jelito grube (intestinum crassum) rozpoczyna się w p r a w y m dole biodrow y m szerokim w o r k o w a t y m odcinkiem ślepo zakończonym, zwanym jelitem ślepym albo kątnicą. Jelito cienkie łączy się z jelitem g r u b y m nie w jego odcinku początkowym, lecz ok. 7 cm powyżej końca jelita ślepego. W pozostał y m odcinku jelita grubego rozróżniamy następujące części: okrężnicę, dzielącą się na: a) wstępującą, b) poprzeczną, c) zstępującą i d) esowatą oraz odbytnicę. Długość jelita grubego wynosi 130 - 150 cm. Najszerszą częścią jelita grubego jest kątnica (ok. 75 mm średnicy). W miarę posuwania się ku odbytowi średnica jelita nieco się zmniejsza. W jelicie g r u b y m , w odróżnieniu do jelita cienkiego, warstwa mięśniówki podłużnej nie jest rozłożona w ścianie jelita równomiernie, lecz skupiona w trzech taśmach szerokości ok. 6 m m , przebiegających podłużnie w jednakowej od siebie odległości. Jedna z nich, tzw. taśma swobodna, ciągnie się na brzegu w o l n y m jelita, taśma sieciowa - wzdłuż przyczepu sieci większej do okrężnicy poprzecznej i trzecia, taśma krezkowa - wzdłuż krawędzi przyczepu jelita. Taśmy te są o 1 / 6 krótsze niż jelito, wskutek czego powstają między nimi trzy rzędy wybrzuszeń, zwanych wypukleniami okrężnicy, porozdzielanych poprzecznymi rowkami, które wpuklają się do światła jelita jako fałdy półksiężycowate. Poza t y m jelito grube odznacza się t y m , że z jego powierzchni zwisają małe kieszonki wypełnione tkanką tłuszczową, tzw. przyczepki sieciowe.
13 - Anatomia i fizjologia
193
Jelito ślepe i w y r o s t e k robaczkowy Jelito ślepe (cecum) stanowi początkowy odcinek jelita grubego, położony poniżej ujścia jelita krętego. Ma ono kształt szerokiego, niesymetrycznego worka, długości około 7 cm, średnicy - ok. 9 cm. Jelito ślepe leży w p r a w y m dole b i o d r o w y m , pokrywając częściowo mięsień lędźwiowo-udowy większy; jest ono pokryte ze wszystkich stron przez otrzewną. Górny jego koniec przechodzi w okrężnicę wstępującą. Vi f i
krętniczo-katnicza
Okrężnica wstępująca
ślepe
Ryc. 112. Jelito ślepe z uwidocznioną zastawką krętniczo-kątniczą i wyrostkiem robaczkowym.
Na granicy z okrężnicą wstępującą do jelita grubego uchodzi jelito cienkie ograniczone zastawką krętniczo-kątniczą. Utworzona przez błonę śluzową i warstwę okrężną mięśniówki końcowego odcinka jelita krętego zastawka ta nie pozwala na cofanie się zawartości jelita grubego do jelita cienkiego. Około 2,5-3,5 cm poniżej zastawki w ścianie tylno-przyśrodkowej jelita ślepego znajduje się ujście wyrostka robaczkowego, zaopatrzone w fałd śluzówki, zwany zastawką tego wyrostka. Wyrostek robaczkowy ( a p p e n d i x v e r m i f o r m i s ) ma kształt cienkiego uchyłka kątnicy o zmiennej długości i średnicy (przeciętnie 7 - 9 cm długości i 6 mm średnicy). Wyrostek robaczkowy jest w całości pokryty otrzewną i ma dosyć dużą krezeczkę, która sięga zwykle do jego końca i łączy go z końcową częścią krezki jelita krętego. Wyrostek robaczkowy nie bierze bezpośredniego udziału w trawieniu. Światło jego ulega często zarośnięciu.
194
Okrężnica Okrężnica wstępująca (colon ascendens) rozpoczyna się powyżej ujścia jelita krętego, skąd biegnie ku górze i nieznacznie ku tyłowi. Po dojściu do trzewnej (dolnej) powierzchni wątroby tworzy zgięcie prawe okrężnicy i przechodzi w okrężnicę poprzeczną, skierowaną przyśrodkowo ku dołowi. Długość okrężnicy wstępującej wynosi 12,5-20 cm; jest ona zwykle szersza niż okrężnica zstępująca. Okrężnica wstępująca jest w większości przypadków pokryta otrzewną z trzech stron, tj. od przodu i z boków. Niekiedy jednak powłoka surowicza otacza ją ze wszystkich stron, tworząc krezkę. Okrężnica poprzeczna (colon transversum) rozpoczyna się w podżebrzu prawym i biegnie ku stronie lewej i ku górze, gdzie zmienia kierunek tworząc zgięcie lewe okrężnicy, by przejść w okrężhicę zstępującą. Długość jej w y n o s i 47-50 cm, a więc więcej niż linia prosta łącząca dwa jej końce, ponieważ okrężnica poprzeczna biegnie łukiem, którego wypukłość, skierowana ku dołowi, zwisa zwykle do wysokości pępka. Okrężnica poprzeczna jest otoczona ze wszystkich stron otrzewną, która tworzy na jej ścianie tylnej podwójny fałd, zwany krezką okrężnicy poprzecznej. Krezka ta przyczepia się do tylnej ściany jamy brzusznej na wysokości górnego brzegu trzustki. Krezka okrężnicy poprzecznej jest krótka w początkowej i końcowej części okrężnicy poprzecznej, natomiast w części środkowej jest znacznie dłuższa, dzięki czemu ta część okrężnicy może znacznie się oddalać od tylnej ściany jamy brzusznej i zmieniać swe położenie w dosyć szerokich granicach. Od przodu okrężnicę poprzeczną pokrywa sieć większa (omentum majus). Ponad nią znajduje się wątroba, pęcherzyk żółciowy, krzywizna większa żołądka i część trzewna (dolna) powierzchni śledziony. W tyle za okrężnicą poprzeczną znajduje się część zstępująca dwunastnicy, głowa trzustki i krezka jelita cienkiego; poniżej - pętle jelita cienkiego. W lewym podżebrzu okrężnica poprzeczna, jak wspomniano, przechodzi w okrężnicę zstępującą. Zgięcie lewe leży wyżej i głębiej niż prawe i styka się ze śledzioną za częścią wpustową żołądka. W położeniu t y m zgięcie okrężnicy utrzymywane jest przez fałd otrzewnej, zwany więzadłem przeponowo-okrężniczym. Okrężnica zstępująca ( colon descendens) rozpoczyna się od zgięcia lewego okrężnicy, skąd biegnie ku dołowi wzdłuż lewej ściany j a m y brzusznej i na wysokości grzebienia miednicy przechodzi w okrężnicę esowatą. Długość jej wynosi 22-30 cm, średnica jest nieco mniejsza niż okrężnicy wstępującej. Okrężnica zstępująca jest w większości przypadków pokryta otrzewną z trzech stron - od przodu i z boków. Okrężnica esowata ( colon sigmoideum) rozciąga się od grzebienia biodrowego do poziomu II i III kręgu krzyżowego. Długość jej jest zmienna i waha się w dużych granicach: od 1 2 d o 8 4 cm (przeciętnie 40 cm). Pokryta jest w całości otrzewną i ma dobrze rozwiniętą krezkę, w związku z czym tworzy dużą pętlę, której położenie bywa różne, zależnie od długości krezki. Najczęściej pętla ta zwisa do miednicy małej. 195
Odbytnica Odbytnica ( rectum) jest końcowym odcinkiem jelita grubego. Rozpoczyna się na wysokości III kręgu krzyżowego i kończy ok. 3,5 cm poniżej wierzchołka kości guzicznej. Długość jej wynosi 15-20 cm. Średnicę ma mniejszą niż okrężnica esowata. Jest ona najmniejsza przy połączeniu z okrężnicą, największa - w części niższej, gdzie występuje wyraźne rozszerzenie, zwane bańką odbytnicy. Średnica pustej odbytnicy wynosi 2,5 cm, wypełnionej - może dochodzić do 7,5 cm. Taśmy okrężnicy na odbytnicy łączą się ze sobą i tworzą jednolitą warstwę mięśniówki podłużnej. Odbytnica na s w y m przebiegu tworzy dwa zgięcia w płaszczyźnie strzałkowej i kilka zgięć w płaszczyźnie czołowej. Pierwsze zgięcie strzałkowe - zgięcie krzyżowe - odpowiada wygięciu kości krzyżowej, drugie - zgięcie kroczowe, skierowane wypukłością ku przodowi, powstaje wskutek zagięcia się końcowego odcinka odbytnicy ku tyłowi w okolicy wierzchołka kości guzicznej. W płaszczyźnie czołowej występują najczęściej również dwa zgięcia, które zaznaczają się na zewnętrznej powierzchni wcięciami, tworzącymi po stronie wewnętrznej fałdy poprzeczne. Otrzewna pokrywa tylko niewielką część odbytnicy, mianowicie jej ściany boczne i przednią do połowy ich długości. Ściana tylna jest przymocowana za pomocą tkanki łącznej wiotkiej i tłuszczowej do ściany i narządów miednicy. Błona mięśniowa odbytnicy składa się z dwóch warstw: zewnętrznej podłużnej, i wewnętrznej - okrężnej. Warstwa podłużna jest znacznie grubsza niż taśmy okrężnicy. Mięśniówka okrężna grubieje w kierunku ku dołowi, by u ujścia odbytnicy utworzyć gruby i wysoki (1-2 cm) pierścień - mięsień zwieracz w e w n ę t r z n y odbytu. Błona śluzowa odbytnicy nie tworzy regularnych fałdów okrężnych. Wyraźnie występują jedynie wspomniane wyżej fałdy poprzeczne, odpowiadające zgięciom odbytnicy. W końcowym odcinku odbytnicy, zwanym częścią kroczową, błona śluzowa tworzy 8 - 1 2 pionowych podłużnych s ł u p ó w odbytniczych, pooddzielanych od siebie zatokami. W słupach tych znajdują się sploty żylne. Otwór końcowy przewodu pokarmowego nazywamy o d b y t e m ( anus).
WIELKIE GRUCZOŁY TRAWIENNE
Wątroba Wątroba (hepar) jest największym gruczołem ciała ludzkiego. Przeciętny jej ciężar wynosi ok. ciężaru całego ciała, tj. ok. 1,5 kg. 0 n o w o r o d k ó w i dzieci ciężar względny wątroby jest jeśzcze większy. Wątroba jest narządem bardzo obficie ukrwionym, ma więc barwę czerwonobrunatną. Zaraz po wyjęciu ze zwłok traci swój kształt i ulega spłaszczeniu. Konsystencja wątroby jest miękka, sprężystość nieznaczna. Miąższ wątroby jest bardzo kruchy, toteż przy upadkach lub urazach wątroba łatwo może pękać. 196
Wątroba zajmuje prawie całe podżebrze prawe, znaczną część nadbrzusza i sięga do podżebrza lewego. Dolna granica wątroby pokrywa się z łukiem żebrowym p r a w y m aż do linii sutkowej prawej i w warunkach prawidłowych spod tego łuku nie wychodzi. W linii środkowej ciała dolny brzeg wątroby znajduje się w połowie odległości pomiędzy podstawą wyrostka mieczykowatego a pępkiem.
Budowa w ą t r o b y Wątroba ma kształt zbliżony do połowy jaja. Powierzchnia wypukła, zwana powierzchnią przeponową, jest zwrócona do przepony, zaś bardziej płaska ku dołowi. Na powierzchni dolnej, trzewnej. Widnieją trzy bruzdy ułożone w kształcie litery H. Bruzda poprzeczna, długości ok. 5 cm, nosi nazwę w r ó t w ą t r o b y (porta hepatis). Przez wrota wchodzą do wątroby: tętnica wątrobowa, żyła wrotna i nerwy, wychodzą zaś przewód v^ątrobowy oraz naczynia limfatyczne. Dwie inne bruzdy - podłużna lewa i podłużna prawa - przebiegają w kierunku strzałkowym. Każda z tych bruzd dzieli się na dwie części przednią i tylną. W części przedniej bruzdy lewej przebiega więzadło obłe w ą t r o b y (ligamentum teres hepatis), będące pozostałością żyły pępkowej. W części tylnej znajduje się więzadło żylne - pozostałość po przewodzie żylnym (ductus venosuś), który w życiu p ł o d o w y m łączył żyłę pępkową z żyłą główną dolną. Część przednia bruzdy prawej nazywa się dołem pęcherzyka żółciowego ( fossa vesicae felleae), gdyż leży tu pęcherzyk żółciowy. Część
Więzadło wieńcowe Więzadło trójkątne lewe
Więzadło trójkątne prawe
Pęcherzyk żołdowy , Ryc. 113. Więzadła wątroby.
197
tylna nosi nazwę bruzdy żyły g ł ó w n e j dolnej ( sulcus venae cavae inferioris), która tędy przebiega. Wymienione bruzdy wyznaczają na powierzchni trzewnej (dolnej) wątroby granice czterech płatów. Na lewo od rowka podłużnego lewego znajduje się l e w y płat w ą t r o b y (lobus sinister hepatis). Pomiędzy dołem pęcherzyka żółciowego a więzadłem obłym leży płat czworoboczny ( lobus quadratus), Płatem ogoniastym (lobus caudatus) nazywa się część położona pomiędzy bruzdą żyły głównej dolnej a więzadłem żylnym. Część powierzchni trzewnej położona na prawo od bruzdy podłużnej prawej należy do prawego płata w ą t r o b y (lobus dexter). Powierzchnia przeponowa wątroby dzieli się na części: przednią, górną, tylną i prawą. Części te oddzielone są od siebie niezbyt wyraźnymi zaokrąglon y m i brzegami. Jedynie powierzchnia przednia odgraniczona jest od trzewnej (dolnej) wyraźnym brzegiem d o l n y m (margo inferioi•). . Wątroba otoczona jest błoną łącznotkankową, zwaną torebką w ą t r o b y . Torebka ta wnika razem z przewodem w ą t r o b o w y m i naczyniami do miąższu wątroby, tworząc wewnątrz gruczołu zrąb dla tkanki gruczołowej wątrobowej. Końcowe wyrostki tej torebki, w postaci włókien podporowych dla naczyń włosowatych, noszą nazwę w ł ó k i e n kratowych. Większa część powierzchni wątroby pokryta jest otrzewną, która jest ściśle zrośnięta z torebką wątroby. Jedynie część powierzchni przeponowej płata prawego w otoczeniu żyły głównej dolnej, zrośnięta z przeponą, oraz część powierzchni dolnej, przylegająca do pęcherzyka żółciowego, nie mają pokrywy otrzewnowej. Otrzewna przechodząca z wątroby na przeponę tworzy poprzecznie biegnące więzadło w i e ń c o w e (ligamentum coronarium hepatis); wotne brzegi prawy i lewy tego więzadła ze względu na ich kształt nazywa się wiązadłami t r ó j k ą t n y m i p r a w y m i l e w y m (ligamentum triangulare dextrum etsinistrum). Więzadłem s i e r p o w a t y m w ą t r o b y ( l i g a m e n t u m falciforme hepatis) nazywa się szeroki fałd otrzewnej, który odchodzi od górnej i przedniej części powierzchni przeponowej wątroby do dolnej powierzchni przepony oraz do przedniej ściany jamy brzusznej aż do pępka. Więzadło to leży w płaszczyźnie strzałkowej. Linia przyczepu więzadła sierpowatego wątroby wyznacza na jej powierzchni granicę oddzielającą płat prawy od płata lewego. Dolny brzeg więzadła sierpowatego jest wolny. Biegnie w nim pasmo włókniste, zwane więzadłem o b ł y m w ą t r o b y (ligamentum teres hepatis), będące pozostałością istniejącej w życiu p ł o d o w y m żyły p ę p k o w e j (v. umbilicalis). Podwójny fałd otrzewnej odchodzący od w r ó t wątroby ku krzywiźnie mniejszej żołądka i początkowej części dwunastnicy nosi nazwę sieci mniejszej (omentum minus). W sieci tej rozróżniamy dwie części, zwane wiązadłami w ą t r o b o w o - d w u n a s t n i c z y m i w ą t r o b o w o - ż o ł ą d k o w y m . W więzadle wątrobowo-dwunastniczym przebiega przewód żółciowy wspólny, tętnica wątrobowa, żyła wrotna, naczynia limfatyczne i nerwy zdążające do wątroby. Na utrzymanie stosunkowo ciężkiego narządu, jakim jest wątroba, w jego położeniu w górnej części jamy brzusznej pod przeponą składa się kilka 198
Najważniejszą rolę w tym względzie odgrywa tłocznia brzuszna, żyła główna dolna oraz tkanka łączna, wiążąca część powierzchni wątroby z przeponą. Udział więzadeł w utrzymywaniu wątroby jest mały. czynników.
Budowa mikroskopowa wątroby Z różnorodnymi czynnościami, jakie pełni w ustroju wątroba, pozostaje w związku jej swoista budowa, różniąca się znacznie od budowy innych gruczołów nabłonkowych. Wątroba wydziela żółć, bierze udział w przemianie mąterii, pełni rolę obronną, oczyszczając krew z substancji szkódIiwych, wytwarza mocznik, magazynuje węglowodany w postaci glikogenu oraz częściowo tłuszcze. Poza tym jest magazynem krwi oraz narządem niszczącym starzejące się krwinki czerwone. Do wątroby dochodzi nie tylko krew tętnicza, lecz także krew żylna, prowadzona przez żyłę wrotną z narządów jamy brzusznej. Budowa wewnętrzna wątroby w znacznej mierze zależna jest od właściwości jej ukrwienia. Wątroba składa się z wielkiej liczby małych zrazików o kształcie wielokątnych pryzmatów, średnicy 1 - 2 mm, ściśle do siebie przylegających, połączonych tkanką łączną. 0 człowieka rozgraniczenie poszczególnych zrazików nie jest wyraźne, a ich zasięg rozpoznajemy pod mikroskopem według charakterystycznego układu naczyń. Natomiast niektóre zwierzęta, jak np. świnia, mają bardzo wyraźnie oddzielone zraziki wątrobowe. Tętnica wątrobowa i żyła wrotna wchodzące przez wrota wątroby dzielą się na coraz mniejsze gałązki. Ostatecznie gałązki te przebiegają pomiędzy po-
Kanalik żółciowy
Odgałęzienie żyły wrotnej
Ryc. 114. Budowa zrazika wątrobowego.
199
d
szczególnymi zrazikami, nosząc nazwę naczyń międzyzrazikowych. Od gałązek międzyzrazikowych żyły wrotnej odchodzą żylne naczynia włosowate, zdążające promienisto do środka każdego zrazika, gdzie wpadają do tzw. żyły ś r o d k o w e j zrazika (v. centralis), która biegnie w jego osi. Żyły środkovye poszczególnych zrazików, łącząc się ze sobą w coraz to większe naczynia, tworzą ostatecznie kilka żył w ą t r o b o w y c h (w. hepaticae), które wpadają do żyły głównej dolnej w pobliżu tylnego brzegu w ą t r o b y pod przeponą. Każdy zrazik utworzony jest z wielkiej liczby komórek wątrobowych, które są ułożone w tzw. beleczki, przebiegające promieniście dokoła osi zrazika. Są to wielokątne komórki nabłonkowe o okrągłym jądrze i drobnoziarnistej iprotoplazmie, w której zawarte są barwniki żółciowe, kropelki glikogenu i tłuszczu. Wspomniana wyżej sieć naczyń włosowatych żylnych przebiega pomiędzy beleczkami komórek wątrobowych, przy czym śródbłonkowa ściana tych naczyń jest niekompletna, wskutek czego komórki w ą t r o b o w e w wielu miejscach pozostają w bezpośrednim kontakcie z krwią. Niektóre komórki śródbłonkowe mają postać tzw. komórek gwiaździstych. Należą one do układu siateczkowo-śródbłonkowego, pełniącego w ustroju ważną rolę w procesach obronnych. Beleczki wątrobowe są złożone z dwóch warstw komórek, pomiędzy którymi znajdują się wąskie włosowate kanaliki żółciowe; do nich komórki w ą t r o b o w e wydzielają żółć. Kanaliki te nie mają własnych ścian, lecz stanowią właściwie szczeliny pomiędzy sąsiednimi komórkami. Mają one, podobnie jak sieć naczyń żylnych, kierunek promienisty od środka ku o b w o d o w i zrazika. Sieć kanalików żółciowych jest jednak niezależna od sieci naczyniowej. Naczynia krwionośne włosowate biegną wzdłuż krawędzi komórek wątrobowych, kanaliki zaś - wzdłuż ich p o w i e ^ c h n i . Kanaliki żółciowe wpadają do przewodzików żółciowych międzyzrazikowych, których ściany wyścieła nabłonek walcowaty. Przewody żółciowe łącząc się ze sobą tworzą ostatecznie przewód wątrobowy wspólny, który wychodzi z w r ó t wątroby. Zrąb zrazików, na których ułożone są komórki wątrobowe, utworzony jest przez sieć włókien podporow y c h , będących wypustkami końcowymi torebki włóknistej.
Drogi żółciowe i pęcherzyk żółciowy Kanaliki żółciowe tworzą ostatecznie dwa przewody w ą t r o b o w e : większy, wychodzący z prawego płata, i mniejszy - z lewego. We wrotach wątroby . przewody te łączą się ze sobą pod kątem rozwartym tworząc tzw. p r z e w ó d w ą t r o b o w y w s p ó l n y (ductus hepaticus communls). Przewód ten, długości 25-30 m m , średnicy ok. 4 m m , biegnie w wiązadle Wątrobowo-dwunastniczym na prawo i ku d o ł o w i i łączy się pod kątem ostrym z p r z e w o d e m pęcherzykowym (ductus cysticus) wychodzącym z pęcherzyka żółciowego. Pęcherzyk żółciowy ( vesica fellea) wraz z przewodem pęcherzykowym można rozpatrywać jako uchyłek przewodu wątrobowego, służący za zbiornik 200
„
żółci, w którym ulega ona zagęszczeniu wskutek wchłaniania w o d y przez błonę śluzową. Pęcherzyk żółciowy ma kształt gruszkowatego worka długości 8-12 cm, szerokości 30-50 m m , pojemności 30-50 ml. Rozróżniamy w nim trzon (corpus vesicae felleae), dno (fundus) i szyjkę ( c o l l u m ) . Dno pęcherzyka żółciowego
/
Trzon pęcherzyka żółciowego
Szyjka pęcherzyka żółciowego }
Przewód pęcherzykowy < r—Przewód wątrobowy Pęzewód —żółciowy wspólny
Ryc. 115. Pęcherzyk żółciowy.
Trzon leży w opisanym wyżej dole pęcherzyka żółciowego. Dno pęcherzyka rzutuje na przednią ścianę jamy brzusznej w kącie pomiędzy brzegiem bocznym mięśnia prostego brzucha i łukiem żebrowym prawym. Dno wystaje nieco spod dolnego brzegu wątroby. Szyjka, skierowana ku w r o t o m wątroby, zwęża się stopniowo w kierunku przewodu w ą t r o b o w e g o wspólnego i ma dwa esowate wygięcia. W skład ściany pęcherzyka żółciowego wchodzi błona śluzowa i błona mięśniowa. Błona surowicza pokrywa tylko dolną jego powierzchnię. Powierzchnia błony śluzowej tworzy drobną sieć niewielkich fałdów. W obrębie przewodu pęcherzykowego fałdy układają się poprzecznie, tworząc tzw. fałd spiralny (plica spiralis). Błona mięśniowa jest słabo rozwinięta. Przewód pęcherzykowy ( ductus cysticus) jest nieco dłuższy od przewodu w ą t r o b o w e g o wspólnego (30—35 mm), o mniejszej średnicy (3 mm). Przewód żółciowy w s p ó l n y (ductus choledochus), długości ok. 75 m m , średnicy zmiennej (ok. 8 mm), powstaje po połączeniu przewodu wątrobowego wspólnego z przewodem pęcherzykowym. Końcowy odcinek przewodu żółciowego wspólnego spotyka się z przewodem trzustkowym przed wejściem do ściany dwunastnicy, którą wspólnie przebijają ukośnie i otwierają się 201
na brodawce większej dwunastnicy, w odległości 8,5-10 cm od odźwiernika. Przed ujściem oba przewody tworzą rozszerzenie, zwane bańką w ą t r o b o w o -trzustkową, leżącą w obrębie brodawki dwunastniczej. W ujściu prowadzącym z bańki do. dwunastnicy znajduje się zwieracz, który reguluje przepływ wydzieliny do dwunastnicy. Nosi on nazwę zwieracza bańki w ą t r o b o w o - t r z u s t k o w e j ( sphincter ampullae hepatopancreaticae). j Cholecystografia. Układ dróg żółciowych daje się uwidocznić na zjdjęciu rentgenowskim, jeżeli badanemu podamy doustnie lub dożylnie pewne związki jodu, które są wiązane w wątrobie i w y d a l a n e j żółcią. Badanie takie, zwane cholecystografią, umożliwia rozpoznanie kamicy żółciowej. /
Czynności w ą t r o b y Czynności wątroby są bardzo różnorodne. Poza udziałem w procesach trawienia, o m ó w i o n y m dokładnie w innym miejscu (str. 209), wątroba bierze udział w gospodarce węglowodanowej, w przemianie białkowej i tłuszczowej, pełni rolę ochronną oraz wykonuje wiele innych czynności, niedostateczriie jeszcze poznanych. W wątrobie odbywa się przemiana glukozy (cukru gronowego) na glikogen, czyli skrobię zwierzęcą, który jest magazynowany wJ
202
Trzustka Trzustka ( pancreas) jest długim i wąskim gruczołem o budowie zrazikowej, barwy szaroróżowej lub żółtoróżowej, konsystencji miękkiej, o ciężarze ok. 70-90 g. Długość jej wynosi 15-20 cm, szerokość 2,5-5,5 cm, grubość 1,7-2 cm. Trzustka jest ułożona poprzecznie na tylnej ścianie jamy brzusznej za żołądkiem na poziomie I i II kręgu lędźwiowego. Rozróżniamy w niej: 1) g ł o w ę (caput pancreatis), położoną wewnątrz pętli dwunastniczej, 2) trzon ( c o r p u s pancreatis), ciągnący się poprzecznie ku stronie lewej i nieco ku górze, oraz 3) ogon ( cauda pancreatis), sięgający do wnęki śledziony. Głowa trzustki, nieco spłaszczona, jest zrośnięta z częścią zstępującą dwunastnicy. Trzon i ogon na przekroju poprzecznym mają kształt trójkątny, w związku z czym odróżniamy tu trzy powierzchnie. Powierzchnia przednia, pokryta w całości otrzewną, zwrócona jest ku przodowi i ku górze, tworząc łoże dla żołądka. Powierzchnia dolna, skierowana ku przodowi i dołowi, również pokryta otrzewną, styka się z krezką okrężnicy poprzecznej. Ściana tylna, zwrócona ku tyłowi, związana jest tkanką łączną wiotką z narządami położonymi na tylnej ścianie jamy brzusznej. Centralnie w osi gruczołu przebiega przewód t r z u s t k o w y ( ductus pancreaticus), do którego dochodzą mniejsze przewody zrazikowe. Przewód g ł ó w n y , jak i przewody zrazikowe, dają się łatwo rozpoznać, gdyż ściana ich ma barwę białawą, która odznacza się na tle ciemniejszego miąższu gruczołu. Przewód trzustkowy otwiera się do dwunastnicy razem z przewodem żółciowym w s p ó l n y m . Często istnieje jeszcze przewód dodatkowy, znacznie mniejszy i bardziej zmienny, który uchodzi około 2 cm powyżej ujścia przewodu głównego na brodawce dodatkowej.
Budowa w e w n ę t r z n a trzustki Trzustka ma b u d o w ę zrazikową. Liczne drobne rozgałęzienia przewodów zrazikowych w postaci długich, cienkich cewek łączą się z pęcherzykami gruczołowymi, umieszczonymi w zrazikach. Pęcherzyki wydzielnicze są małe, kuliste, wysłane wysokimi walcowatymi lub stożkowatymi komórkami gruczołowymi, które ograniczają bardzo wąskie światło. Wewnętrzna część tych komórek, zwrócona ku światłu pęcherzyków, jest jaśniejsza i zawiera dużo Ziarenek wydzieliny (zymogen), znikających po wydostaniu się wydzieliny do światła pęcherzyka. Zewnętrzna część komórek jest ciemniejsza i składa się z jednorodnej protoplazmy. Wśród tkanki w każdym zraziku rozsiane są grupy odmiennych komórek, tworzące w y s p y trzustki. Komórki te nie mają łączności z przewodami wyprowadzającymi, lecz są oplecione gęstą siecią naczyń krwionośnych włosowatych. Komórki beta tych w y s p wytwarzają h o r m o n insulinę, która jest bezpo203
średnio wydzielana do krwi. Wyspy trzustki tworzą tzw. aparat w y s e p k o w y trzustki, który należy do gruczołów dokrewnych i zostanie o m ó w i o n y dokładnie w odpowiednim rozdziale.
Czynności trzustki
T
Trzustka wytwarza dwa rodzaje wydzieliny: sok trzustkowy, wydzielany do dwunastnicy, który bierze znaczny udział w trawieniu pokarmów (p. str. 208), oraz powstające w wyspach trzustki dwa h o r m o n y - insulinę, która jest horm o n e m regulującym przemianę w ę g l o w o d a n o w ą (p. str. 524), i glukagon (czynnik hiperg i ikerhićzny), pod noszący poziom cukru gronowego we krwj przez rozpad glikogenu.
OTRZEWNA Otrzewna ( p e r i t o n e u m ) jest cienką, gładką błoną surowiczą, która wyśćieła ściany jamy brzusznej i miednicy oraz pokrywa całkowicie lub częściowo położone w tych jamach narządy. Otrzewna stanowi jak gdyby worek, który u mężczyzn jest całkowicie zamknięty, a u kobiety łączy się ze światem zewnętrznym za pośrednictwem bardzo wąskiego ujścia jajowodu. Część otrzewnej wyściełającą od wewnątrz ściany jamy brzusznej i miednicy nazyw a m y otrzewną ścienną ( p e r i t o n e u m parietale), część zaś pokrywającą narządy - otrzewną trzewną ( p e r i t o n e u m viscerale). Otrzewna ścienna wyścieła warstwą ciągłą ścianę przednią, ściany boczne i przeponę. Na tylnej ścianie jamy brzusznej i na ścianach miednicy otrzewna ścienna przechodzi ze ściany na narządy i staje się otrzewną trzewną. W zależności od tego, w jakim stopniu narządy wewnętrzne są pokryte przez otrzewną, m ó w i m y o różnym ich położeniu albo stosunku do otrzewnej. Jedne narządy są pokryte otrzewną ze wszystkich stron, inne tylko częściowo, a do niektórych otrzewna przylega tylko z jednej strony. O pierwszych m ó w i m y , że są położone w e w n ą t r z o t r z e w n o w o . Należą tu: żołądek, jelito cienkie z wyjątkiem dwunastnicy, śledziona, wątroba, jelito ślepe, wyrostek robaczkowy, okrężnica poprzeczna, esica, górna część odbytnicy, macica. Do drugiej grupy należą narządy pierwotnie położone wewnątrzotrzewnowo; w trakcie rozwoju osobniczego znalazły się one wtórnie w położeniu częściowo pozaotrzewnowym, które nazywamy położeniem ś r ó d o t r z e w n o w y m ; należą tu: pęcherz moczowy, środkowa część odbytnicy, okrężnica wstępująca i zstępująca. Wreszcie położenie pozaotrzewnowe mają nerki, nadnercza, trzustka i dwunastnica. Otrzewna, schodząc ze ściany j a m y brzusznej na narządy, tworzy fałdy składające się z dwóch blaszek błony surowiczej, zwane krezkami (mesenterium) lub wiązadłami ( l i g a m e n t u m ) . Blaszki, z których składa się krezka lub więzadło, rozchodzą się przy brzegu narządu, do którego dochodzą i który 204
otaczają, tworząc jego błonę surowiczą. Na krezce zawieszone jest jelito cienkie (z wyjątkiem dwunastnicy), okrężnica poprzeczna i okrężnica esowata. Krezka okrężnicy poprzecznej (mesocolon transversum) jest ułożona poprzecznie wzdłuż brzegu trzustki. Zgięcia okrężniczę lewe i prawe łączą się z przylegającą do nich przeponą za pośrednictwem więzadeł przeponowo-okrężniczych lewego i prawego ( l i g a m e n t u m phrenicocolicum sinistrum et dextrum). Fałd otrzewnej przechodzący z przepony na przeponową powierzchnię wątroby nazywa się więzadłem w i e ń c o w y m w ą t r o b y ( l i g a m e n t u m coronarium hepatis). Przednią ścianę jamy brzusznej łączy z wątrobą więzadło sierpowate ( l i g a m e n t u m falciforme hepatis). Pomiędzy w r o t a m i wątroby a krzywizną małą żołądka i górną częścią dwunastnicy rozpięta jest tzw. sieć mniejsza (omentum minus), składająca się z dwóch więzadeł: w ą t r o b o w o -dwunastniczego i wątrobowo-żołądkowego. Więzadło wątrobowo-żołądkowe dzieli się wzdłuż krzywizny małej żołądka na dwie blaszki, które pokrywają przednią i tylną powierzchnię żołądka, po czym znów łączą się wzdłuż krzywizny większej żołądka, by utworzyć tzw. sieć większą (omentum majus). Dwie blaszki sieci większej zwisają od krzywizny większej żołądka do spojenia łonowego, pokrywając od przodu pętle jelita cienkiego, po czym zawijają się ku górze i zrastają się ze sobą i z krezką okrężnicy poprzecznej. Przestrzeń położona za siecią mniejszą i żołądkiem nazywa się torbą sieciową (bursa omentalis). Wejście do tej przestrzeni stanowi tzw. o t w ó r sieciowy (foramen epiplóicum), położony poza więzadłem wątrobowo-dwunastniczym. TRAWIENIE POKARMÓW W JELICIE CIENKIM
Ruchy jelita cienkiego Ruchy jelita cienkiego mają do spełnienia dwa podstawowe zadania: 1) możliwie dokładne przemieszanie miazgi pokarmowej z sokami trawiennymi i zetknięcie strawionego pokarmu z powietrzchnią chłonną jelita i 2) przesuwanie z odpowiednią szybkością miazgi pokarmowej w kierunku odbytu. Zadania te spełnia błona mięśniowa. Jej warstwa podłużna kurcząc się skraca określony odcinek jelita i powiększa jego średnicę, zaś warstwa okrężna zwęża światło jelita, co prowadzi do przesunięcia treści do dalszego odcinka jelita. Do czynników pobudzających mięśniówkę należą: 1. Mechaniczne podrażnienie - rozciąganie przez pokarm; im szybciej odbywa się rozciąganie, t y m silniejsze są ruchy; 2. Podrażnienie chemiczne błony śluzowej przez soki trawienne i substancje pokarmowe - kwasy, ługi i sole - w p ł y w a w różny sposób na motorykę jelit: słabe roztwory kwasów, ługu, mydła wzmagają, silniejsze roztwory kwasów, zwalniają i hamują ruchy jelit; 205
3. Bodźce nerwowe nadchodzące do jelit drogą nerwu błędnego i współczulnego: nerw błędny pobudza ruchy jelit i zwiększa napięcie mięśniówki, nerw współczulny działa antagonistycznie - drażnienie nerwów trzewnych zwalnia ruchy jelitowe i zmniejsza napięcie ich ściany. Bodźce te tłumaczą wpływy różnych stanów emocjonalnych na motorykę jelit; strach może wywołać burzliwe skurcze jelit (biegunka nerwowa). Należy jeszcze zaznaczyć, że mięśniówka jelita cienkiego poza oddziaływaniem na podrażnienia, z zewnątrz obdarzona jest automatyzmem, który przejawia się w rytmicznych skurczach nawet po przecięciu wszystkich połączeń z ośrodkowym układem nerwowym.
206
Rozróżniamy trzy rodzaje ruchów jelita: 1) ruchy wahadłowe, 2) ruch robaczkowy (perystaltyczny) i 3) ruchy rozdzielcze (segmentacyjne). 1. Ruchy w a h a d ł o w e polegają na rytmicznym wydłużaniu się i kurczeniu lub skracaniu odcinka jelita wskutek przesuwania się (2,5 cm/s) łagodnej fali skurczu wzdłuż jelita na krótkim odcinku 10-12 razy na minutę. 2. Ruch robaczkowy odbywa się w ten sposób, że w p e w n y m miejscu, położonym za masą pokarmową, następuje skurcz mięśniówki podłużnej wywołujący skrócenie i rozszerzenie danego odcinka jelita. Natychmiast po t y m skurczu w odcinku przed masą pokarmową następuje skurcz mięśniówki okrężnej, który wyciska treść jelita w kierunku odbytu. Skurcz ten przesuwa się naprzód jako tzw. fala perystaltyczna. Jednocześnie wzdłuż jelita przebiega kilka takich fal, które nadają ruchom jelita wygląd poruszającej się dżdżownicy. Szybkość posuwania się fali perystaltycznej zależna jest od czynników w y m i e nionych wyżej. W niektórych przypadkach może nastąpić zmiana kierunku posuwania się fali perystaltycznej, co nazywamy antyperystaltyką - zawartość jelita zostaje przesunięta w kierunku żołądka. 3. Ruchy rozdzielcze (segmentacyjne) polegają na silnych skurczach mięśniówki okrężnej w różnych odcinkach jelita przy biernym rozluźnieniu mięśniówki podłużnej. W momencie następnym skurczone odcinki jelita ulegają rozluźnieniu, a skurcze pojawiają się w innych miejscach. Wskutek takiej rytmicznej segmentacji miazga pokarmowa zostaje podzielona na szereg mas wtórnych, co ułatwia jej wymieszanie z sokami trawiennymi. Wszystkie opisane wyżej ruchy odbywają się przy t r w a ł y m napięciu ( tonus) ściany mięśniówki jelita.
Trawienie chemiczne w jelicie cienkim Miazga pokarmowa (chymus) nadchodząca z żołądka do dwunastnicy nie zawiera już grubszych cząstek pokarmowych i wskutek zawartości kwasu solnego ma odczyn kwaśny. Pod w p ł y w e m soku dwunastniczego odczyn ten zmienia się na zasadowy, niezbędny dla czynności enzymów jelita cienkiego. Większość składników pokarmowych nie jest jeszcze całkowicie strawiona. W dwunastnicy pod w p ł y w e m soku trzustkowego, żółci oraz wydzieliny gruczołów dwunastniczych i jelitowych pokarm podlega złożonym procesom chemicznym, w wyniku których następuje rozszczepienie białek, węglowodanów i tłuszczów. Dalszy ciąg tych samych procesów odbywa się w jelicie czczym i krętym pod w p ł y w e m soku jelitowego, wydzielanego przez gruczoły jelitowe. Należy podkreślić, że trawienie jest procesem ciągłym - pokarm, który się znalazł w jelicie cienkim, podlega równoczesnemu działaniu wszystkich enzym ó w trzustkowych i jelitowych. Wyodrębnienie z soków trawiennych poszczególnych enzymów, z których każdy daje się badać oddzielnie, przyczyniło się do wytworzenia poglądu, że trawienie odbywa się etapami. 207
Aby zapoznać się bliżej z tymi procesami, musimy kolejno rozpatrzyć właściwości poszczególnych soków trawiennych, które działają na miazgę pokarmową w jelicie. Sok trzustkowy jest płynem wodojasnym, bezbarwnym, o odczynie zasadowym. U człowieka wytwarza się na dobę ok. 1200 ml soku trzustkowego. Odczyn zasadowy nadaje mu głównie wodorowęglan sodowy. W skład soku trzustkowego wchodzą trzy grupy enzymów, które hydrolizują białka, węglowodany i tłuszcze. Są to: 1) proteazy: trypsyna i erepsyna, 2) amylaza trzustkowa (diastaza) i 3) lipaza, zwana także steapsyną. Enzymy proteolityczne, trypsyna [erepsyna, rozkładają cząsteczki białka na aminokwasy. Trypsyna jest wydzielana w postaci nieczynnej, zwanej trypsynogenem. Trypsynogen zamienia się na czynny enzym pod wpływem swojstego enzymu zawartego w soku jelitowym, zwanego enterokinazą. Enterokinaza jest zatem jak gdyby „enzymem enzymu". Do uaktywnienia trypsynogenu potrzebna jest bardzo niewielka ilość enterokinazy. Trypsyna rozkłada zarówno białka, jak i dalsze produkty ich rozpadu albumozy i peptony; erepsyna działa natomiast tylko na peptony wytworzone w żołądku pod wpływem pepsyny. Erepsyna wydzielana jest od razu w postaci czynnej. Trypsyna jest'zatem drugim z kolei enzymem proteolitycznym, jaki napotykamy w przewodzie pokarmowym. Działanie jej jednak nie polega na zwykłym dublowaniu roli pepsyny żołądkowej. W przeciwieństwie do niej trypsyna rozwija większą aktywność w środowisku słabo zasadowym, działa znacznie wolniej na pewne gatunki białek oraz doprowadza rozpad wielu białek do końca, tj. do aminokwasów. Amylaza trzustkowa (diastaza) jest enzymem amylolitycznym, działającym podobnie jak ptialina ślinowa. Rozkłada wielocukry do maltozy, ta zaś pod wpływem swoistego enzymu maltazy, zawartej w soku jelitowym, ulega przemianie na cukier gronowy (glukozę). Lipaza trzustkowa (steapsyna) rozkłada tłuszcze na glicerol i kwasy tłuszczowe. Najsilniejsze działanie rozwija lipaza w środowisku zbliżonym do obojętnego. Kwas solny niszczy lipazę. Czynność lipazy wzmaga się znacznie pod wpływem żółci. Proces hydrolizy poprzedzony jest zemulgowaniem tłuszczów. Odbywa się to w ten sposób, że po rozbiciu pewnej ilości tłuszczów przez lipazę powstające kwasy tłuszczowe tworzą z wodorowęglanem sodow y m mydła, które przyczyniają się do emulgowania tłuszczów. Tłuszcze pod postacią takiej emulsji (zawiesiny drobnych kuleczek) mają znacznie większą powierzchnię, co bardzo ułatwia dalsze działanie lipazy. Dzięki zawartości wymienionych wyżej enzymów sok trzustkowy odgrywa w procesie trawiennym bardzo ważną rolę. Wydzielanie soku trzustkowego odbywa się w 3 - 5 min po spożyciu pokarmu i trwa 6-14 h, zależnie od składu pokarmu. Mechanizm wydzielania soku trzustkowego jest dwojakiego rodzaju: nerw o w y i humoralny. Wydzielanie nerwowe odbywa się pod wpływem impulsów przekazywanych przez nerw błędny równocześnie zfazą nerwową wydzielania żołądkowe208
go. Sok trzustkowy wytwarzany w fazie nerwowej jest gęsty, bogaty w enzymy i substancje białkowe, zawiera mało wody i elektrolitów. Znacznie większe znaczenie w czynności trzustki ma mechanizm humoralny. Istnieją dwa hormony pobudzające trzustkę do wydzielania soków trawiennych: sekretyna i pankreozymina. Sekretyna jest to niewielki polipeptyd znajdujący się w błonie śluzowej górnego odcinka jelita cienkiego. Wyzwalanie i aktywacja sekretyny następuje pod wpływem nadchodzącej do jelita miazgi pokarmowej, najsilniejszym jednak bodźcem jest zawarty w niej kwas solny. Zaktywowana sekretyna dochodzi do trzustki drogą krwionośną pobudzając ją do wydzielania dużej ilości płynu o wysokim stężeniu jonu wodorowęglanowego (do 145 mEq/l). W myśl reakcji chemicznej: HCI + NaHC0 3 -*NaCI + H 2 C0 3 następuje zobojętnienie kwasu solnego, a powstający zamiast niego kwas węglowy ulega szybko dysocjacji na dwutlenek węgla i wodę. Wskutek dalszego wydzielania wodorowęglanu sodowego zawartość dwunastnicy uzyskuje odczyn lekko zasadowy, optymalny dla działania enzymów trzustkowych. Drugi hormon wydzielany przez błonę śluzową jelita pod wpływem bodźca pokarmowego - głównie produktów częściowego trawienia białek - pobudza trzustkę do wydzielania dużej ilości enzymów trawiennych. Żółć jest produktem czynności wydzielniczej komórek wątrobowych. Wytwarzana przez komórki żółć przechodzi do światła kanalików żółciowych, a następnie do przewodu żółciowego wspólnego, skąd dostaje się bezpośrednio do dwunastnicy albo przez przewód pęcherzykowy do pęcherzyka żółciowego. Żółć jest wytwarzana przez komórki nieustannie, ale do^dwunastnicy wydalana jest okresowo w czasie przesuwania się treści pokarmowej z żołądka do jelita. W przerwach między trawieniem żółć gromadzi się w pęcherzyku żółciowym. Tutaj następuje wchłanianie wody przez błonę śluzową pęcherzyka, wskutek czego żółć ulega zagęszczeniu. W związku z tym żółć pęcherzyka różni się od żółci wątrobowej znacznie większą zawartością ciał stałych. Równocześnie pęcherzyk reguluje ciśnienie w kanalikach żółciowych. Ilość dobowa żółci wynosi u człowieka 500-1000 ml. Do najważnjejszych składników żółci należą kwasy żółciowe i barwniki żółciowe. W żółci człowieka występują głównie kwasy glikocholowy i taurocholowy w postaci soli sodowych. Barwnikiem żółciowym jest bilirubina. Bilirubina powstaje z produktu rozpadu krwinek czerwonych - hemoglobiny. Bilirubina ma barwę czerwonopomarańczową. Poza tym w skład żółci wchodzą: lecytyna, cholesterol, tłuszcze i mydła oraz mucyna. Rola żółci w procesach trawienia polega na: 1) aktywowaniu enzymów trzustkowych, przede wszystkim lipazy; 2) emulgowaniu tłuszczów - kwasy żółciowe zmniejszają napięcie powierzchniowe tłuszczów, wskutek czego rozbijają się one na drobne kuleczki, co ułatwia działanie lipazy; 3) rozpuszczaniu kwasów tłuszczowych; 4) wzmaganiu perystaltyki jelit. Z powyższego wynika, że najważniejsze zadania przypadają żółci w procesie 14 - Anatomia i fizjologia
209
trawienia tłuszczów. Zaburzenie w wydzielaniu żółei pociąga za sobą zmniejszenie przyswajania tłuszczów. Żółć wytwarza się w wątrobie nieustannie. W przerwach pomiędzy posiłkami ujście przewodu żółciowego wspólnego do dwunastnicy jest jednak zamknięte wskutek stosunkowo silnego napięcia mięśnia zwieracza bańki wątrobowo-trzustkowej, a żółć napływa do rozluźnionego pęcherzyka żółciowego. Po spożyciu pokarmu mała ilość żółci zostaje wstrzyknięta z pęcherzyka do dwunastnicy na drodze odruchowej, jednakże dokładniejsze opróżnienię pęcherzyka żółciowego następuje później pod wpływem hormonu pankreozyminy. Pankreozymina wytwarza się w śluzówce dwunastnicy pod wpływem kontaktu z treścią żołądkową. Dostając się do krwi powoduje skurcze i opróżnienie pęcherzyka żółciowego. Po opróżnieniu się pęcherzyka żółć spływa do dwunastnicy bezpośrednio z wątroby dopóty, dopóki trwa okres trawienia, po czym następuje stopniowy wzrost napięcia zwieracza bańki wątrobowo-trzustkowej i ponowne gromadzenie się żółci w pęcherzyku. Otwarcie ujścia przewodu żółciowego do dwunastnicy następuje również po słabym zadrażnieniu nerwu błędnego, natomiast silne zadrażnienie tego nerwu powoduje skurcz zwieracza bańki wątrobowo-trzustkowej. Przeciwnie działają bodźce doprowadzane przez nerw współczulny. Jest rzeczą ciekawą, że unerwienie pęcherzyka żółciowego jest sprzężone z unerwieniem zwieracza bańki wątrobowo-trzustkowej w ten sposób, że skurczowi pęcherzyka towarzyszy rozluźnienie zwieracza i odwrotnie.
WCHŁANIANIE Aby końcowe produkty trawienia pokarmu mogły być wykorzystane przez organizm, muszą się one przedostać z przewodu p o l ^ r m o w e g o ^ k j w j , która rozniesie je po całym ustroju. Procesy, które dó tego prowadzą, nazywamy wchłanianiem. Wchłanianie odbywa się w ścianie przewodu pokarmowego i jest zależne od wielkości powierzchni chłonącej, od stężenia składników podlegających wchłonięciu, od czasu, w jakim się te składniki stykają z powierzchnią chłonną, i od szybkości, w jakim wchłonięte substancje zostają ze ściany jelita odprowadzone z krwią. Wszystkie te warunki najkorzystniej przedstawiają się w jelicie cienkim, które jest głównym miejscem procesów wchłaniania. Fałdy okrężne i kosmki jelitowe zwiększają powierzchnię chłonną jelita ok. 23 razy; ciągłość procesów trawienia pod wpływem enzymów zapewnia w jelicie duże stężenia końco;wych produktów podlegających wessaniu; czas, przez jaki przebywa miazga pokarmowa w jelicie cienkim, jest stosunkowo długi (kilka godzin), obfite ukrwienie jelita zapewnia szybkie odprowadzenie wchłoniętego materiału. O roli kosmków jelitowych w procesie wchłaniania wspomnieliśmy już przy opisie budowy jelita cienkiego. Należy tu dodać, że w kosmkach znajdują się włókna mięśni gładkich, dzięki którym mogą Się one rytmicznie kurczyć. 210
Ryc. 118. Schemat kosmka jelitowego.
Skurcze kosmków zaczynają się w jakiś czas po jedzeniu i są wywołane przez bodźce mechaniczne drażniące, zawarte w miazdze pokarmowej, działające więc ze strony jelita. Kurczące się kosmki pełnią rolę pewnego rodzaju pompy. Przy skurczu kosmka zawartość jego naczyń limfatycznych zostaje wyciśniętą w kierunku splotów położonych głębiej w podśluzówce. W ten sposób zostaje ułatwione przenikanie tych substancji, które są wchłaniane do układu limfatycznegó. Udział irfnych odcinków przewodu pokarmowego w procesach wchłaniania jest niewielki. W żołądku mogą się wchłaniać tylko bardzo w o l n o sole mineralne, alkohol i woda. W jelicie grubym wchłaniane są bardzo duże ilości wody. Końcowymi produktami trawienia są: cukry proste, powstające z różnych węglowodanów, aminokwasy - produkty rozpadu hydrolitycznego białek, oraz kwasy tłuszczowe j glicerol pochodzące z rozkładu tłuszczów. Wszystkie te substancje w odróżnieniu od tych, z których powstały, mają zdolność przechodzenia przez błony zwierzęce (dyfuzja). Rozpuszczalne w wodzie cukry proste i aminokwasy zostają wchłonięte do naczyń włosowatych krwionośnych układu żyły wrotnej i zanim przejdą do ogólnego krwiobiegu, dostają się najpierw do wątroby. Tłuszcze zostają wchłonięte do układu limfatycznego. Podczas trawienia pokarmu tłuszczowego naczynia limfatyczne jelita cienkiego wypełniają się mleczem ( chylus ) , który składa się z limfy obładowanej kuleczkami tłuszczu. Uwolnione wskutek trawienia kwasy tłuszczowe i glicerol w czasie przechodzenia przez błonę śluzową jelita ulegają w komórkach śluzówki ponownemu połączeniu na tłuszcze obojętne, które wchodzą do centralnego naczynia 211
limfatycznego kosmka i ostatecznie zostają odprowadzone do układu żylnego przez przewód piersiowy limfatyczny ( ductus thoracicus). Wchłonięte w ten sposób tłuszcze mają postać drobniutkich kuleczek, zwanych chylomikronami, o średnicy 0,5-1,0 ^im, pokrytych na powierzchni błoną złożoną z białek. Pewna część kwasów tłuszczowych i glicerolu przechodzi jednak do naczyń włosowatych i wędruje z krwią do wątroby. Po obfitym pożywieniu białkowym również część aminokwasów może przechodzić do naczyń limfatycznych.
Trawienie pokarmów w jelicie grubym Ruchy jelita grubego Ujście jelita cienkiego do jelita grubego pozostaje pod kontrolą zastawki krętniczo-kątniczej, która normalnie jest zamknięta wskutek stałego napięcia tonicznego mięśniówki okrężnej, tworzącej w t y m ujściu rodzaj mięśnia zwieracza. W momentach opróżnienia się żołądka jelito kręte zostaje pobudzone na drodze odruchowej do skurczów, które przesuwają półpłynną jego zawartość do jelita ślepego. Skurcze takie występują w odstępach 5-minutowych przez cały czas przebywania pokarmu w żołądku. W ten sposób napełnia się stopniowo jelito ślepe, okrężnica wstępująca i poprzeczna, przy czym te odcinki okrężnicy pozostają wciąż nieruchome. Czas, w jakim następuje wypełnienie tych części jelita grubego, jest różny u różnych osobników. Po wypełnieniu się jelita ślepego i wspomnianych odcinków okrężnicy powstają w nich lokalne skurcze, przy których pojawiają się i znikają wybrzuszenia ( haustra ), powodując dokładne wymieszanie ich zawartości. Inny rodzaj ruchów jelita grubego, zwany perystaltyką, pojawia się 3 - 4 razy na dobę i trwa tylko przez krótki czas. Ruchy te polegają na szybko następującym skurczu, który rozpoczyna się mniej więcej w połowie okrężnicy i w c i ą g u kilku sekund przesuwa zawartość do części zstępującej i okrężnicy esowatej. Ruchów tych zwykle nie odczuwamy. Udział jelita grubego w procesach trawiennych jest niewielki, gdyż pokarm zostaje strawiony i wchłonięty w głównej mierze już w jelicie cienkim. Sok wydzielany przez jelito grube zawiera dużo mucyny, ma odczyn słabo zasadowy i niez:awiera enzymów. Wtreści, jaka dostaje się do jelita grubego, znajduje się pewna ilość nie strawionego pokarmu. Pozostając przez dłuższy czas w jelicie grubym, nie strawione składniki ulegają dalszemu działaniu enzym ó w , które tu przechodzą razem z miazgą pokarmową z jelita cienkiego. W odróżnieniu od jelita cienkiego flora bakteryjna jelita grubego jest bardzo bogata. Pod w p ł y w e m działania bakterii w zasadowym środowisku jelita grubego następuje gnicie białek, które nie zostały strawione i wchłonięte w jelicie cienkim. Gnicie różni się od trawienia między innymi t y m , że rozpad jbiałj^ ^ - aminokwasy zostają rozbite i zamienione na związki prostsze. 212
Niektóre z nich, jak np. indol, skatol, fenole itp., są dla ustroju trujące. Część ich zostaje wydalona z kałem, część zaś wchłonięta do krwi dostaje się do wątroby, która zamienia je na związki mniej toksyczne, tworząc z nich połączenia estrowe, ostatecznie wydalane przez nerki z moczem. Zawartość tych związków w moczu wskazuje przeto na nasilenie procesów gnilnych w jelicie grubym.
Kał W jelicie grubym wskutek wchłaniania wody następuje zagęszczenie treści i zamiana jej na kał. W formowaniu mas kałowych dużą rolę odgrywa śluz wytwarzany przez błonę śluzową jelita grubego, który zlepia nie strawione resztki pokarmu. W skład kału wchodzi: 1) woda, 2) nie strawione i nie dające się strawić resztki pokarmu, 3) barwniki pochodzące z nie strawionego pokarmu oraz barwniki żółciowe, 4) duże ilości drobnoustrojów, 5) produkty działania bakterii, 6) cholesterol, 7) sole mineralne sodu, potasu, wapnia, magnezu i żelaza. Ilość kału zależna jest od ilości spożywanego pokarmu. Przy diecie roślinnej wytwarza się więcej kału niż przy pokarmie mięsnym. Cały, proces trawienia przy pokarmie roślinnym lub mieszanym trwa 1 - 2 doby, przy czym połowę tego czasu zajmuje przesuwanie się resztek pokarmowych w jelicie grubym.
Defekacja Oddawanie kału, czyli defekacja, rozpoczyna się od parcia na stolec, które zostaje wywołane przez rozciąganie ścian odbytnicy nadchodzącymi masami kałowymi. U większości ludzi odbytnica pozostaje pusta aż do chwili pojawienia się czucia parcia. W przerwach pomiędzy oddawaniem stolca ujście zewnętrzne odbytnicy jest zamknięte wskutek stałego tonicznego skurczu obydwóch mięśni zwieraczy odbytu. Z chwilą gdy rozciąganie odbytnicy osiągnie pewien stopień nasilenia, następują odruchowe skurcze jej ściany i rozluźnienie zwieraczy odbytu, przy czym kał zaczyna się wydobywać na zewnątrz. Do wydalenia kału przyczynia się wydatnie aktywne parcie tłoczni brzusznej, tj. wzrost ciśnienia w jamie brzusznej, powstający wskutek silnego wydechu przy zamkniętej głośni i skurczu mięśni brzucha. Defekacja kończy się skurczem mięśnia dźwigacza odbytu, który wciąga wynicowującą się przy tym błonę śluzową odbytnicy. Ośrodki nerwowe kierujące odruchem defekacji mieszczą się w odcinku krzyżowym rdzenia kręgowego. Zniszczenie tego ośrodka prowadzi do rozluźnienia mięśni zwieraczy odbytu i nietrzymania kału.
213
Przemiana materii i energii. Odżywianie Cechą szczególną procesów przemiany materii i energii w ustrojach żywych jest to, że odbywają się one w stosunkowo niskiej temperaturze (30-40°). Podobne reakcje przeprowadzone sztucznie wymagają długiego czasu, wysokiej temperatury albo silnie działających związków, jak silne kwasy lub ługi. Reakcje chemiczne w ustroju żywym przebiegają stosunkowo szybko i w niskiej temperaturze dzięki biologicznym katalizatorom - enzymom, które wzmagają i przyśpieszają przebieg reakcji chemicznych. Działanie enzymów jest swoiste, tzn. każdy enzym działa tylko na pewną substancję (substrat) lub grupę substancji. Enzymy muszą „pasować do substratu jak klucz do zamka." Enzymy rozwijają najsilniejsze działanie w temperaturze ciała (ok. 38°). Działanie różnych enzymów jest również zależne w znacznym stopniu od odczynu środowiska, w jakim się znajdują. Jedne działają najskuteczniej w odczynach kwaśnych, inne - w obojętnych lub słabo zasadowych. Procesy przemiany można podzielić na dwie grupy: 1) anabolizm, asymilacja i 2) katabolizm, dysymilacja. Anabolizmem nazywamy sumę procesów, w wyniku których komórki pobierają z krwi substancje pokarmowe i tworzą z nich część^własnej protoplazmy. Substancje nie mające cech życia zostają przy tym zamienione na żywą materię. O naturze zmian anabolicznych wiemy stosunkowo niewiele; najprawdopodobniej są to reakcje chemiczne odwrotne do hydrolizy, przy których małe cząsteczki zostają zamienione na duże z utratą wody. Katabolizmem nazywamy procesy rozpadu substancji wchodzących w skład protoplazmy. Duże cząsteczki zostają rozbite na związki prostsze, które zostają zużyte przez jnne komórki, przeważnie jednak wnątrz. Procesy kataboliczne polegają głównie na: 1) rozszczepianiu dużych cząsteczek na mniejsze, 2) hydrolizie, tj. rozbijaniu dużych cząsteczek na mniejsze z przyłączeniem wody, 3) utlenianiu. Substrat Substrat
Enzym
Enzym
a .J Enzym
Enzym
Ryc. 119. Mechanizm działania enzymu.
214
Produkty działania enzymu
Przemiana białka Białka są najważniejszymi składnikami komórek i płynów tkankowych. Są to związki niezwykle skomplikowane, odznaczające się bardzo dużą masą cząsteczkową. W skład cząsteczki białka może wchodzić wiele tysięcy atomów. Białka zawierają zawsze węgiel - C, wodór - H, tlen - O i azot - N, czasami siarkę, fosfor lub żelazo. Poddając cząsteczki białkowe reakcjom rozkładowym stwierdzono, że typowymi produktami rozpadu białek są stosunkowo proste związki - aminokwasy. Trawienie białek pokarmowych polega na serii przemian pod wpływem enzymów proteolitycznych zawartych w soku żołądkowym, soku trzustkowym i jelitowym, które doprowadzają do rozbicia wielkich cząsteczek na podstawowe składniki - aminokwasy. Aminokwasy są wchłaniane w jelicie cienkim, a przechodzenie ich przez komórki błony śluzowej ze światła jelita do krwi jest procesem czynnym wymagającym energii. Aminokwasy zostają przeniesione do wątroby z krwią żyły wrotnej, gdzie część ich ulega zatrzymaniu, część zaś przechodzi do krążenia ogólnego. Wątroba jest głównym, choć nie jedynym, miejscem syntezy i degradacji aminokwasów. Znaczenie fizjologiczne rozmaitych aminokwasów badano karmiąc zwierzęta doświadczalne różnymi białkami, w których pewne określone aminokwasy nie występują. Stwierdzono przy tym, że wyłączenie z pokarmu pewnych aminokwasów pociąga za sobą znaczne zaburzenia w czynnościach fizjologicznych ustroju. Okazało się, że ustrój człowieka nie potrafi wytwarzać pewnych aminokwasów i te muszą być doprowadzone z pokarmem. Niżej podajemy listę aminokwasów wyosobnionych z białkowego pokarmu człowieka. Rozstrzelonym drukiem są wydrukowane aminokwasy, które muszą być doprowadzone z zewnątrz:
glikokol alanina w a l i na leucyna izoleucyna arginina lizyna
norleucyna fenyloalanina tyrozyna seryna treonina histydyna cystyna
metionina kwas asparaginowy kwas glutaminowy kwas hydroksyglutaminowy prolina hydroksyprolina tryptofan
Aminokwasy służą do syntezy własnego białka ustrojowego, stąd też największe zapotrzebowanie na pokarm białkowy istnieje w każdym ustroju rosnącym, budującym nowe komórki i tkanki. Ale również dojrzały organizm potrzebuje dość pokaźnych ilości aminokwasów na ten cel, gdyż pewna część białka zawartego w żywych komórkach ulega rozpadowi i musi być zastąpiona nowym. Aminokwasy nie użyte do budowy własnych tkanek, zostają rozłożone z wyzwoleniem energii. Pierwszym etapem tego rozkładu jestdezaminacja,tj. 215
oiafKoj
Karboksy-peptydaza Trypsyna Karboksypeptydaza
Ryc. 120. Schemat trawienia białka.
216
odszczepienie od reszty cząsteczki grupy aminowej zawierającej azot. Odłączony zostaje przy tym amoniak, który zostaje zamieniony na mocznik CO/ NH2/2- Synteza mocznika odbywa się w wątrobie i ma wielkie znaczenie, gdyż amoniak, gromadząc się we krwi, działa szkodliwie na ustrój, natomiast mocznik nie ma trujących właściwości. Mocznik jest wydalany z ustroju z moczem. Białko Jama ustna Enzymy żołądkowe Żołądek
Proteozy Enzymy trzustkowe
T
Jelito cienkie
Peptony
Peptony Enzvmv trzustkowe
Trójpeptydy
Polipeptydy
T
*
•
9
Dwupeptydy
T Aminokwasy Ryc. 121. Proces trawienia białka. Enzymy proteolityczne żołądka (pepsyna) rozkładają białko do proteoz. Enzymy trzustkowe działając na proteozy rozkładają je do peptonów i dalej do peptydów. Enzymy soku jelitowego rozbijają oligopeptydy na aminokwasy.
Po dezaminacji cząsteczki aminokwasów zostają spalone w taki sam sposób, jak węglowodany, przy czym końcowymi produktami są dwutlenek węgla i woda. Przy rozpadzie białek powstają zatem swoiste produkty zawierające azot. Obok mocznika, którego powstaje najwięcej, należą tu jeszcze kreatynina, kreatyna, kwas hipurowy. Wszystkie te odpadki przemiany białkowej są wydalane z moczem. Przeciętna zawartość azotu w białkach wynosi 16%. Jeżeli więc oznaczymy ilość azotu wydalonego na dobę z moczem, możemy łatwo obliczyć, jaka ilość białka uległa w ustroju rozpadowi. Oznaczając ilość azotu doprowadzonego z pożywieniem i wydalonego z moczem, możemy obliczyć ilość białka wprowadzonego i ilość rozłożonego w ustroju. Stosunek ilościowy azotu doprowadzonego do azotu wydzielonego z moczem nazywamy bilansem azotowym. W warunkach prawidłowych, w zdrowym dorosłym ustroju, wystarczająco odżywionym, ilość azotu doprowadzonego równa się ilości wydzielonego 217
z moczem. Mówimy, że organizm taki znajduje się w stanie równowagi azotowej. Inaczej przedstawia się bilans azotowy w ustroju, który zużywa duże ilości białka na budowę własnych tkanek, np. w ustroju rosnącym lub wtedy, gdy ustrój musi uzupełnić nadmierne straty białek, np. wstanie rekonwalescencji po przebyciu długotrwałych chorób wyniszczających lub po dłuższym okresie głodowania. W tych wypadkach ilość doprowadzonego białka przewyższa ilość białka rozpadającego się i bilans azotowy jest dodatni. ' Jeżeli rozpad białka jest większy niż jego dowóz z pokarmem, co występuje np. w ciężkich chorobach zakaźnych lub w okresie głodowania, mamy do ęzynienia z ujemnym bilansem azotowym. Nawet przy zupełnym wstrzymaniu dowozu białka z pokarmem ustrój stale wydala pewną ilość azotu wskutek rozpadu białka ustrojowego. Ilość ta wynosi u człowieka 13-20 g na dobę i nosi nazwę kwoty zużycia. Najmniejsza ilość białka doprowadzana wciągu doby z pokarmem, konieczna dla utrzymania równowagi ustrojowej azotowej, nazywa się minimum białkowym. Ilość ta jest większa od kwoty zużycia i wynosi u człowieka 30-40 g. To minimum białkowe zależne jest od rodzaju białka podawanego ustrojowi. Niektóre białka podawane nawet w największych ilościach nie doprowadzają do równowagi azotowej. Wspomnieliśmy już, że ustrój nie potrafi wytwarzać pewnych aminokwasów i że aminokwasy te, zwane niezbędnymi, muszą być dostarczone z pokarmem. W skład rozmaitych białek wchodzą różne rodzaje i różne ilości aminokwasów. Otóż białko ma tym większą wartość biologiczną dla ustroju, im więcej zawiera aminokwasów niezbędnych i im bardziej jego skład aminokwasowy zbliża się do składu białek ustroju. Ma to duże znaczenie dla praktyki odżywiania i z tego punktu widzenia dzielimy białka na pełnowartościowe, tj. takie, które zawierają wszystkie niezbędne aminokwasy, i na białka niepełnowartościowe, tj. takie, w których te aminokwasy występują w ilościach bardzo małych lub brak ich zupełnie.
Przemiana w ę g l o w o d a n ó w Węglowodany są głównym i najbardziej ekonomicznym źródłem energii. Wszystkie mięśnie szkieletowe, mięśnie gładkie oraz mięsień sercowy używają węglowodanów jako materiału energetycznego. W skład węglowodanów wchodzi węgiel (C), wodór (H), i tlen (O), przy czym ilościowy stosunek wodoru do tlenu jest taki sam, jak w wodzie, tj. 2:1, czemu związki te zawdzięczają swą nazwę. Wśród węglowodanów rozróżniamy cukry proste (monosacharydy) i cukry złożone, czyli wielocukry (polisacharydy). Cukry proste stanowią cząsteczki, o wzorze CeHi206. Są one rozpuszczalne w wodzie i mogą być wchłonięte bez zmiany przez organizm. Cukry proste są cegiełkami, z których zbudowane są wszystkie węglowodany. Najważniej218
szym dla ustroju cukrem prostym jest glukoza (cukier gronowy). Wszystkie wyższe ustroje zwierzęce czerpią energię z rozpadu tego właśnie cukru. Niezależnie od tego, w jakiej postaci węglowodany zostaną do ustroju doprowadzone, spalone zostają tylko wtedy, jeżeli ulegną zamianie na cukier prosty. W razie niedostatecznego dowozu węglowodanów z pokarmem organizm wytwarza glukozę z materiałów niecukrowych, przede wszystkim z aminokwasów. Dwucukrówce, o wzorze C12H22O11, są zbudowane z dwóch cząsteczek monosacharydów. Wielocukry, czyli polisacharydy, o wzorze (C6H1005)n są zbudowane z większej liczby grup jednocukrowych. Najpospolitszym wielocukrem roślinnym jest skrobia. Złożone wielocukry, zanim zostaną rozłożone na cukry proste, muszą ulec wielokrotnemu rozbiciu nazwiązki coraz prostsze o t y m samym względnym stosunku składników. Skrobia i inne węglowodany ulegające strawieniu zostają zamienione na cukry proste przez serię enzymów hydrolitycznych, do których należą: 1) amylaza ślinowa - rozbijająca skrobię na szereg oligosacharydów, 2) amylaza trzustkowa - bardzo aktywna - rozkładająca skrobię na dekstryny i maltozę, 3) maltaza trzustkowa - katalizująca hydrolizę maltozy na glukozę, 4) maltaza, sacharaza i laktaza, wydzielane w jelicie cienkim i rozkładające odpowiednie dwucukry - maltozę, sacharozę i laktozę. Wchłonięciu mogą ulec tylko cukry proste. Wchłanianie ich jest procesem czynnym. Po wchłonięciu w jelicie cukry te przechodzą do żyły wrotnej i dalej do wątroby. W wątrobie wszystkie cukry proste zostają zamienione na glukozę. Glukoza może przechodzić z wątroby do krążenia ogólnego i wszystkich tkanek, dla których stanowi główny materiał energetyczny, bądżteż może być zamieniona na glikogen. Glikogen jest wielocukrem - swego rodzaju „skrobią zwierzęcą", która odznacza się tym, iż jest łatwo dostępnym źródłem energii zmagazynowanej w wątrobie i w mięśniach. Zapasy glikogenu są jednak ograniczone i po ich wyczerpaniu ustrój zaczyna intensywniej wykorzystywać energię zgromadzoną w związkach tłuszczowych. Glikogen zmagazynowany w tkance mięśniowej z powrotem do krwi już nie wraca, lecz zostaje spalony w mięśniach do dwutlenku węgla i wody lub zamieniony na kwas mlekowy z wyzwoleniem energii. Dopiero kwas mlekowy powstający w mięśniach wraca z powrotem do wątroby, która jest zdolna zamienić go ponownie na glikogen. W razie braku węglowodanów w pokarmie wątroba wytwarza glukozę z aminokwasów. Nadmiar glukozy nie ulegającej spaleniu w ustroju, zostaje zamieniony na kwasy tłuszczowe i w dalszym etapie - na tłuszcz, który jest magazynowany przez ustrój jako zapasowy materiał energetyczny.
220
Cząsteczki glukozy
Ryc. 123. Proces rozkładu węglowodanów (skrobi) przez enzymy trawienne (amylazy) śliny, soku trzustkowego i soku jelitowego.
221
Regulacja poziomu cukru we k r w i Poziom glukozy we krwi odznacza się dużą stałością i wynosi 80-120 mg/100 ml. Utrzymanie stałego poziomu cukru we krwi jest sprawą niezwykle ważną dla ustroju. Znaczniejsze obniżenie poziomu cukru we krwi (poniżej 50 mg/100 ml.) powoduje u większości zwierząt objawy hipoglikemii - utratę przytomności, drgawki, a nawet śmierć. W utrzymaniu poziomu glukozy we krwi i w regulacji przemiany węglowodanów ogromną rolę odgrywają w p ł y w y gruczołów dokrewnych. Pierwsze miejsce zajmuje tu część wewnątrzwydzielnicza trzustki, wytwarzająca hormon - insulinę. Insulina zmniejsza poziom cukru we krwi dzięki następującym właściwościom: 1) 2) 3) 4)
sprzyja wytwarzaniu kwasów tłuszczowych z glukozy, przyczynia się do utleniania glukozy w narządach i tkankach, wzmaga syntezę glikogenu w wątrobie i mięśniach, zmniejsza wytwarzanie glikogenu z aminokwasów.
Hormony przedniego płata przysadki mózgowej i nadnercza oraz tarczycy mają tendencję do zwiększania poziomu cukru we krwi, działając antagonistycznie w stosunku do insuliny. Zaburzenie regulacji hormonalnej przemiany węglowodanów, powstające wskutek względnego lub bezwzględnego niedoboru insuliny, jest stanem chorobowym zwanym cukrzycą. W chorobie tej zawartość cukru we krwi wzrasta bardzo ponad normę wskutek te]go, że tkanki riie mogą go wykorzystać; glukoza, będąca cennym i wysokowartościowym źródłem energii, zostąje w dużych ilościach wydalana z moczem. Przemiana tłuszczów Tłuszcze są to estry glicerolu i kwasów tłuszczowych. Glicerol jest alkoholem trójwodorotlenowym, tłuszcz powstaje przez połączenie jednej cząsteczki glicerolu z trzema cząsteczkami kwasu tłuszczowego. W skład tłuszczów zwierzęcych wchodzą zwykle kwasy palmitynowy, stearynowy i oleinowy. Pokarm tłuszczowy, który nadchodzi do górnego odcinka jelita cienkiego, powoduje wydzielanie przez błonę śluzową jelita hormonu - cholecystokininy, którą, jak ostatnio stwierdzono, jest identyczna z pankreozyminą i pobudza pęcherzyk żółciowy do wyrzucenia żół
Trójglicerydy dłuqołańcuchowe
Ryc. 124. Schemat trawienia tłuszczów.
postać trójglicerydów, przeto głównym produktem działania lipazy są: glicerol, mono- i dwuglicerycły, kwasy tłuszczowe i powstające z nich mydła. Kwasy tłuszczowe, mono- i dwuglicerydy zostają zamienione w komórkach błony śluzowej jelita cienkiego z powrotem na trójglicerydy i przechodzą do naczyń limfatycznych i do limfy. W limfie znajdujemy tylko tłuszcze obojętne. Krótkołańcuchowe kwasy tłuszczowe mogą się wchłaniać jako wolne kwasy, przechodzą one jednak nie do limfy, lecz do żyły wrotnej i do wątroby. Nadmiar tłuszczu odkłada się w ustroju jako zapasowy materiał energetyczny. Ustrój dorosłego prawidłowo odżywionego człowieka zawiera 7-10 kg 223
Lipaza trzustkowa
Lipaza trzustkowa
Lipaza trzustkowa
Trójglicerydy
Dwuglicerydy
Monoglicerydy
Glicerol
Kwasy tłuszczowe Ryc. 125. Schemat działania lipazy trzustkowej. Lipaza odszczepia od trójglicerydów jeden, dwa lub trzy kwasy tłuszczowe, w wyniku czego powstaje mieszanina kwasów tłuszczowych, mono-, dwu-, i trójglicerydów.
tłuszczu, tj. 10-18% ogólnego ciężaru ciała. Przeważająca część tłuszczu gromadzi się w postaci tkanki tłuszczowej, występującej najobficiej pod skórą, w otoczeniu narządów jamy brzusznej, w krezce. Tłuszcz w ustroju powstaje także z węglowodanów. Ogólnie można powiedzieć, że wszelkie nadwyżki kalorii pokarmowych, niezależnie od postaci, w jakiej się dostają do ustroju, są odkładane jako tłuszcz. Tłuszcz ten stanowi zapasowy materiał energetyczny, który zostaje uruchomiony, gdy wydatek energii przewyższa jej dowóz z pokarmem. Tłuszcze są spalane w ustroju do C0 2 i H20. Pierwszym etapem na tej drodze jest rozkład tłuszczów przez lipazę na kwasy tłuszczowe i glicerol. Spalanie tłuszczów nie odbywa się w t y m miejscu, gdzie są magazynowane; w t y m celu muszą być one przeniesione do innych, bardziej aktywnych tkanek, przede wszystkim do wątroby. Rozpad kwasów tłuszczowych odbywa się stopniowo i przechodzi przez szereg etapów, w których powstają coraz to mniejsze cząsteczki kwasów. Końcowym produktem rozkładu kwasów tłuszczowych jest 224
ostatecznie kwas acetylooctowy, który zostaje spalony do C02 i H20. W pewnych warunkach chorobowych we krwi gromadzą się większe ilości kwasu acetylooctowego i produktów jego rozpadu, wywołując zaburzenia równowagi kwasowo-zasadowej - kwasicę, co stanowi duże niebezpieczeństwo dla prawidłowych funkcji ustroju.
Przemiana w o d y i soli mineralnych Woda (H20) jest materiałem niezbędnym dla wszelkich procesów życiowych. Zawartość w o d y w organizmie zwierząt żywych i człowieka wynosi około 65%. Większość reakcji chemicznych, odbywających się w trakcie przemiany, przebiega w roztworach wodnych. Woda w ustroju zawsze zawiera rozpuszczone sole mineralne i inne substancje, które wytwarzają w płynach organizmu, objętych nazwą środowiska wewnętrznego, pewne stałe ciśnienie osmotyczne. Objętość wody i ilość rozpuszczonych w niej soli mineralnych musi być taka, by utrzymać ciśnienie osmotyczne na stałym poziomie. Woda przy tym jest nie tylko zwykłym rozpuszczalnikiem reagujących substancji, lecz sama bierze aktywny udział w wielu przebiegających reakcjach.
Tabela 7 Dobowy bilans w o d n y (wg Besta i Taylora)
Dowóz Pokarm stały i półpłynny Woda powstająca w procesach przemiany Płyn (woda, mleko, herbata. kawa itd.) Razem
m) 1200 300 1000
2500 ml
Straty
ml
skóra płuca z powietrzem wydychanym mocz kał
350 500 1500 150 2500 ml
Parowanie wody wydzielanej z potem przez skórę stanowi bardzo istotny czynnik w mechanizmie regulacji ciepłoty ustroju. Część wody wchodzącej w skład tkanek znajduje się w nich w postaci związanej. Człowiek ginie szybciej z braku dowozu wody niż z braku pożywienia. Jeżeli bez pobierania pokarmu stałego człowiek może utrzymać się przy życiu około 30 dni, to bez w o d y śmierć następuje już po kilku dobach. Dobowe zapotrzebowanie wody u człowieka dorosłego wynosi ok. 40 g/kg ciężaru ciała. Zapotrzebowanie wody zostaje pokryte przez wprowadzone płyny, których używamy dziennie ok. 1-1,5 I, wodę zawartą w pokarmach 15 - Anatomia i fizjologia
225
(1-1,8 I) i wodę powstającą w tkankach w procesach spalania. Dowóz wody musi uzupełniać stałą stratę wody wydalanej przez nerki (mocz), skórę (pot), płuca (para wodna) i przewód pokarmowy (wypróżnienia). Te straty w o d y są związane z przebiegiem ważnych czynności fizjologicznych ustroju. Dowóz wody do ustroju jest regulowany swoistym czuciem pragnienia. Pragnienie powstaje wskutek odruchowego pobudzenia odpowiednich ośrodków nerwowych przy pierwszych objawach zmiany ciśnienia osrnotycznego krwi. Wchłanianie wody odbywa się w jelicie cienkim i grubym. Wchłonięta do krwi woda przechodzi przez wątrobę i dostaje się do krążenia wielkiego. Prawie cała wchłonięta woda przechodzi bardzo szybko z krwi do płynów tkankowych, toteż nawet po wypiciu znacznej ilości płynu nie obserwuje się istotnego rozwodnienia krwi. Przemiana wody pozostaje w ścisłym związku z przemianą innych substancji - białek, węglowodanów, tłuszczów, przede wszystkim zaś z przemianą soli mineralnych. Niektóre jony soli nieorganicznych zatrzymują wodę w ustroju, inne - przyczyniają się do jej utraty. Sole sodowe zatrzymują wodę w tkankach, sole potasowe i wapniowe sprzyjają wydalaniu wody z ustroju. Sole mineralne, podobnie jak woda, nie dostarczają ustrojowi energii, są jednak niezbędne do normalnego przebiegu procesów życiowych. Sole mineralne wchodzą w skład kości, nadając im twardość. Stanowią one nieodłączny składnik protoplazmy komórek, jako rozpuszczalne składniki płynów ustrojowych ddgrywają dużą rolę w procesach pobudzenia mięśni i nerwów, przyczyniają się do utrzymania równowagi kwasowo-zasadowej ustroju stałego ciśnienia osmotycznego płynów ustrojowych. Zawartość poszczególnych soli mineralnych w tkankach i płynach utrojowych jest^tała, a odchylenie od tego optymalnego stężenia stanowi poważną groźbę dla życia.
Sole wapnia Wapń (Ca) jest nieodzownym składnikiem protoplazmy i płynów tkankowych. W dużych ilościach znajduje się w kościach i zębach; 99% wszystkich soli wapnia w ustroju przypada na kości. Wapń jest ściśle związany z procesami pobudliwości mięśniowej, z czynnością serca, przepuszczalnością błon komórkowych.W postaci zjonizowanej jest czynnikiem koniecznym w krzepnięciu krwi. Organizm nie może się przystosować do niedoboru wapnia, toteż konieczny jest stały jego dowóz w pokarmach. Dla utrzymania prawidłowego poziomu wapnia wystarcza doprowadzenie 0,75 g wapnia na dzień. Najlepszym źródłem wapnia jest mleko. Dzieci, kobiety ciężarne i karmiące wymagają dowozu większych ilości wapnia.
226
Sole fosforu Fosfor (P) znajduje się w ustroju nie tylko w postaci nieorganicznych fosforanów, lecz wchodzi także w skład złożonych związków organicznych. Organiczne związki fosforu odgrywają niezwykle ważną rolę w procesach życiowych. Sole sodu i potasu Sód (Na) i potas (K) występują przeważnie w postaci jonów soli dobrze rozpuszczalnych we wszystkich tkankach ustroju. Największe ilości sodu znajdują się w płynach pozakomórkowych, potas jest natomiast przede wszystkim jonem wewnątrzkomórkowym. Inne sole Siarka (S) wchodzi w skład większości białek. Żelazo (Fe) znajduje się nie tylko w krwinkach czerwonych, lecz także we wszystkich komórkach ciała. Bierze ono udział w przenoszeniu tlenu. Zapasy żelaza w ustroju są ograniczone,toteż konieczny jest jego dowóz z pokarmem. Źródłem żelaza są: chleb razowy, żółtko jaja, mięso, owoce, zielone jarzyny. Jod (J) wchodzi w skład hormonu tarczycy - t y r o k s y n y . Zapotrzebowanie ustroju na ten pierwiastek jest niezwykle małe i wynosi 0,000044 g na dobę. Miedź (Cu) występująca w ustroju jest czynnikiem biorącym udział w tworzeniu hemoglobiny, chociaż sama w jej skład nie wchodzi.
Witaminy Witaminami nazywamy organiczne składniki pokarmowe, które nie dostarczają ani energii, ani budulca dla nowo tworzących się komórek, są jednak niezbędne dla prawidłowego funkcjonowania organizmu. Witaminy są związkami o stosunkowo niewielkiej masie cząsteczkowej i różnorodnej budowie chemicznej. W porównaniu z pbdstawowymi składnikami pokarmowymi białkami, tłuszczami i węglowodanami ilość witamin potrzebna do prawidłowego przebiegu procesów życiowych jest znikoma. Głównym źródłem witamin w pokarmie człowieka i zwierząt są rośliny, w których witaminy są syntetyzowane. Człowiek otrzymuje witaminy bądź bezpośrednio z pokarmem roślinnym, bądźz pokarmem pochodzenia zwierzęcego, w którym są witaminy nagromadzone uprzednio z materiału roślinnego. Brak lub niedobór witamin w pożywieniu pociąga za sobą znaczne zaburzenia w przemianie materii i energii, wywołując schorzenia zwane hipowitaminozami i awitaminozami. Badania ostatnich dziesięcioleci wykazyły, że istnieje ścisły związek pomiędzy witaminami a enzymami, które normują procesy przemiany materii i energii w ustroju. Okazało się, że szereg enzymów powstaje przez połączenie 227
różnych witamin z białkiem. W ten sposób choroba wywołana brakiem lub niedoborem witamin w pokarmach jest w swej istocie następstwem niedostatecznej aktywności odpowiedniego enzymu, który katalizuje odpowiednie ogniwo przemian chemicznych. Wszystkie znane dziś witaminy dzielimy na dwie grupy: 1) rozpuszczalne w tłuszczach i 2) rozpuszczalne w wodzie. Do pierwszej grupy należą witaminy A, D, E i K. Do drugiej zespół witamin B i witamina C.
Tabela 8 Witaminy Witamina
Działanie
Objawy niedoboru
Źródło
koenzym barwników wzrokowych
ślepota zmierzchowa, wysychanie skóry
zwierzęce: tran, pełne 5000 j.m. mleko, śmietana, sery, masło, żółtko jaja roślinne: żółte warzywa i owoce
Bi (Tiamina)
koenzym w przemianie węglowodanowej
beri-beri, nerwów
zwierzęce: mięso wiep- 0,5 mg rzowe, wątroba, produk- na 1000 ty mleczne kcal roślinne: rośliny strączkowe, orzechy, otręby, chleb razowy
B2 (Ryboflawina)
składnik enzymów zapalenie błony śluzo- zwierzęce: mleko, ser, jatkankowych wej języka, zajady ja, wątroba, serce, mięso roślinne: jarzyny liściaste zielone, pieczywo z razowej mąki
PP (Niacyna)
składnik nukleotydów DPN i TPN
Bg (Pirydoksyna)
228
zapalenie
pelagra, zapalenie skóry, biegunki, otępienie umysłowe (trzy D - dermatitis, diarrhoea, dementia)
stanowi grupy drgawki, nadmierna prostetyczne nie- pobudliwość nerwoktórych enzymów wo-mięśniowa (dekarboksylaz i aminotransferaz)
Minimalne zapotrzebowanie dzienne
1,8 mg
zwierzęce: wątroba, nerki, 10-15 mg chude mięso, drób roślinne: wyroby z mąki razowej, nie oczyszczane kasze, orzeszki ziemne zwierzęce: mięso, wątroba, nerki, w niewielkiej ilości w mleku roślinne: kiełki pszenicy, pieczywo razowe, orzeszki ziemne, kukurydza, w niewielkiej ilości zielone jarzyny
2 mg
Witamina
Działanie
Objawy niedoboru
Kwas pantotenowy
składnik mu A
koenzy-
zapalenie skóry, zapa- szeroko rozpowszechniolenie jelit, niewydol- ny w wielu pokarmach ność nadnerczy, łysie- zwierzęce: nerki, wątronie ba, jaja roślinne: większość świeżych jarzyn, wyroby z mąki razowej, orzeszki ziem ne
Biotyna
bierze udział w syntezie kwasów tłuszczowych
zapalenie skóry, zapa- szeroko rozpowszechniolenie jelit na w wielu pokarmach zwierzęce: nerki, wątroba, jaja, mleko roślinne: większość świeżych jarzyn
Kwas foliowy
udział w transpor- sprue, niedokrwistość cie reszt jednowęglowych
udział w metaboBi2 (cyjanokoba- lizmie aminokwalamina) sów pobudza erytropoezę
(Kwas askorbinowy)
D (Kalcyferol)
(Alfa-tokoferol)
bliżej nie znane; udział w syntezie kolagenu
Źródło
Minimalne zapotrzebowanie dzienne
szeroko rozpowszechniny w wielu pokarmach zwierzęce: wątroba kurcząt roślinne: liście zielonych warzyw
niedokrwistość złośliwa
zwierzęce: wątroba, nerki, mięczaki, chuda wołowina, cielęcina, baranina, drób, jaja, sery
gnilec
roślinne: owoce cytrusowe, jagody (czarna porzeczka), melony, ananasy, zielone jarzyny liściaste (papryka zielona, kapusta), pomidory, ziemniaki
zwiększa wchłanianie wapnia i fosforanów z przewodu pokarmowego
krzywica
koenzym w transporcie elektronów w układzie cytochromów
u zwierząt zaburzenia zwierzęce: wątroba, jaja czynności rozrodczych roślinne: olej z kiełków pszenicy, zielone jarzyriy liściaste, olej bawełniany, orzechy, rośliny strączkowe
500 ng
2,8 jtg
75 mg
zwierzęce: tran w niewielkich ilościach w mleku i 400 j.m. maśle, wątrobie, żółtku jaja kurzego
229
Witamina
Działanie
Objawy niedoboru
Źródło
K (Przeciwkrwotoczna)
bierze udział w syntezie protrombi ny w wątro bie
krwawienia
zwierzęce: wątroba roślinne: zielone jarzyny liściaste (kapusta, szpinak) kalafiory, soja
Minimalne zapotrzebowanie dzienne
Przemiana energii Przemiany w żywym ustroju toczą się na tych samych zasadach co w przyrodzie martwej. Do procesów życiowych w całej rozciągłości stosuje się powszechne prawo zachowania energii i materii, głoszące, że energia nie może z niczego powstawać ani też ginąć, możliwe są tylko przemiany jednego rodzaju energii w inny. Źródłem, z którego żywy ustrój czerpie entergię, są wysokocząsteczkowe składniki pokarmowe (węglowodany, tłuszcze, białka); w nich nagromadzony jest pewien zasób energii, określany jako energia chemiczna. Jest to wewnętrzna energia wiązań międzyatomowych w postaci drgań i ruchów atomów w cząsteczkach. W gruncie rzeczy jest to energia promieniowania słonecznego, zatrzymana i związana przez rośliny zielone w procesie fotosyntezy i ewentualnie wtórnie przekształcona przez organizmy zwierzęce. Głównym zadaniem badań nad przemianą jest poznanie dróg i sposobów, w jaki energia chemiczna substancji pokarmowych zostaje przekształcona i zużyta na ruchy mięśni, wydzielanie gruczołów, przewodzenie bodźców w nerwach, wzrost tkanek i inne przejawy energetyczne charakteryzujące żywy organizm. Metody pomiaru wydatkowania energii Wszystkie rodzaje energii wyzwalanej przez żywe komórki, z wyjątkiem pracy zewnętrznej, ulegają ostatecznie przemianie na ciepło, które ustrój oddaje otoczeniu. Ilość ciepła, a co za tym idzie i energii wytwarzanej przez ustrój, można oznaczyć za pomocą metody zwanej kalorymetrią. Istnieją dwa rodzaje kalorymetrii - bezpośrednia i pośrednia. Kalorymetria bezpośrednia polega na oznaczeniu całej ilości ciepła, jaką ustrój oddaje w pewnym określonym czasie. W zastosowaniu do człowieka metoda ta jest trudna do realizacji. Sam kalorymetr, w którym umieszcza się badanego, musi być duży i tak urządzony, aby badany mógł w nim spędzić 230
dłuższy okres czasu, gdyż ustrój oddaje ciepło stosunkowo wolno. Kalorymetr musi być należycie zabezpieczony przed utratą ciepła. Wszystkie te trudności zostały pokonane i udało się dokonać tą metodą p o m i a r ó w wydatkowanej przez ustrój energii. Otrzymane wyniki świadczą o t y m , że organizm człowieka podlega najzupełniej prawu zachowania energii i materii. Obliczenia uzyskane metodą kalorymetrii bezpośredniej pozwoliły na w p r o wadzenie o wiele prostszego sposobu oznaczania bilansu energetycznego człowieka, mianowicie kalorymetrii pośredniej. Ilość energii produkowanej przez żywy ustrój zależna jest od ilości spalonych białek, tłuszczów i węglowodanów. Stwierdzono, że przy utlenieniu 1 g białka wytwarza się w organizmie 4,1 kcal, 1 g tłuszczu daje 9,2 kcal, 1 g w ę g l o w o d a n ó w dostarcza 4,1 kcal ciepła.
Współczynnik
oddechowy
Współczynnikiem o d d e c h o w y m nazywamy stosunek objętościowy w y dzielonego dwutle.nku węgla do pobranego w t y m samym czasie tlenu, tj. C0 2 02 Stosunek ten jest różny przy utlenianiu w ę g l o w o d a n ó w , białek i tłuszczów, toteż oznaczanie współczynnika oddechowego pozwala nam do pewnego stopnia zorientować się, jakie substancje są w ustroju utleniane. Wyjaśnijmy to na przykładzie utleniania cukru gronowego. Reakcja chemiczna przebiega według równania: " C 6 H 12 0 6 + 60 2 -> 6C0 2 + 6H 2 0 Z równania tego widać, że ilość cząsteczek pobranego tlenu równa się ilości cząsteczek wydzielonego dwutlenku węgla, a ponieważ, zgodnie z prawem Avogadry, jednakowe ilości cząsteczek gazu w tych samych warunkach ciśnienia i temperatury zajmują jednakową objętość, to stosunek objętościowy CO, 02 czyli współczynnik oddechowy przy spalaniu glukozy oraz wszystkich innych w ę g l o w o d a n ó w , wynosi 1. Spalenie białka wymaga większej ilości tlenu i współczynnik oddechowy dla białek w y n o s i 0,8. Przy spalaniu tłuszczów stosunek C 0 2 : 0 2 = 0,71. Przy pokarmie mieszanym, takim jaki spożywamy w zwykłych warunkach, współczynnik oddechowy wynosi 0,85-0,9. Oznaczenie pobranego 0 2 i wydzielonego C0 2 wykonuje się za pomocą specjalnie do tego celu skonstruowanych przyrządów. Zużyciu 1 I tlenu przy spalaniu w ę g l o w o d a n ó w towarzyszy wydzielanie 5,047 kcal; przy spalaniu 231
białek i tłuszczów zużycie 1 I tlenu odpowiada nieco mniejszej ilości energii. A zatem różnym współczynnikom o d d e c h o w y m przy zużyciu 1 I tlenu odpowiadają różne ilości wyzwalajiącej się energii, co ilustruje tab. 9. Tabela 9 Współczynnik oddechowy Ilość energii wyzwolonej przy zużyciu 1 l tlenu (w kcal)
0,70
0,75
0,80
0,85
0,90
0,95
1,0
4,686
4,739
4,801
4,863
4,924
4,985
5,047
Oznaczanie wytwarzanej energii na przemiany gazowej
podstawie
Za pomocą specjalnych aparatów, z których badany pobiera tlen i do których wydycha dwutlenek węgla, oznaczamy ilość zużytego tlenu oraz wydzielonego dwutlenku węgla i z wartości tych obliczamy współczynnik oddechowy. Zta beli odczytujemy, jaka ilość kalorii p r z y d a n y m współczynniku odpowiada 1 I tlenu. Liczbę tę mnożymy przez ilość litrów zużytego tlenu i otrzymujemy w ten sposób ilość energii wytworzonej w kaloriach przy danym współczynniku oddechowym w określonym czasie. Przypuśćmy np., że w ciągu 10 m i n badany zużył 2012 ml 0 2 i wydziel ił 1970 ml C0 2 . Współczynnik o d d e c h o w y ^ = = 0,85 O2 io/U Równoważnik kaloryczny 1 I tlenu przy takim współczynniku znajdujemy w tabeli: równa się on po zaokrągleniu 4,9. A zatem w ciągu 10 m i n badany wytworzył 2,01 • 4,9 = 9,85 kcal. W przeliczeniu na dobę wyniesie to 11,6 • 6 • 24 = 1 4 1 8 kcal.
Przemiana
podstawowa
Ilość energii wytwarzanej przez żywy ustrój zależna jest od w a r u n k ó w , które zmieniają nasilenie procesów przemiany. W jednakowych warunkach wydatkowanie energii jest t y m mniejsze, im mniejsza jest praca wykonana przez ustrój. Ale nawet wtedy, gdy nie w y k o n u j e m y żadnej pracy, żadnych zbędnych ruchów, pewna ilość energii zostaje zużyta na podtrzymanie stałej temperatury ciała, pracę serca, ruchy oddechowe oraz procesy wegetatywne. Tę ilość energii nazywamy przemianą p o d s t a w o w ą albo spoczynkową. U zwierząt stałocieplnych znaczna część przemiany podstawowej idzie na utrzymanie temperatury ciała na poziomie wyższym niż temperatura otoczenia. Ponieważ organizm traci stale ciepło przez swą powierzchnię stykającą się z atmosferą, wielkość przemiany podstawowej jest bardziej zależna od powierzchni niż od ciężaru ciała. 232
U ludzi przemianę podstawową mierzymy na czczo, co najmniej 12 h po ostatnim posiłku i po kilkudniowej diecie bezbiałkowej. Przed rozpoczęciem badania układa się badanego w łóżku, poleca rozluźnienie wszystkich mięśni j pozostanie w możliwie zupełnym bezruchu. Po 30 min całkowitego spokoju włącza się aparat respiracyjny, służący do oznaczania w y m i a n y gazowej, na podstawie której oznacza się wartość przemiany spoczynkowej. U dobrze odżywionego zdrowego człowieka średniego wzrostu i ciężaru ciała przemiana podstawowa wynosi około 1800 kcal na dobę albo 40 kcal na 1 m 2 powierzchni ciała/h lub 1 kcal na 1 kg ciężaru ciała/h. Wartość przemiany podstawowej zależna jest od Wieku (u dzieci jest większa niż u dorosłych), od temperatury ciała (wzrost temperatury zwiększa podstawową przemianę materii), od temperatury zewnętrznej (chłód - zwiększa, gorąco - zmniejsza przemianę), od czynności gruczołów dokrewnych (hormony tarczycy, adrenalina i wyciąg z przedniego płata przysadki zwiększają podstawową przemianę materii poprzez h o r m o n tyreotropowy). Głód lub długotrwałe niedożywienie zmniejszają podstawową przemianę materii. Wartość przemiany podstawowej wyraża się zwykle w procentach powyżej lub poniżej normy. U osób zdrowych odchylenia od wartości prawidłowej wahają się w granicach 15%.
Przyrost
czynnościowy
przemiany
energii
Czynnikiem w p ł y w a j ą c y m najszybciej na wzrost przemiany energii w ustroju jest praca fizyczna - im cięższa jest praca mięśni, t y m większe jest wydatkowanie energii. Tabela 10 ilustruje wzrost przemiany dobowej przy pracy w różnych zawodach. Różne ilości energii zużywanej przez ustrój wymagają różnych ilości pokarmu. W czasie snu podstawowa przemiana obniża się o 8 - 1 0 % . Praca umysłowa nie w p ł y w a w sposób widoczny na wysokość przemiany. Bardzo znacznie zwiększa przemianę h o r m o n gruczołu tarczowego. W niedoczynności tarczycy T a b e l a 10 Zużycie energii Zawód
(w kcal na dobę)
- w procentach w stosunku do przemiany podstawowej = 1800 kcal
Urzędnik Krawiec Mechanik Stolarz Żołnierz na manewrach Żniwiarz Drwal
2556 2681 3189 3257 3960 4388 5600
142 149 177 181 220 244 311
233
przemiana obniża się poniżej normy, a w nadczynności (choroba Basedowa) wybitnie wzrasta.
Swoistodynamiczne
działanie
pokarmu
Spożywanie pokarmu w ilościach przewyższających zapotrzebowanie kaloryczne prowadzi do odkładania się w ustroju zapasów, głównie w postaci tłuszczów i niewielkiej ilości glikogenu. Stwierdzono, że pokarm podany zwierzęciu głodującemu nie tylko zostaje zużyty na odnowę tkanek i pokrycie potrzeb energetycznych, lecz także powoduje wzrost przemiany. Jeżeli np. głodującemu zwierzęciu podamy samo tylko białko w ilości równej pod względem kalorycznym wartości podstawowej przemiany materii, to produkcja ciepła będzie o 30% większa, niżby to wynikało z podanej ilości pokarmu. Węglowodany powodują podobny wzrost tylko o 4%, a tłuszcze o 6%. Ten wzrost przemiany pod w p ł y w e m spożytego pokarmu został nazwany swoistodynamicznym działaniem pokarmu albo „ t e r m i c z n y m e f e k t e m " pokarmu. Wytwarzanie tej dodatkowej ilości ciepła wskutek swoistodynamicznego działania pokarmu następuje w wątrobie. Przypuszcza się, że energia ta powstaje w procesie dezaminacji aminokwasów.
Żywienie Poznanie procesów przemiany materii i energii dało naukowe podstawy do ustalenia zasad odżywiania człowieka. Zasady żywienia stanowią dziś samo,dzielną gałąź wiedzy, niezwykle ważną w warunkach współczesnej cywilizacji, gdy naturalne środki spożycia nie są konsumowane na miejscu ich powstania, lecz wytwarzane na wielką skalę, konserwowane, przechowywane i transportowane na duże1 odległości. W naszych warunkach zagadnienie to jest szczególnie ważne wqfeec wielkiego wzrostu zakładów masowego żywienia. Problemami żywienia prócz fizjologii zajmuje się jeszcze szereg innych dyscyplin, jak dietetyka, higiena żywienia, technologia środków spożywczych. Fizjologia zajmuje się głównie d w o m a zagadnieniami związanymi z żywieniem: 1) ustaleniem zapotrzebowania organ izmu na różne składniki pokarmowe w różnych warunkach życia i pracy, 2) ustaleniem ilości tych składników w różnych pokarmach. Zapotrzebowanie kaloryczne M ó w i l i ś m y już, że w stanie zupełnego spoczynku zapotrzebowanie energetyczne ustroju wynosi około 40 kcal na 1 m 2 powierzchni ciała na godzinę, tj. ok. 1800 kcal na dobę. Przy ustalaniu liczby kalorii, jakie chcemy wprowadzić z pokarmem na pokrycie potrzeb energetycznych, musimy dodać około 15% całkowitej ilości kalorii na pokrycie swoistodynamicznego działania pokarmu, tj. 270 kcal. '234
A zatem na pokrycie potrzeb energetycznych ustroju pozostającego w spoczynku potrzeba około 2070 kcal. Zapotrzebowanie kaloryczne każdego człowieka zależne jest od charakteru jego czynności. Im większy udział w tych T a b e l a 11 Czynność Spokojne stanie Praca siedząca Wolny chód Szybki chód Praca średnio ciężka Praca ciężka Praca bardzo ciężka
kcal/h ponad podstawową przemianę 15 20-25 120 180-200 90-140 180-300 320-380
czynnościach ma praca mięśniowa, tym więcej kalorii musi ustrój otrzymać z pokarmem. Tabela 11 (wg Wintona i Byalissa) podaje przykłady zapotrzebowania kalorycznego ponad przemianę podstawową przy różnych czynnościach. Ustalenie dobowego zapotrzebowania energetycznego ilustruje poniższy przykład: 8 h snu - przemiana podstawowa 70 kcal/h 8 h pracy średnio ciężkiej (70 + 130 kcal/h) 8 h spokojnego zachowania się bez snu (70 + 25 kcal/h) Razem
560 kcal 1600 kcal 760 kcal 2920 kcal
Z powyższego wynika, że przeciętnie pracujący dorosły człowiek wymaga dowozu z pokarmem około 3000 kcal na dobę. Zapotrzebowanie kaloryczne dzieci jest mniejsze w wartościach bezwzględnych niż osób dorosłych, jednakże w stosunku do ciężaru ciała jest ono znacznie większe. Tabela 12 przedstawia dane liczbowe dotyczące zapotrzebowania na energię i najważniejsze składniki pokarmowe zależne od wieku (wg Food and Nutritional Board National Research Council). Każdy gram węglowodanów i białek dostarcza 4,1 kcal, 1 g tłuszczu - 9,2 kcal. Większość zapotrzebowania energetycznego pokrywają węglowodany. Ilość podstawowych składników pożywienia przy racji dziennej np. 3380 kcal powinna wynosić: 100 g białka 100 g tłuszczu 500 g węglowodanów
410 kcal 920 kcal 2050 kcal
'235
° 8 <2 ^
o o o o o o
o
~ u
C O C
O O o O o co o co
• i. s £ U £ o
I
2
o o o o o o oO Co oNI Co oM o o 00 o C CD ID C5
o o o
oo ho co O IO U) t ld co r^
CO CD ««- r- CM
o o o 5 o o
o o o CNI o 00 CO
oM oLO 0o0 C CM CM CM
O O LD LD 00 CO CO CO
o ifl o io o ^r
o 00 o 00 o
CO B S
CD O CM CM (M
lo r^ ld
o o rt I-
O— — * C— *O Cv— * CD*—<-CMo CNI
> O) .2 E
0) 'Z o <0
o o o o o o o o o o o o o o
(35 CO CM «coCM 00 o Mct>i co - CM CM CM CM CNI C
2 2 °
o o o
o o o o o CD oco o o
o o o o coo o 00 o
o o o 0o o «-0 CM
o o o o o 00 LoD 00
o o o o o Ol o NO
o o 00
o o O
CM
o o
o o o o o o o o CO ^J LD
o o o o o o o o o o o o LD CD 00 O CM t
o o o o OJ o O 0o5
O o o LO
o o o ID
o o o co
o o o oo
o o o o o o O o O o ID ID — ł CM CM
o o o o o oO o o o o o o LDLD O O LD LD •tf LD CM CM CO
o o o o o o LD O O
LO ID O
O CM
o CM
CO CD
o o 00
o o 00
o o
o
00
® m
Ao - lt
C3 ®
EE co < ~'
co O) ®E
« o t! <2 3 a a o < no
o ® S?>0-0 e 5 tn c o ® Nm £ ">O
E ^ o Z
ra a)
m
o o
CO
CO
00
LO
Ci
^ 11) ID
05
o o
o o 00
o o o o o o
o o o o oO 0 o0 «- C
C CM r-Mł-h-CMOCM
CM O CO
O LD LD CD t^ 00
LLD DC CM D L00 D
p^ a) co ld" pC oo"
00 00
CO LD — t CM CO CD
co CO r^ Lco D
c>-
o O lo lo co id lo r--
I I inI a i-ooifl o lo
03 O.
3 O
'236
ID ID ID LD ID CO LD r^ CO LO CO ID
o > ot <-Ł J> !>) Jli) i-
m
oi »-
co a) fl) © ctr i
Sr ••N 92 la Ci A o
CO I CD I O)I
i- co co
m
CO CO U) U) E '£ E E
00 CO O) CM
I CM I in i. C) CM LD O!
^ -N g\9>
i i i T o r^ o
o
•C
o
Tabela
13
Normy zapotrzebowania dziennego mężczyzn na białko, tłuszcze, węglowodany i kalorie zależnie od wieku i ciężaru ciała. Wiek
Ciężar należny
Białko
Tłuszcze
Węglowodany
Kalorie
lat
kg
9
9
9
(kcal)
50 55 60 65 70 75 80 85
50 55 60 65 70 75 80 85
62 66 70 74 78 82 86 90
370 392 415 438 460 485 505 528
2300 2450 2600 2750 2900 3050 3200 3350
50 55 60 65 70 75 80 85
50 55 60 65 70 75 80 85
55 59 63 67 70 74 78 82
325 348 370 393 405 428 450 473
2050 2200 2350 2500 2600 2750 2900 3050
50 55 60 65 70 75 80 85
50 55 60 65 70 75 80 85
47 50 52 56 59 62 66 69
270 283 298 320 333 345 368 380
1750 1850 1950 2100 2200 2300 2450 2550
18-35
35-55
55-75
T a b e l a 14 Normy zapotrzebowania dziennego kobiet na białko, tłuszcze, węglowodany i kalorie zależnie od wieku i ciężaru ciała Wiek lat
18-35
35-55
Ciężar należny kg
Białko g
Tłuszcze g
Węglowodany g
Kalorie (kcal)
40 45 50
40 45 50
55 60 65 70
55 60 65 70
43 47 51 54 58 62 64
253 275 298 310 333 355 370
1600 1750 1900 2000 2150 2300 2400
40 45
40 45
39 43 46 48
225 248 260 275
1450 1600 1700 1800
5Q
5Q
55
55
'237
Wiek lat
Ciężar należny kg
Białko
Tłuszcze
Węglowodany
g
g
g
Kalorie (kcal)
35-55
60 65 70
60 65 70
52 55 59
298 310 333
1950 2050 2200
40 45 50 55 60 65 70
40 45 50 55 60 65 70
32 35 39 42 44 47 50
180 192 215 228 243 255 268
1200 1300 1450 1550 1650 1750 1850
55-75
Regulacja temperatury Człowiek, podobnie jak zwierzęta stałocieplne, ma zdolność utrzymywania temperatury swego ciała na stałympoziomie, dużo wyższym i niezależnym od otoczenia. U człowieka temperatura ciała mierzona pod pachę waha się w granicach 36,5-36,9°C. Temperatura mierzona w odbytnicy jest o 0,4-0,5° wyższa. Temperatura powierzchni skóry jest niższa: przy 37° pod pachą temperatura skóry twarzy wynosi 20-26°, kończyn 20°, skóry brzucha 34°. Temperatura narządów wewnętrznych jest wyższa, temperatura wątroby wynosi 38-38,6°. Temperatura ciała ulega wahaniom dobowym: najniższa jest wczesnym rankiem, najwyższa późnym popołudniem. U kobiety występują rytmiczne wahania temperatury w związku z cyklem miesiączkowym - w okresie miesiączkowym i bezpośrednio po nim temperatura jest niższa. Bilans cieplny ustroju Jak już wspomnieliśmy, energia cieplna ustroju powstaje nieustannie wskutek utleniania składników pokarmowych. Ilóść ciepła produkowanego przez człowieka wynosi przeciętnie 2400-2800 kcal na dobę, zaś w warunkach zupełnego spoczynku wartość ta wynosi ok. 1800 kcal. Ciepło to jest oddawane otoczeniu głównie przez powierzchnię ciała, w mniejszym stopniu z wydychanym powietrzem i wydzielinami. Jest rzeczą zrozumiałą, że dla utrzymania stałej temperatury ciała musi istnieć równowaga między ilością ciepła produkowanego w ustroju i oddawanego do otoczenia. Ciepło w ciele człowieka nie jest zatrzymywane, gdyż spowodowałoby to stały wzrost temperatury ciała, prowadzący do przegrzania ustroju. Strata ciepła przez ustrój zależna jest głównie od ciągle zmieniających się warunków klimatycznych środowiska zewnętrznego. Mechanizmy regulacyjne stojące na straży utrzymania stałej temperatury ciała polegają z jednej strony na regulowaniu produkcji ciepła, z drugiej na regulowaniu ilości oddawanego ciepła. '238
Produkcja ciepła Ciepło wytwarzane jest w ustroju we wszystkich tkankach, w których odbywa się utlenianie, przede wszystkim zaś w mięśniach i w wątrobie. Produkcja ciepła w wątrobie i w mięśniu sercowym jest mniej więcej stała, natomiast w mięśniach szkieletowych zmienia się w bardzo szerokich granicach - j e s t duża w czasie pracy i bardzo mała w stanie spoczynku. W stanie spoczynku w mięśniach powstaje t y m więcej ciepła, im większe jest ich napięcie (tonus), utrzymywane przez impulsy nerwowe, nieustannie dochodzące do nich z mózgu. Produkcja ciepła w pozycji leżącej przy świadomie napiętych mięśniach jest o 10% wyższa niż przy mięśniach rozluźnionych. Nieznaczne ruchy powodują wzrost produkcji ciepła o 25%, chodzenie o 60-80%, a ciężka praca do 500%. W wyniku ochłodzenia powierzchni ciała powstaje na drodze odruchowej m i m o w o l n y , okresowy skurcz mięśni w postaci „dreszczu", wskutek czego zwiększa się produkcja ciepła. Udział mięśni w wytwarzaniu ciepła można wykazać doświadczalnie na zwierzętach zatruwając je kurarą. Kurara poraża mięśnie poprzecznie prążkowane, czemu towarzyszy znaczne obniżenie temperatury ciała.
Straty ciepła Wspomnieliśmy wyżej, że wytwarzane w ustroju ciepło nie może się w nim zatrzymywać, gdyż doprowadziłoby to do przegrzania ciała i podniesienia jego temperatury do takiego poziomu, przy którym przebieg procesów życiowych jest niemożliwy. Oddawanie ciepła jest zatem czynnikiem niezbędnym do utrzymania stałej temperatury ciała nawet przy normalnym, a t y m bardziej przy zwiększonym wytwarzaniu ciepła. Natomiast przy normalnej temperaturze wewnętrznej zaczynają działać mechanizmy ochronne zabezpieczające ustrój przed nadmierną utratą ciepła. Główną drogą, jaką organizm traci ciepło, jest powierzchnia ciała. Przez skórę człowiek traci około 4 / 5 całej ilości ciepła. Procentowy stosunek ilości ciepła oddawanego przez organizm człowieka różnymi drogami podaje tab. 15. Oddawanie ciepła przez powierzchnię ciała odbywa się w trojaki sposób: 1) przez promieniowanie, 2) przez przewodzenie ciepła i 3) przez parowanie potu. Ilość ciepła oddawanego w każdym z tych sposobów zależna jest od T a b e l a 15 Straty ciepła Przez powierzchnię skóry Przez płuca Na ogrzanie spożywanego pokarmu i w o d y Wydalanego z moczem i kałem
% 82,0 13,0 4,6 1,3
'239
w a r u n k ó w otoczenia zewnętrznego. Gdy temperatura skóry jest wyższa od temperatury otoczenia, utrata ciepła odbywa się przez promieniowanie i przewodzenie. Ogrzana powierzchnia ciała oddaje energię cieplną w postaci promieni podczerwonych, które można odczuć zbliżając do siebie dłonie na odległość kilku centymetrów. Odczuwa się wówczas ciepło j a k o d nagrzanego przedmiotu. Utrata ciepła przez przewodnictwo polega na t y m , że bardziej ogrzane ciało oddaje ciepło cząsteczkom otaczającego go środowiska. Ilość ciepła oddawana tą drogą zależna jest od różnicy temperatur między ciałem a otoczeniem, od ciepła własnego środowiska stykającego się z ciałem oraz od ruchu cząsteczek ogrzanych (konwekcji). Przy temperaturze otoczenia wynoszącej 15° człowiek normalnie ubrany traci 44% oddawanej ilości ciepła przez promieniowanie, a 30% przez przewodzenie i konwekcję. Regulacja oddawania ciepła przez promieniowanie i przewodzenie polega na zmianach ukrwienia skóry. Gdy temperatura środowiska zewnętrznego jest wysoka, naczynia krwionośne skóry rozszerzają się, naczynia zaś narządów wewnętrznych zwężają się. W ten sposób krew przenosi ciepło z miejsca jego powstania do skóry, gdzie następuje jego utrata. I na odwrót, przy niskiej temperaturze otoczenia naczynia skóry ulegają zwężeniu, dzięki czemu utrata ciepła zostaje ograniczona. Utrata ciepła przez parowanie wysuwa się na pierwszy plan wówczas, gdy temperatura otoczenia dorównuje lub przewyższa temperaturę ciała. Przejście w o d y w parę w y m a g a energii cieplnej dla pokonania wzajemnego przyciągania cząsteczek (ciepło parowania). Strata ciepła przy parowaniu potu ze skóry jest znaczna - wyparowanie z powierzchni skóry 1 g w o d y w temperaturze 30° odbiera około 0,58 kcal. Ilość ciepła oddawana przez parowanie jest zależna od ilości potu wytwarzanego przez gruczoły potowe. W w a r u n k a c h normalnych odbywa się przez skórę parowanie niewyczuwalne (perspiratio insensibilis), przy wzroście temperatury ciała lub wysokiej temperaturze otoczenia wydzielanie potu zwiększa się bardzo znacznie. W warunkach tropikalnych lub przy bardzo ciężkiej pracy w wysokiej temperaturze (górnicy, hutnicy) ilość wydzielanego potu może dochodzić do 10-121 w ciągu dnia. Parowanie w o d y z powierzchni ciała zależne jest od temperatury, wilgotności i ruchu otaczającego powietrza. Im mniejsza jest wilgotność otaczającego powietrza, t y m większe jest parowanie, dlatego też w zakładach pracy, gdzie panuje wysoka temperatura i utrata ciepła jest możliwa tylko przez parowanie, wielkie znaczenie ma kontrola wilgotności powietrza. Stosunkowo niewielka ilość ciepła jest oddawana z w y d y c h a n y m powietrzem i wydalinami (mocz i kał). Regulacja temperatury ciała Wytwarzanie i utrata ciepła zależne są od: 1) nasilenia procesów przemiany, 2) zmian w ukrwieniu i 3) wydzielania potu. Wszystkie te czynniki pozostają pod kontrolą układu nerwowego i gruczołów dokrewnych. W ośrodkowym '240
układzie nerwowym znajduje się specjalny ośrodek termoregulacji, położony w podwzgórzu, który wywiera wpływ na temperaturę ciała za pośrednictwem ośrodka naczynioruchowego, nerwów idących do gruczołów potowych, nadnerczy, wątroby i mięśni szkieletowych. Nakłucie igłą tej okolicy podwzgórza powoduje u zwierząt wzrost temperatury ciała. W warunkach fizjologicznych ośrodek termoregulacji może być pobudzony albo na drodze odruchowej przez bodźce dochodzące z powierzchni skóry, albo też przez zmiany temperatury krwi, dopływającej do tego ośrodka. Pobudzenie z ośrodka termoregulacji zostaje przekazane za pośrednictwem układu autonomicznego do narządów biorących udział w wytwarzaniu bądź utracie ciepła.
UKŁAD O D D E C H O W Y
Dla procesów życiowych organizmu konieczny jest tlen, który pobieramy z powietrza w procesie oddychania. Ustroje jednokomórkowe oddychają całym ciałem. W miarę różnicowania się układów w przebiegu procesu filogenetycznego wykształcają się narządy oddechowe. U stawonogów są nimi cienkie, rozgałęzione rureczki, zwane tchawkami. Ryby pobierają tlen rozpuszczony w wodzie za pomocą narządów zwanych skrzelami. Są to fałdy
Ryc. 126. Schemat układu oddechowego.
'242
bogato unaczynionej tkanki, między którymi przepływa woda zawierajęca tlen. U ssaków i u człowieka narządy, które biorą udział w procesie oddychania, tworzą układ oddechowy (apparatus respiratorius). W s k ł a d układu oddechowego wchodzą górne i dolne drogi oddechowe oraz właściwy narząd oddechowy - płuca. Do dróg oddechowych górnych zaliczamy: jamę nosową i gardło, będące jednocześnie składnikiem dróg pokarmowych. Do dróg oddechowych dolnych należą: krtań, tchawica i oskrzela. Drogi oddechowe kończą się w płucach, właściwych narządach oddechowych, w których odbywa się wymiana gazowa.
DROGI ODDECHOWE
l\los i jama nosowa Nos (nasus) ma kształt trójściennej piramidy, podstawą zwróconej ku dołowi. Wierzchołek piramidy tworzy nasada nosa. Leży ona między łukami b r w i o w y m i , oddzielona od gładzizny zagłębieniem. Powierzchnie boczne nosa mają kształt trójkątny; stykając się ze sobą, tworzą one grzbiet nosa. W części górnej powierzchnie te są równe, w części dolnej są uwypuklone i stanowią skrzydła nosa. Rusztowanie nosa tworzą kości nosowe i chrząstki. Część górna nosa jest kostna, część dolna ma szkielet chrząstkowy. Poniżej kości nosowych grzbiet nosa stanowi chrząstka przegrody nosa. Po obu jej stronach znajdują się chrząstki boczne. Górny brzeg każdej z nich łączy się z w o l n y m brzegiem kości nosowej, zaś brzeg przyśrodkowy - z chrząstką przegrody nosa. W skrzydłach nosa leżą chrząstki skrzydłowe. W ustawionej poziomo podstawie nosa znajdują się otwory, zwane nozdrzami przednimi, które prowadzą do j a m y nosowej. Nos pokryty jest skórą, w której znajdują się liczne gruczoły łojowe. Poza człowiekiem nos występuje u niektórych małp człekokształtnych, ale jest on słabo wyrażony. Ze względu na kształt profilu rozróżniamy nos prosty (grecki), nos w y p u k ł y (rzymski), nos haczykowaty (semicki) i nos wklęsły (zadarty). Kształt nozdrzy przednich zależy od ksżtałtu nosa. Nos wąski ma nozdrza ustawione strzałkowo, w nosach szerokich kierunek długiej osi nozdrzy zbliżony jest do poprzecznego. Kształty nosa są dziedziczone przez potomstwo. Charakterystyka kształtu nosa ma bardzo ważne znaczenie w badaniach antropologicznych. Jama nosowa (cavum nasi) podzielona jest na dwie połowy przegrodą nosa. Przegroda ustawiona w płaszczyźnie strzałkowej zbudowana jestz kości, chrząstki i skóry. W skład części kostnej przegrody nosa wchodzi lemiesz i blaszka pionowa kości sitowej (p. Układ kostny), część chrzęstną tworzy chrząstka przegrody nosa. Najbardziej ku przodowi wysunięta część przegro'243
dy zbudowana jest ze skóry i nosi nazwę części błoniastej. Od tyłu jama nosowa łączy się z gardłem przez nozdrza tylne. Każda z połówek jamy nosowej podzielona jest dodatkowo przez trzy małżowiny nosowe na trzy przewody nosowe. Przewód nosowy górny leży między małżowiną nosową górną a środkową, środkowy znajduje się między małżowinami nosową środkową a dolną i przewód nosowy dolny leży między małżowiną nosową dolną a dnem jamy nosowej. Między wolnymi brzegami małżowin a przegrodą nosa znajduje się przewód nosowy wspólny. Ku przodowi od przewodów nosowych leży przedsionek nosa. Tworzy go rozszerzona część jamy nosowej między nozdrzami przednimi a fałdem błony śluzowej, zwanym groblą nosa. Grobla nosa biegnie od końca przedniego małżowiny nosowej środkowej ku przodowi. Od tyłu przewody nosowe łączą się z przewodem nosowo-gardłowym, otwierającym się przez nozdrza tylne do części nosowej gardła. Jama nosowa łączy się z wypełnionymi powietrzem zatokami przynosowymi. Należą do nich: zatoka klinowa, zatoka szczękowa, zatoka czołowa oraz komórki kości sitowej (p. Układ kostny). Z przewodem nosowym górnym łączy się zatoka klinowa i komórki sitowe tylne. Przewód nosowy środkowy łączy się z zatoką klinową, zatoką szczękową i komórkami sitowymi przednimi. Przewód nosowy dolny nie łączy się z zatokami przynosowymi, lecz ma połączenie z oczodołem przez przewód nosowo-łzowy. Przez ten przewód ciecz łzowa odpływa z worka łzowego do jamy nosowej. Jama nosowa wysłana jest błoną śluzową, pokrytą nabłonkiem wielowarst w o w y m migawkowym; jedynie przedsionek jamy nosowej pokrywa skóra z tkwiącymi w niej krótkimi, grubymi włosami. Błona śluzowa jest silnie unaczyniona i zawiera liczne komórki wydzielające śluz. Zatoki przynosowe wyścielone są błoną podobną do błony śluzowej jamy nosowej. Błona śluzowa jamy nosowej jest obficie unaczyniona. Najliczniejsze rozgałęzienia naczyń obserwujemy w błonie śluzowej przegrody nosa i stąd najczęściej powstają krwawienia. Nerwy błony śluzowej nosa należą do I i II gałęzi nerwu trójdzielnego. Ponadto część węchowa zaopatrywana jest przez włókna węchowe dochodzące do nerwów węchowych. Czynnościowo jamę nosową można podzielić na: 1) okolicę węchową i 2) okolicę oddechową. Okolica węchowa zajmuje górną część jamy nosowej, leżącą między małżowiną nosową górną a przegrodą nosa. W błonie śluzowej okolicy węchowej znajdują się zakończenia nerwów węchowych (p. Narządy zmysłów). Okolica oddechowa obejmuje pozostałą część jamy nosowej. Przez okolicę oddechowa przechodzi powietrze do dalszych odcinków dróg oddechowych. Powietrze wdychane w jamie nosowej zostaje: a) oczyszczone z kurzu, który osiada na wilgotnej błonie śluzowej, b) ogrzane, c) nasycone parą wodną. Dzięki temu powietrze wdychane do płuc nie zanieczyszcza, nie oziębia i nie wysusza dolnych dróg oddechowych i samych płuc. '244
Krtań Droga oddechowa prowadzi z jamy nosowej do j a m y gardła, gdzie krzyżuje się z drogą pokarmową. Jamę gardła opisano w rozdziale,o przewodzie pokarmowym. Z j a m y gardła droga oddechowa prowadzi do krtani. Krtań (larynx) ma kształt trójściennej piramidy, szerszej u góry, zwężającej się ku dołowi. Zawieszona za pomocą więzadeł i mięśni na kości gnykowej leży na wysokości IV i V kręgu szyjnego. Połączenie z kością gnykową sprawia, że ruchy jej są przekazywane krtani, tak że mięśnie unoszące kość gnykową unoszą jednocześnie krtań. Od dołu krtań łączy się z tchawicą. Do powierzchni bocznych krtani przylega gruczoł tarczowy (tarczyca) i częściowo mięśnie szyi. Tylna powierzchnia krtani sąsiaduje z gardłem. Krtań jest jednocześnie narządem głosu i wskutek wysoce wyspecjalizowanych czynności jej budowa jest bardzo złożona.
Ryc. 127. Krtań widziana od strony lewej i nieco od przodu.
'245
Chrząstki krtani Szkielet krtani tworzy 9 chrząstek, które są połączone za pomocą więzadeł i mięśni. Chrząstki dzielą się na nieparzyste i parzyste (tab. 16). Chrząstka tarczowata składa się z dwóch czworokątnych płytek chrzęstnych, złączonych pód kątem skierowanym wierzchołkiem ku przodowi. Kąt ten jest większy u dzieci i u kobiet (wynosi zwykle powyżej 120°), u mężczyzn zbliża się do kąta prostego (90°). W części górnej połączenia widnieje głębokie wcięcie tarczowe górne, w części dolnej - s ł a b o zaznaczone wcięcie tarczowe dolne. Tylne krawędzie obu płytek przechodzą w rogi górne, ku dołowi w rogi dolne. Rogi dolne łączą się stawami z chrząstką pierścieniowatą. Chrząstka tarczowata daje się łatwo wyczuć przez skórę na przedniej powierzchni szyi. Niejednokrotnie, szczególnie u mężczyzn, można zauważyć wyniośłość krtaniową, zwaną także „jabłkiem Adama". Tworzą ją wywinięte brzegi wcięcia tarczowego górnego. 0 T a b e l a 16 Chrząstki krtani Chrząstki nieparzyste Chrząstka tarczowa
(cartilago
thyroidea)
Chrząstka pierścieniowata
(cartilago
cricoidea)
Chrząstka nagłośniowa
(cartilago
epiglottica)
Chrząstki parzyste chrząstka nalewkowata
(cartilago
arytenoidea)
chrząstka różkowata
(cartilago
corniculata)
chrząstka klinowata
(cartilago
cuneiformis)
Chrząstka pierścieniowata leży poniżej chrząstki tarczowej i ma kształt sygnetu. Składa się ze skierowanego ku przodowi łuku i leżącej od tyłu płytki. Na płytce, w miejscu przejścia jej brzegu górnego w boczny, leżą powierzchnie stawowe, służące do połączenia z chrząstkami nalewkowatymi. Na bocznych powierzchniach łuku chrząstki pierścieniówatej znajdują się powierzchnie stawowe dla rogów dolnych chrząstki tarczowej. Chrząstka nagłośniowa ma kształt zbliżony do liścia. Jej węższa część szypułka - łączy się za pomocą więzadła z okolicą wcięcia tarczowego górnego. Inne więzadła łączą nagłośnię z podstawą języka. Chrząstka nagłośniowa ma dużą sprężystość i pochylając się ku tyłowi zamyka wejście do krtani. Chrząstka nalewkowata, parzysta, ma kształt zbliżony do ostrosłupa trójściennego, o podstawie trójkątnej. Kąt podstawy, zwrócony ku przodowi, tworzy wyrostek głosowy, kąt skierowany ku bokowi - wyrostek mięśniowy. Na dolnej powierzchni trzeciego tylnego kąta widnieje powierzchnia stawowa łącząca się po obu stronach z powierzchniami stawowymi płytki chrząstki pierścieniówatej. Wierzchołek chrząstki nalewkowatej skierowany ku górze łączy się z chrząstką różkowatą. '246
Parzysta chrząstka różkowata jest bardzo mała, ma kształt stożka i łączy się z wierzchołkiem chrząstki nalewkowatej. Chrząstka klinowata, parzysta, mała, o zmiennym kształcie, tkwi w tylnej części fałdu nalewkowo-nagłośniowego. Chrząstki tarczowata, pierścieniowata oraz nalewkowate są połączone ze sobą stawami, w których zachodzą ruchy w ściśle określonych kierunkach. Chrząstka tarczowata wykonuje ruchy w płaszczyźnie strzałkowej, oddalając się przy t y m lub zbliżając do chrząstek nalewkowatych. Obraca się przy t y m dookoła osi poprzecznej, biegnącej przez stawy pierścienno-tarczowe. Chrząstki nalewkowate wykonują ruchy obrotowe dookoła osi pionowej, biegnącej od wierzchołka tej chrząstki do jej podstawy. Stawy między chrząstkami krtani zbudowane są podobnie do innych stawów, ruchy jednak są w nich ograniczone.
Więzadła krtani Więzadła krtani dzielą się na dwie grupy (tab. 17). Więzadło t a r c z o w o - g n y k o w e pośrodkowe i oba więzadła tarczowo-gnykowe boczne łączą chrząstkę tarczowatą z kością gnykową. Przestrzeń między tymi więzadłami zajmuje szeroka i cienka błona tarczowo-gnykowa. Więzadło pierścienno-tchawicze wypełnia przestrzeń między chrząstką pierścieniowatą a pierwszą chrząstką tchawicy, tworząc połączenie między t y m i narządami. Więzadło głosowe (ligamentum vocale), zwane teżwięzadłem tarczowo-nal e w k o w y m dolnym, biegnie od wyrostka głosowego chrząstki nalewkowatej do wewnętrznej powierzchni chrząstki tarczowatej w miejsce połączenia obu jej płytek; drgania tego więzadła powodują powstawanie głosu. Więzadło kieszonki krtaniowej przebiega nieco wyżej ku bokowi i równolegle do więzadła poprzedniego. Między tymi d w o m a więzadłami leży wejście do kieszonki krtaniowej. Więzadło pierścienno-tarczowe w postaci szerokiego, łącznotkankowego pasma łączy chrząstkę pierścieniowatą z chrząstką tarczowatą. Więzadło tarczowo-nagłośniowe łączy szypułę chrząstki nagłośniowej z górnym wcięciem chrząstki tarczowej. Tabela
17
Więzadła krtani Więzadła łączące krtań z otoczeniem
Więzadła właściwe krtani
Więzadło tarczowo-gnykowe pośrodkowe
Więzadło kieszonki krtaniowej
Więzadła tarczowo-gnykowe boczne
Więzadło pierścienno-tarczowe
Więzadło pierścienno-tchawicze
Więzadło tarczowo-nagłośniowe
Więzadło głosowe
'247
Mięśnie krtani Mięśnie krtani, podobnie jak więzadła, dzielą się na dwie grupy: mięśnie przebiegające między krtanią, kością gnykową i mostkiem oraz mięśnie właściwe krtani. Pierwsze działają na całą krtań i mogą ją unosić lub obniżać. Dokładniej zostały one omówione w rozdziałach o mięśniach szyi; Mięśnie właściwe krtani wpływają ha ruchy poszczególnych chrząstek, zmieniając ich wzajemne położenie. Jedne z nich zbliżają lub oddalają chrząstkę tarczowatą od chrząstek nalewkowatych, inne zwężają szparę głośni, jeszcze inne wywołują jej rozszerzenie. Mięśnie właściwe krtani są podane w t a b . 18. Mięsień pierścienno-tarczowy jest jedynym mięśniem zewnętrznym krtani. Rozpoczyna się na łuku chrząstki pierścieniowatej, kończy na powierzchni Chrząstka tarczowata Więzadło głosowe
#I ll-
#
i Wi \ i
"'•U 0
iiNe A
B
Chrząstka nalewkowata
w
C
Ryc. 128. Schemat położenia więzadeł g ł o s o w y c h : A- głośnia zamknięta, B - g ł o ś n i a częściowo otwarta, C - g ł o ś n i a otwarta maksymalnie. Tabela
18
Mięśnie w ł a ś c i w e krtani Czynność
Mięsień
zbliżanie chrząstki tarczowatej
Mięsień g ł o s o w y (m. vocalis)
do nalewkowatych
Mięsień tarczowo-nalewkowy (m.
thyroarytenoideus)
Mięsień pierścienno-tarczowaty (m.
Mięsień pierścienno-nalewkowy t y l n y
'248
od nalewkowatych
cricothyroideus)
Mięsień nalewkowy poprzeczny (m. artytenoideus transversus) Mięsień nalewkowy skośny (m. artytenoideus obliguus) Mięsień pierścienno-nalewkowy boczny (m. cricoarytenoideus lateralis)
(m.
oddalanie chrząstki tarczowatej
cricoarytenoideus
posterior)
zwężanie głośni
rozszerzanie szpary głośni
bocznej chrząstki tarczowatej, przebiegając skośnie od dołu, przodu i środka ku górze, bokowi i tyłowi. Zbliża on łuk chrząstki pierścieniówatej do chrząstki tarczowatej, oddalając od siebie chrząstki nalewkowatą i tarczowatą..Pociąga to za sobą napięcie fałdów głosowych. Mięsień pierścienno-nalewkowy boczny przyczepia się do górnej części bocznej powierzchni łuku chrząstki pierścieniówatej, kończy na wyrostku mięśniowym chrząstki nalewkowatej. Pociąga ten wyrostek ku przodowi, przy czym wyrostek głosowy przesuwa się w kierunku przyśrodkowym. W ten sposób następuje zwężenie głośni. Mięsień pierścienno-nalewkowy tylny rozpoczyna się na tylnej powierzchni płytki chrząstki pierścieniówatej, kończy na wyrostku mięśniowym chrząstki nalewkowatej. Pociągając ten wyrostek ku tyłowi, powoduje oddalenie od siebie wyrostków głosowych i w ten sposób rozszerza głośnię. Mięsień g ł o s o w y przebiega równolegle do więzadła głosowego, przylegając do niego od strony bocznej. Tworzy on wspólnie z więzadłem g ł o s o w y m podstawę fałdu głosowego. Mięsień t a r c z o w o - n a l e w k o w y ma takie same przyczepy, jak więzadło kieszonki krtaniowej. Obydwa powyższe mięśnie zbliżają chrząstkę tarczowatą do chrząstek nalewkowatych. Wraz z mięśniem pierścienno-tarczowym, działającym odwrotnie, regulują napięcie fałdów głosowych, a co za t y m idzie wysokość głosu. Mięsień n a l e w k o w y poprzeczny łączy obydwie chrząstki nalewkowate, przyczepiając się do ich tylnej powierzchni. Mięsień kurcząc się zbliża do siebie chrząstki nalewkowate, przez co zwęża szparę głośni. Mięsień n a l e w k o w y skośny przebiega ukośnie między jedną a drugą chrząstką nalewkowatą i podobnie jak poprzedni zwęża szparę głośni.
Jama krtani Jama krtani (cavum laryngis) wysłana jest błoną śluzową, pokrytą nabłonkiem wielorzędowym m i g a w k o w y m . Wyjątek stanowią fałdy głosowe, pokryte nabłonkiem w i e l o w a r s t w o w y m płaskim. Migawki nabłonka poruszają się w kierunku iamy gardła i wyrzucają drobne ciałka, trafiające z powietrza (kurz, sadze itp.). Wnętrze krtani dzielimy na trzy odcinki: 1) przedsionek krtani, 2) głośnię i 3) jamę podgłośniową. Przedsionek krtani (vestibulum laryngis) rozpoczyna się wejściem do krtani ograniczonym od przodu krawędzią nagłośni, z boku fałdami nalewkowo-nagłośniowymi. Każdy z nich przebiega od bocznej krawędzi nagłośni do wierzchołka chrząstki nalewkowatej. W tylnej części fałdu widoczne są dwie wyniosłości. Boczną tworzy guzek klinowaty, który odpowiada chrząstce klinowatej. Przyśrodkowa, zwana guzkiem różkowatym, jest utworzona przez leżącą w niej chrząstkę o tej samej nazwie. '249
Kieszonka krtaniowa
Jama podgłośniowa
Ryc. 129. Przekrój czołowy przez krtań widzianą od tyłu.
Głośnia ( g l o t t i s ) jest najwęższą częścią jamy krtani. Od góry ograniczają ją fałdy kieszonki krtaniowej. Każda z nich biegnie od wewnętrznej powierzchni chrząstki tarczowatej do chrząstki nalewkowatej. Pod błoną śluzową znajduje się w t y m miejscu znane nam już więzadło kieszonki krtaniowej. Poniżej fałdów kieszonki krtaniowej przebiegają fałdy głosowe. We wnętrzu tych fałdów znajduje się więzadło głosowe oraz przebiegający bocznie mięsień głosowy. Pomiędzy fałdem kieszonki krtaniowej i fałdem głosowym widzimy zagłębienie, które bocznie od fałdu kieszonki krtaniowej uwypukla się ku górze, tworząc przestrzeń, zwaną kieszonką krtaniową. Tylne końce fałdów głosowych mogą się zbliżać lub oddalać od siebie, co zależy od położenia chrząstki nalewkowatej. Część szpary głośni zawarta między fałdami głosowymi nosi nazwę części międzybłoniastej. Ku t y ł o w i między wyrostkami chrząstki nalewkowatej znajduje się część międzychrząstkowa szpary głośni. Szerokość głośni, jak i napięcie fałdów głosowych zależą od działania mięśni krtani. Powietrze przechodząc przez zwężoną głośnię wprawia w drganie fałdy głosowe. Wysokość głosu, który przy t y m powstaje, zależy m. in. od napięcia fałdów głosowych, natomiast siła głosu zależy od szybkości prądu powietrza przechodzącego przez głośnię. Wysokość głosu zależy również od b u d o w y krtani. Na ogół u osób o małej krtani głos jest wyższy. Mężczyźni mają głos niższy z powodu większej krtani i co za t y m idzie - dłuższych fałdów głosowych. '250
Barwa głosu, po której możemy poznać mówiącego, zależy od b u d o w y i funkcji przestrzeni rezonansowych. Są nimi: jama gardłowa, jama nosowa oraz zatoki przynosowe. Inną czynnością krtani jest ochrona dróg oddechowych przed wtargnięciem doń pokarmu z jamy gardła. Rolę tę spełnia przede wszystkim nagłośnia, a następnie mięśnie zamykające szparę głośni. Wejście do krtani, szparę głośni, fałdy kieszonki i fałdy głosowe możemy obejrzeć u człowieka żywego posługując się lusterkiem laryngologicznym, w p r o w a d z o n y m do jamy gardła lub też specjalnym przyrządem, zwanym laryngoskopem. Badanie laryngoskopowe krtani ma bardzo ważne znaczenie diagnostyczne w schorzeniach tego narządu. Poniżej fałdów głosowych światło krtani ponownie się rozszerza, tworząc jamę podgłośniową ( cavum infraglotticum), która przechodzi bezpośredniow tchawicę. Jak w s p o m n i a n o , krtań z tchawicą łączy się za pośrednictwem więzadła pierścienno-tchawiczego. W błonie śluzowej krtani, poza fałdami g ł o s o w y m i , występują liczne gruczoły cewkowe i cewkowo-pęcherzykowe o charakterze surowiczym i surowiczo-śluzowym. Gruczoły swoją wydzieliną zwilżają fałdy głosowe, jak również całe wnętrze krtani.
Tchawica Tchawica ( trachea) ma kształt sprężystej, spłaszczonej od tyłu rury, która g ó r n y m końcem łączy się z krtanią, dolnym przechodzi do wnętrza klatki piersiowej, gdzie dzieli się na dwa oskrzela główne. Na przekroju poprzecznym tchawica ma kształt zbliżony do podkowy. W odcinku g ó r n y m w y m i a r strzałkowy tchawicy przewyższa w y m i a r poprzeczny, w połowie długości oba w y m i a ry są sobie równe, w części dolnej w y m i a r poprzeczny jest większy od strzałkowego. Długość tchawicy u człowieka dorosłego waha się w granicach od 10 do 15 cm, szerokość od 13 do 22 mm. Tchawicę dzielimy na część szyjną i piersiową. Część szyjna łączy się z chrząstką pierścieniowatą krtani więzadłem pierścienno-tchawiczym. Od przodu sąsiaduje ona z mięśniami powierzchownymi szyi. Spłaszczona powierzchnia tylna tchawicy przylega do przełyku. W d o l n y m odcinku części szyjnej tchawica objęta jest przez tarczycę. Między przełykiem a tchawicą po obu stronach leżą: nerw błędny, tętnica szyjna wspólna i żyła szyjna wewnętrzna. W części szyjnej tchawica jest dostępna badaniu dotykiem. Poniżej o t w o r u górnego klatki piersiowej znajduje się część piersiowa tchawicy, zakończona rozdwojeniem tchawicy. Po stronie lewej piersiowej części tchawicy leży łuk aorty, od przodu pień ramienno-głowowy, a u dzieci również grasica, która u osób dorosłych ulega stłuszczeniu. Rusztowanie tchawicy tworzą sprężyste chrząstki tchawicze, które utrzymują drożność światła tchawicy, nie pozwalając na zapadanie się jej ścian. Każda z chrząstek ma kształt podkowy wypukłością zwróconej ku przodowi. Poszczególne chrząstki łączą się ze sobą za pomocą więzadeł obrączkowych. '251
Tylna, przylegająca do przełyku, część tchawicy nie zawiera chrząstek i nosi nazwę ściany błoniastej. Ściana błoniasta zawiera również włókienką mięśni gładkich, z których jedne, leżące od wewnątrz, łączą w o l n e końce chrząstek tchawiczych, inne mają przebieg podłużny. Błona śluzowa wyściełająca wnętrze tchawicy pokryta jest nabłonkiem wielorzędowym cylindrycznym migawkowym. W błonie śluzowej, szczególnie pokrywającej ścianę błoniastą, widać liczne gruczoły tchawicze o budowie cewkowej i cewkowo-pęcherzykowej.
Oskrzela Tchawica na wysokości IV krążka międzykręgowego dzieli się na dwa oskrzela g ł ó w n e (bronchi principales), rozchodzące się pod kątem 90°. Oskrzele prawe jest grubsze, krótsze (3-5 cm) i przebiega bardziej pionowo, leżąc niejako w przedłużeniu tchawicy. Lewe, o długości 5 - 8 cm, jest cieńsze i biegnie bardziej poziomo. Wskutek tego ciała obce, które dostały się do tchawicy, częściej wpadają do oskrzela prawego. Stosunki topograficzne obu oskrzeli są odmienne. Oskrzele główne prawe jest objęte od góry łukiem żyły nieparzystej, która biegnie z tylnej ściany klatki piersiowej do żyły głównej górnej. Nad oskrzelem g ł ó w n y m lewym przerzuca się od przodu ku t y ł o w i łuk aorty. Budowa oskrzeli głównych jest podobna do budowy tchawicy. Występują tu również podkowiaste chrząstki, połączone ze sobą więzaałami. Tylną część oskrzeli głównych stanowi ściana błoniasta. Błonę śluzową pokrywa nabłonek wielorzędowy migawkowy, zawierający gruczoły oskrzelowe.
Płuca Płuca ( pulmones ), leżące w klatce piersiowej, są właściwym narządem oddechowym, w którym dochodzi do w y m i a n y gazów między krwią a powietrzem. Płuca mają kształt stożków, obciętych z jednej strony. Górna część płuca nazywa się szczytem, dolna podstawą. Oba płuca różnią się nieco między sobą: płuco prawe jest krótszeł szersze od lewego. Na każdym płucu wyróżniamy trzy powierzchnie: przeponową, żebrową i przyśrodkową. Kształt ich jest dostosowany do zarysów ścian klatki piersiow e j i sąsiadujących z płucami narządów. Powierzchnia przeponowa jest wklęsła, powierzchnia żebrowa wypukła, powierzchnia przyśrodkową, zwrócona do śródpiersia i kręgów, na pewnej przestrzeni przylega do serca. Miejsce to, nieco zagłębione, nosi nazwę wycisku sercowego. Powierzchnie płuc oddzielone są od siebie brzegami. Brzeg dolny oddziela powierzchnię żebrową i przyśrodkową od przeponowej. Brzeg przedni leży na granicy powierzchni żebrowej i przyśrodkowej. Na płucu lewym w dolnej części tego brzegu widnieje wcięcie sercowe płuca lewego. Część kręgowa '252
powierzchni przyśrodkowej jest miejscem stopniowego przejścia powierzchni przyśrodkowej w powierzchnię żebrową od tyłu. Każde płuco jest podzielone na płaty ( lobi ). Płuco prawe liczy ich trzy: płat górny, ś r o d k o w y i dolny. Płuco lewe ma dwa płaty - górny i dolny. Płaty płuc oddzielone są szczelinami. Jedna z nich, zwana szczeliną skośną, przebiega jednakowo w o b y d w u płucach. Rozpoczyna się w okolicy wnęki, biegnie ku górze i ku tyłowi, przechodząc na tylną powierzchnię płuc na wysokości wyrostka kolczystego III kręgu piersiowego. Stąd schodzi po powierzchni żebrowej ku dołowi i ku przodowi, dochodząc do przedniego brzegu płuca w okolicy chrząstki VI żebra. Od tego miejsca podąża ku górze i ku tyłowi, osiągając wnękę. Szczelina pozioma znajduje się jedynie w płucu prawym i jest znacznie płytsza od szczeliny skośnej. Biegnie poziomo wzdłuż IV żebra od brzegu przedniego płuca do linii pachowej prawej. Z przebiegu szczeliny wynika, iż patrząc na płuco prawe od przodu widzimy jedynie płat górny i płat środkowy. Z boku widoczne są trzy płaty, od tyłu - dolny i część górnego, odpowiadająca szczytowi płuca. Każdy z płatów rozpada się na odrębne jednostki czynnościowe, zwane segmentami oskrzelowo-płucnymi, do których prowadzą odpowiadające im
Ryc. 130. Płuco prawe od strony przyśrodkowej. Zwróć uwagę na ułożenie głównych oskrzeli w stosunku do naczyń. 17-Anatomia i fizjologia
253
oskrzela segmentowe. Granice między poszczególnymi segmentami nie są tak wyraźnie zaznaczone, jak granice płatów, niemniej segmenty są jednostkami autonomicznymi, mającymi własne oskrzela i własne naczynia. W każdym płucu wyróżnia się 10 segmentów. Znajomość granic segmentów płucnych jest szczególnie ważna w chirurgii klatki piersiowej, ponieważ pozwala na operacyjne usunięcie zmienionej części płuca, obejmującej zwykle jeden lub kilka segmentów. Na przyśrodkowej powierzchni płuca znajduje się miejsce nie pokryte opłucną, zwane wnęką płuca (hilus pulmonis). Przez wnękę przechodzą do płuc: tętnica płucna, gałęzie oskrzelowe aorty piersiowej, oskrzela i nerwy. Opuszczają płuca: żyły płucne, żyły oskrzelowe i naczynia limfatyczne. T w o r y , które wnikają do płuc bądź też z nich wychodzą w obrębie wnęki, noszą wspólną nazwę korzenia płuc (radix pulmonis). Kolejność składników korzenia płuc jest inna po stronie prawej niż po lewej. Po stronie prawej, idąc od góry, napotykamy kolejno: oskrzele, tętnicę i żyłę. Po stronie lewej: tętnicę, oskrzele i żyłę. Barwa płuc zależy od szeregu czynników, które zmieniają się w ciągu życia. Bezpowietrzne płuca płodu z p o w o d u dużej zawartości krwi mają barwę ciemnoczerwoną; noworodek, który zaczerpnął powietrza, ma płuca bladoróżowej barwy. W miarę upływu czasu do pęcherzyków płucnych dostają się
T. płuc Oskrzele górne prawe Ż.ż. płucne
Ryc. 131. Płuco lewe od strony przyśrodkowej.
'254
drobniutkie cząsteczki ciał stałych (pyłki) i płuco przybiera barwę zależną od rodzaju tych pyłków, najczęściej szaroniebieskawą. U górników oddychających powietrzem z zawartością pyłków w ę g l o w y c h barwa płuc jest stalowogranatowa. Jeżeli człowiek oddycha powietrzem silnie zapylonym (kamieniarze, stolarze, górnicy, zecerzy), powstaje u niego choroba płuc, zwana pylicą (pneumoconiosis). Płuca płodu niezawierają powietrza, ich ciężar właściwy wynosi 1,060. Płuca takie wrzucone do w o d y toną. Po wykonaniu przez noworodka pierwszego wdechu ciężar właściwy płuc zmniejsza się do 0,490 i płuca takie wrzucone do w o d y utrzymują się na powierzchni. W związku z niewielkim ciężarem właściwym ciężar płuc człowieka dorosłego w y n o s i zaledwie około 1000 g u kobiet i około 1300 g u mężczyzn.
Topografia płuc Jamę klatki piersiowej można podzielić na trzy części. Dwie z nich zajęte są przez płuca, w trzeciej mieści się serce oraz inne narządy śródpiersia. Szczyty płuc wystają nieco ponad I żebro i obojczyk, tak że zranienia przyśrodkowej części okolicy nadobojczykowej mogą przebiegać z uszkodzeniem płuc. Do celów klinicznych konieczne jest poznanie rzutu brzegów płuc na ścianę klatki piersiowej. Brzeg przedni płuca prawego rozpoczyna się na wysokości stawu obojczykowo-mostkowego prawego, biegnie ku dołowi i przyśrodkowo, tak że przy wdechu w okolicy II żebra przechodzi nieco na lewo od linii środkowej ciała, po czym kieruje się ku dołowi. Na wysokości IVżebra.odgina się trochę w prawo, aby na żebrze VI przejść w granicę dolną. W linii środkowo-obojczykowej rzut dolnego brzegu płuca prawego znajduje się na VI żebrze, w linii pachowej na VII żebrze, w linii łopatkowej krzyżuje się z X żebrem, w pośrodkowej tylnej znajduje się na wysokości XI kręgu piersiowego. W płucu lewym rzut brzegu przedniego jest początkowo podobny, przebiega jednak bardziej na lewo od linii środkowej ciała. Dopiero na g ó r n y m brzegu IV żebra odgina się silnie w lewo, dochodzi do linii przympstkowej i zawraca ostro ku dołowi. Od dolnego brzegu VI żebra rozpoczyna się granica dolna. Taki przebieg granicy przedniej uwarunkowany jest obecnością serca. Ta b e l a 19 Linia
Płuco środkowo-obojczykowa
pachowa
łopatkowa
pośrodkowa tylna
Prawe
VI żebro
VII żebro
X żebro
XI kręg piersiowy
Lewe
VII żebro
VIII żebro
X żebro
XI kręg piersiowy
'255
Rzut dolnego brzegu płuca lewego początkowo przebiega niżej niż po przeciwnej stronie. Rzut ten w linii środkowo-obojczykowej znajduje się na VII żebrze, w linii pachowej na VIII, w linii łopatkowej i kręgowej przebiega na takim s a m y m poziomie, jak po stronie prawej. Granice rzutów mogą zmieniać się na szerokości jednego żebra w zależności od położenia ciała, napięcia mięśni powłok brzusznych itd. A b y ułatwić zapamiętanie rzutów brzegów dolnych płuc, zestawiono je w tab. 19.
Budowa oskrzeli i płuc Oskrzela główne prawe i lewe dzielą się na corazto drobniejsze odgałęzienia dochodzące do pęcherzyków płucnych. Oskrzela główne, płatowe, a częściowo i segmentowe, zbudowane są podobnie do tchawicy. Znajdujemy tu chrząstki również podkowiastego kształtu, tylko odpowiednio niniejsze. Część oskrzela nie zajętą przez chrząstki wypełnia ściana błoniasta. Błonę śluzową w y m i e n i o n y c h oskrzeli pokrywa nabłonek wielorzędowy migawkowy. W mniejszych oskrzelach (gałęzie podsegmentowe, małe i najmniejsze) pierwotnie wyrażona budowa ścian ulega zatarciu. Chrząstki stają się coraz cieńsze, aż do małych płytek o nieregularnych kształtach. W oskrzelach tych T a b e l a 20 Schemat rozgałęzienia oskrzeli Droga oddechowa
Odpowiadająca część oskrzeli
Oskrzele główne Oskrzele płatowe Oskrzele segmentowe Gałęzie podsegmentowe małe i najmniejsze
płuco płat płucny segment oskrzelowo-płucny część segmentu
Oskrzelka Oskrzelka końcowe Oskrzelka oddechowe
zraziki grona pęcherzyki płucne
nie spotyka się już zupełnie chrząstek, zaś nabłonek błony śluzowej staje się coraz cieńszy i traci rzęski. W ścianach pojawia się więcej mięśni gładkich. Odcinek płuc zaopatrzony przez jedno oskrzelko nosi nazwę zrazika. Zraziki oddzielone są od siebie warstewką tkanki łącznej. Oglądane na powierzchni płuca przedstawiają się jako pólka o średnicy ok. 10 m m . Z wnikającego do zrazika oskrzelka powstają oskrzelka końcowe. Każde oskrzelko końcowe zaopatruje grono, które jest podstawową jednostką tkanki płucnej. W obrębie grona znajdują się oskrzelka oddechowe, ślepo zakończone woreczkami, na których ścianach znajdują się uwypuklenia, zwane pęcherzykami płucnymi (alveoli pulmonis). Ściany pęcherzyków są bardzo cienkie, zbudowane z jednej '256
warstwy komórek. Pęcherzyki otacza gęsta sieć naczyń włosowatych. Przez ścianki pęcherzyków odbywa się wymiana gazów między krwią a powietrzem pęcherzykowym.
Ryc. 132. Budowa płuca.
Unaczynienie i unerwienie płuc Naczynia, którymi krew dopływa do płuc, są dwojakiego rodzaju. Tętnice płucne, odchodzące od pnia płucnego, prowadzą krew żylną (odtlenioną), cienkie gałęzie oskrzelowe od aorty - krew tętniczą. Do każdego płuca dochodzi tętnica płucna i kilka cienkich gałęzi oskrzelowych. Krew z tętnic płucnych dostaje się do naczyń włosowatych, które oplatają pęcherzyki płucne i po oddaniu dwutlenku węgla i pobraniu tlenu przechodzi do żył płucnych. Gałęzie oskrzelowe zaopatrują w krew ściany oskrzeli i tkankę łączną płuc. Stąd krew odpływa do żył oskrzelowych, uchodzących do układu żyły nieparzystej. Między układem tętnicy płucnej a układem tętnic oskrzelowych istnieją liczne połączenia. Nerwy zaopatrujące płuca należą do układu współczulnego i przywspółczulnego.
'257
Opłucna Płuca oraz wewnętrzną powierzchnię klatki piersiowej pokrywa cienka, gładka^ lśniąca błona, zwana opłucną (pleura). Opłucna należy do błon surowiczych. Zbudowana jest z cienkiej w a r s t w y tkanki łącznej i pokrywającego ją nabłonka śródjamowego. Komórki nabłonka mają kształt wielokątny, są płaskie i ułożone w jednej warstwie. Opłucna dzieli się na opłucną ścienną (pleura parietalis) i opłucną płucną (pleura pulmonalis). Obydwie blaszki opłucnej oddziela szczelinowata przestrzeń, zwana jamą opłucnej. W jamie tej znajduje się niewielka ilość płynu surowiczego, który zmniejsza tarcie w czasie ruchów oddechowych płuc. Opłucna ścienna przechodzi w opłucną płucną na korzeniu płuca oraz w linii biegnącej od niego ku tyłowi i dołowi. Dwie blaszki opłucnej, biegnące wzdłuż wspomnianej linii, tworzą więzadło płucne. Opłucna ścienna przyjmuje nazwy od narządów, do których przylega. Część pokrywająca ścianę klatki piersiowej nosi nazwę opłucnej żebrowej. Opłucna przeponowa wyściela przeponę. Opłucna śródpiersiowa pokrywa przyśrodkową ścianę jamy opłucnej i narządy śródpiersia. Część opłucnej, odpowiadająca szczytowi płuca, nazywa się osklepkiem opłucnej. W miejscu przejścia opłucnej żebrowej w opłucną przeponową znajduje się szczelinowaty zachyłek żebrowo-przeponowy. Podobnie w miejscu, w którym opłucna żebrowa przechodzi w śródpiersiową, powstaje zachyłek żebrowo-śródpiersiowy. Zachyłki opłucnej są przestrzeniami zapasowymi, do których wchodzą płuca podczas wdechu. Najgłębszy jest zachyłek żebrowo-przeponowy i w nim najczęściej zbiera się płyn, np. podczas wysiękowego zapalenia opłucnej. Zachyłek jest na tyle głęboki, że płuco nie wypełnia go nawet w czasie najgłębszego wdechu. Opłucna płucna przylega ściśle do tkanki płuca i wchodzi do szczelin międzypłatowych. W jamie opłucnej panuje ciśnienie ujemne (niższe od atmosferycznego), a ponieważ w płucach znajduje się powietrze pod ciśnieniem atmosferycznym, płuca są rozdęte i przylegają do ścian klatki piersiowej. Dostanie się powietrza do jamy opłucnej i w y r ó w n a n i e ciśnień prowadzi do zapadnięcia się płuca z powodu obecności w tkance płucnej dużej ilości włókien sprężystych. Stan taki nazywa się odmą. Znajomość granic opłucnej, podobnie jak granic płuc, jest niezbędna w pracy klinicznej. Rzut granic opłucnej ściennej na ścianę klatki piersiowej T a b e l a 21 Linia Opłucna
środkowo-obojczykowa
pachowa
łopatkowa
środkowa tylna
Prawa
VII żebro
IX żebro
XI żebro
XII kręg piersiowy
Lewa
VII żebro
X żebro
XI żebro
XII kręg piersiowy
'258
różni się od rzutu granic płuc ze względu na obecność zachyłków. Przednia granica opłucnej ściennej biegnie początkowo równolegle do granicy płuc. Na wysokości od II do IV żebra opłucna ścienna strony lewej styka się z opłucną ścienną strony prawej. Linia zetknięcia biegnie pionowo, nieco na lewo od linii środkowej ciała. Rzut dolnej granicy opłucnej ściennej przedstawia tab. 21. Opłucna ścienna jest bogato unerwiona czuciowo przez gałęzie n e r w ó w międzyżebrowych i przeponowych, dlatego też zapalenie opłucnej powoduje silne bóle w klatce piersiowej.
Śródpiersie Śródpiersie (mediastinum) jest to przestrzeń leżąca między obiema blaszkami opłucnej śródpiersiowej. Od przodu jest ono ograniczone przez mostek, od tyłu przez kręgosłup, od dołu przez przeponę. Od góry jako granicę śródpiersia przyjmujemy płaszczyznę górnego otwOru klatki piersiowej. W śródpiersiu mieści się wiele narządów, a przestrzeń między nimi wypełnia tkanka łączna. W dolnym odcinku śródpiersia znajduje się serce w worku t>sierdziowym. W odcinku g ó r n y m leży część wstępująca aorty wraz z głównymi naczyniami (żyłą główną górną, tętnicami i żyłami płucnymi) oraz piersiowa część tchawicy z oskrzelami g ł ó w n y m i . U młodych osobników tuż za mostkiem napotykamy grasicę. W wieku starszym zamiast niej można znaleźć tka n kę tłu szczo wą. Między osierdziem a opłucną ścienną przebiegają nerwy przeponowe. W części tylnej śródpiersia znajduje się przełyk, na którego przedniej i tylnej powierzchni przebiegają nerwy błędne. Na lewo od przełyku znajduje się część zstępująca aorty. Między przełykiem a kręgosłupem leży przewód piersiowy. Nieco w prawo od niego biegnie żyła ramienno-głowowa, zdążająca do żyły głównej górnej. W śródpiersiu znajdują się ponadto liczne węzły chłonne.
FIZJOLOGIA UKŁADU ODDECHOWEGO W ustroju człowieka nieprzerwanie odbywają się procesy przemiany materii i energii. Energię czerpie ustrój z procesów spalania pokarmów, które zachodzi w warunkach tlenowych. Tlen do tych procesów pobierany jest z powietrza w procesie oddychania. W płucach tlen przechodzi do krwi, która roznosi go po całym organizmie. W komórkach ciała na skutek procesów utleniania powstają produkty przemiany materii, przede wszystkim dwutlenek węgla. Dostaje się on do krwi, z której znowu poprzez płuca zostaje wydalony na zewnątrz. Tak więc rozróżniamy: 1. Oddychanie płucne [albo zewnętrzne), polegające na w y m i a n i e tlenu i dwutlenku węgla między krwią a powietrzem. '259
2. Oddychanie tkankowe (albo wewnętrzne), polegające na wymianie tlenu i dwutlenku węgla między krwią a tkankami. Oddychanie tkankowe omówiono szczegółowo w rozdziale o przemianie materii i energii.
Oddychanie płucne Powietrze wdychane do płuc przez nos nasyca się parą wodną i oczyszcza z kurzu. Przechodząc między małżowinami a przegrodą nosa powietrze styka się z wilgotną błoną śluzową. Osadzają się na niej drobne zanieczyszczenia, które pokryty rzęskami nabłonek nosa przesuwa w kierunku gardła. Zawirowanie powietrza w nosie powoduje, że zostają tu zatrzymane większe cząsteczki kurzu, drobniejsze zaś przechodzą dalej i osiadają na ściankach drzewa oskrzelowego i pęcherzyków. Bardzo małe cząsteczki, o średnicy mniejszej od 0,5 (xm, utrzymują się w powietrzu i wraz z nim są wydalane z dróg oddechowych. Zanieczyszczenia osiadające na ścianach dróg oddechowych usuwane są ze śluzem przez jamę ustną lub nosową. Dzieje się to podczas kaszlu i kichania. Odruch kaszlowy jest niezbędny dla prawidłowej czynności płuc. Zapoczątkowuje go podrażnienie zakończeń czuciowych w błonie śluzowej, po czym następuje: a) głęboki wdech; b) zamknięcie nagłośni i szpary głosowej; c) skurcz mięśni brzucha i zwiększenie tłoczni brzusznej, a jednocześnie napinanie się mięśni wydechowych; prowadzi to do wzrostu ciśnienia w płucach do 100 mm Hg i wyżej; d) nagłe otwarcie szpary głosowej i nagłośni powoduje gwałtowny wyrzut powietrza, które porywa zanieczyszczenia znajdujące się na ściankach drzewa oskrzelowego. Odruch kichania ma podobny przebieg, a zapoczątkowuje go podrażnienie błony śluzowej nosa. Oddychać można również przez jamę ustną; nie jest to jednak objaw prawidłowy i występuje przeważnie przy nieżytach nosa. W tym przypadku do dróg oddechowych wchodzi powietrze zimne i nie oczyszczone, co może prowadzić do rozmaitych schorzeń. Przez tchawicę, oskrzela, oskrzelka i oskrzelka oddechowe powietrze dostaje się do pęcherzyków płucnych. Mają one ścianki zbudowane z jednej warstwy nabłonka i są gęsto oplecione naczyniami włosowatymi, co stwarza należyte warunki do wymiany gazowej między krwią a powietrzem.
Mechanika oddychania Klatka piersiowa tworzy przestrzeń zamkniętą, wypełnioną przez płuca, serce i duże naczynia. Opłucna płucna i opłucna ścienna przylegają do siebie, a znajdująca się między nimi szczelina zawiera nieco płynu tkankowego. Tak zwana jama opłucnej jest przestrzenią istniejącą potencjalnie (jedynie po zapadnięciu się płuca). Przyleganie do siebie listków opłucnej zapewnia ujemne (niższe od atmosferycznego) ciśnienie panujące w szczelinie znajdującej się między nimi. Istnie'260
nie niższego niż atmosferyczne ciśnienia jest możliwe dzięki sprężystości tkanki płucnej, która ma skłonność do zmniejszania swej objętości. Z tego samego powodu ciśnienie w tkankach otaczających pęcherzyki płucne jest również ujemne, podczas kiedy ciśnienie w pęcherzykach płucnych i drogach oddechowych jest równe atmosferycznemu. Różnica ciśnień w pęcherzykach i w tkankach otaczających sprzyja wymianie gazowej. Przy zmniejszonej elastyczności tkanki płucnej u ludzi starszych wymiana gazowa jest utrudniona. Podczas cyklu oddechowego zmienia się objętość klatki piersiowej wskutek zwiększania się jej trzech głównych wymiarów: przednio-tylnego, poprzecznego i pionowego. Wymiar przednio-tylny zwiększa się wskutek oddalania się mostka od kręgosłupa dzięki działaniu mięśni unoszących mostek przez podniesienie pierwszego żebra połączonego z mostkiem za pomocą chrząstkozrostu. Powiększenie wymiaru poprzecznego jest wynikiem podnoszenia łukowato wygiętych ku dołowi żeber środkowych. Powiększenie wymiaru pionowego jamy klatki piersiowej następuje w wyniku obniżenia się przepony. W czasie skurczu przepona uciska na narządy leżące w jamie brzusznej i dlatego podczas oddychania obserwuje się rytmiczne uwypuklanie się ścian brzucha. Wydech jest w warunkach prawidłowych fazą bierną oddychania. W wyniku rozkurczu mięśni wdechowych żebra opuszczają się dzięki sprężystości chrząstek żebrowych. W wydechu bierze również udział tłocznia brzuszna, która, uciskając na trzewa, unosi przeponę ku górze. Tkanka płucna, zawierająca włókna sprężyste, kurczy się i powietrze z pęcherzyków płucnych przez drogi oddechowe wydostaje się na zewnątrz. Przy silnym wydechu i w niektórych stanach chorobowych połączonych z dusznością wydechową w wydechu biorą udział również mięśnie wydechowe obniżające żebra. Czynniki wpływające na ciśnienie w jamie opłucnej Jednocześnie ze zmianami objętości klatki piersiowej zmienia się również ciśnienie w jamie opłucnej. Podczas wdechu wynosi ono - 8 mm Hg, w fazie wydechu podnosi się do - 2 mm Hg. Średnie ciśnienie w jamie opłucnej wynosi zatem - 5 mm Hg. Drugą przyczyną ujemnego ciśnienia są siły osmotyczne. W naczyniach włosowatych sąsiadujących z opłucną płucną znajdują się ciała białkowe i koloidalne, dzięki czemu dochodzi do wchłaniania płynu z przestrzeni między opłucnymi. Dzieje się tak wskutek różnicy ciśnień osmotycznych. Ciśnienie osmotyczne działa stale zarówno podczas wdechu, jak i wydechu i wynosi ok. 20 m m Hg. Trzecim czynnikiem utrzymującym ujemne ciśnienie w jamie opłucnej jest tzw. pompa limfatyczna. Bez odpływu płynu z przestrzeni między listkami opłucnej gromadziłyby się w niej białka zaburzające równowagę ciśnienia osmotycznego. Ciągły ruch płuc, bicie serca przesuwają płyn do naczyń chłonnych. Usuwanie wraz z płynem białek przywraca prawidłowe ciśnienie osmotyczne. '261
Pojemność płuc Podczas oddychania w spoczynku z częstością 14-18 cyklów na minutę przy każdym wdechu napływa do płuc 350-500 ml powietrza. Nazywa się ono powietrzem oddechowym. Ilość powietrza, którą można jeszcze pobrać po wykonaniu przeciętnego wdechu, wynosi ok. 2500 ml i stanowi powietrze dopełniające.
G
O
Ryc. 134. Schemat spirometru.
'262
T a b e l a 22
Wdech
powietrze dopełniające pojemność oddechowa
Wydech
Razem
2500 ml
500 ml
powietrze zapasowe
1000 ml
powietrze zalegające
1200 ml
pojemność całkowita
5200 ml
Powietrze zapasowe jest to ilość powietrza, które można usunąć z płuc po wykonaniu przeciętnego wydechu (ok. 1000 ml). Część pozostałą, nie dającą się usunąć, nazywamy powietrzem zalegającym (1200 ml). Suma powietrza oddechowego, dopełniającego i zapasowego wynosi ok. 4000 ml i określa pojemność życiową płuc. Ze względu na powietrze zalegające pojemność całkowita płuc nie równa się pojemności życiowej, lecz jest większa i wynosi ok. 5200 ml (patrz tab. 22). Z 500 ml powietrza pobranego w czasie przeciętnego wdechu tylko ok. 360 ml dostaje się do pęcherzyków płucnych. Pozostałe 140 ml zalega w drogach oddechowych, wypełniając tzw. przestrzeń martwą i nie bierze udziału w wymianie gazowej. Pojemność płuc mierzymy spirometrem. Przyrząd składa się z dwóch naczyń o kształcie walca, przy czym jedno z nich, odwrócone dnem do góry, znajduje się w drugim. Do walca dolnego nalana jest woda, wypełniająca również walec górny. Powietrze wydychane do wody przez rurę zbiera się pod walcem górnym i unosi go. Ilość powietrza odczytujemy na skali (ryc. 134).
Krążenie płucne Wymiana gazowa w płucach zachodzi za pośrednictwem krwi krążącej w obiegu małym (płucnym). Prawa komora serca tłoczy krew do płuc.Niewielka objętość krwi pompowanej przez komorę lewą przepływa przez naczynia oskrzelowe. Objętość krwi przetłoczonej przez płuca w jednostce czasu równa jest objętości krwi przechodzącej przez obieg wielki. Ciśnienie w czasie skurczu prawej komory jest znacznie niższe niż w komorze lewej. Wzrasta ono do ok. 22 mm Hg w skurczu i spada przy rozkurczu do 0 - 1 mm Hg. (W komorze Ifewej odpowiednie ciśnienia wynoszą 160 i 70 mm Hg). Podczas skurczu ciśnienie w tętnicy płucnej jest równe ciśnieniu w komorze. 263
Po zamknięciu za stawek tętnicy płucnej ciśnienie spada do 13 mm Hg, podczas gdy na poziomie włośniczek obniża się do 8 mm Hg. Końcowe gałęzie tętnicy płucnej przechodzą w mające liczne połączenia naczynia włosowate. W układzie tętnic płucnych brak jest systemu naczyń pośrednich, o rozwiniętym układzie zwieraczy, mogących regulować dopływ krwi do określonych tkanek. Zarówno tętnice płucne, jak i tętnice oskrzelowe kończą się w tej samej sieci naczyń włosowatych, z których krew odpływa do żył płucnych. Naczynia włosowate pęcherzykowe wchodzą w skład ściany błony pęcherzyków płucnych. Ich średnica wynosi ok. 8 nm, a sieć ich jest tak gęsta, że odległości między nimi są mniejsze od średnicy naczyń włosowatych. Otoczone przez włośniczki pęcherzyki płucne mają kształt półkuli o powierzchni od 0,075 do 0,125 mm 2 , a ich liczba sięga 750 000 000. Łączna powierzchnia oddechowa pęcherzyków wynosi ok. 90 m 2 (I). Ciśnienie w naczyniach włosowatych płucnych wynosi średnio ok. 5 mm Hg. Krew przesuwa się w nich z szybkością 0,8 mm/s. Przy wzroście rzutu sercowego (p. str. 389) szybkość przechodzenia krwi przez naczynia włosowate nie zmienia się, ponieważ zwiększona objętość krwi przepływa przez naczynia w w a r u n kach prawidłowych zapadnięte. Krew przechodząca przez włośniczki oddaje C0 2 i pobiera tlen. Jednocześnie do pęcherzyków nie przedostają się białka krwi, ponieważ ciśnienie osmotyczne w naczyniach (28 mm Hg) jest wielokrotnie wyższe niż w pęcherzykach płucnych (5 mm Hg). Duża różnica ciśnień sprzyja natomiast resorpcji płynu z otoczenia naczyń włosowatych. Żyły płucne są krótkie i nie przystosowują się tak do objętości krwi, jak naczynia krążenia wielkiego. Ilość krwi krążącej w płucach wynosi od 12 do 16% ogólnej ilości krwi. Każde z płuc dorosłego człowieka zawiera od 300 do 400 ml krwi, z tego tylko 60-80 ml wypełnia naczynia włosowate, a reszta znajduje się w dużych naczyniach. W zależności od zmian ciśnienia w klatce piersiowej zmienia się objętość krwi w płucach. Na przykład podczas dęcia w instrument muzyczny może być wyparte z płuc 350 ml krwi, podczas ssania objętość krwi w płucach zwiększa się.
Wymiana gazów Mechanizm wymiany gazów możemy śledzić badając skład powietrza wdychanego i wydychanego (tab. 23). Ucieczka tlenu z powietrza pęcherzykowego i wędrówka dwutlenku węgla w kierunku odwrotnym zachodzi zgodnie z prawami fizyki. Jedno z tych praw mówi, że ciśnienie cząstkowe gazu w mieszaninie wynosi taką samą część ogólnego ciśnienia mieszaniny gazów, jaka wynika z procentowej zawartości gazu w mieszaninie. Na przykład ciśnienie powietrza atmosferycznego wynosi przeciętnie 760 mm Hg. Procentowa zawartość tłenu w powietrzu równa się 20,96%, zatem ciśnienie cząstkowe (parcjalne) tlenu wynosi 760 • 20,96 = 159,2 m m Hg. 100 Ciśnienie cząstkowe poszczególnych gazów w powietrzu atmosferycznym przedstawia tab. 24. '264
T a b e l a 22 Zawartość gazu w odsetkach (%) powietrze atmosferyczne
powietrze wydychane
powietrze pęcherzykowe
20,96
16,3
14,2
Dwutlenek węgla
0,04
4,0
5,5
Azot wraz z innymi gazami
79,00
79,7
80,3
100,00
100,00
100,00
Tlen
Razem
T a b e I a 24 Ciśnienie cząstkowe w mm Hg Gaz
krew tętnicza
krew żylna 40 46
100 40
Tlen Dwutlenek węgla
różnica + 60 - 6
Różnica prężności gazów, przedstawiona w tab, 24, w zupełności wystarcza do zaspokojenia potrzeb ustroju, które wynoszą w ciągu minuty ok. 500 ml tlenu. Decydujące znaczenie ma tu powierzchnia oddechowa płuc, która równa się sumie powierzchni wszystkich pęcherzyków płucnych i wynosi - jak wspomniano wyżej - ok. 90 m2. Przy tak dużej powierzchni już różnica 10 mm Hg wystarcza do przeniknięcia odpowiedniej ilości tlenu do krwi. Z tabeli 24 wynika, że różnica ta jest 6 razy większa. Dwutlenek węgla przenika przez ściankę pęcherzyków płucnych znacznie szybciej od tlenu i już różnica wynosząca ułamek milimetra wystarcza do zapewnienia wymiany gazów. Liczbę wskazującą, ile gramów gazu rozpuszcza się w 1 ml cieczy, nazywamy współczynnikiem rozpuszczalności. Jest ona dla dwutlenku węgla ok. 30 razy większa niż dla tlenu (tab. 25). T a b e l a 25 Gaz Tlen Dwutlenek węgla Azot
Współczynnik rozpuszczalności 0,023 0,510 0,011
Z drugiej strony wiadomo, że rozpuszczalność gazów w cieczach zależy od ciśnienia cząstkowego gazu,który styka się z cieczą, oraz od ciśnienia tego gazu w cieczy. Jeżeli ciśnienie cząstkowe gazu znajdującego się nad cieczą przewyższa ciśnienie tego gazu w cieczy, to następuje dalsze jego rozpuszcza265
nie. W warunkach odwrotnych gaz ulatnia się z cieczy aż do wyrównania ciśnień. Krew zawiera rozpuszczalny tlen i dwutlenek węgla, przy czym ilości rozpuszczonych gazów we krwi tętniczej i żylnej różnią się między sobą, podobnie różnią się ich ciśnienia cząstkowe (tab. 26). " Ta b e l a 26 Ciśnienie cząsteczkowe w mm Hg Gaz
Tlen Dwutlenek węgla Azot wraz z innymi gazami Razem
powietrze atmosferyczne
powietrze wydychane
159,2 0,3 600,5 760,0
116,2 28,5 615,3 760,0
v _
powietrze pęcherzykowe <"
101,0 40,0 619,0. 760,0
Rola krwi w procesie oddychania Podstawową rolę w przenoszeniu krwi i dwutlenku węgla odgrywają krwinki czerwone zawierające hemoglobinę. Obecność hemoglobiny w krwinkach czerwonych pozwala przenieść od 30 do 100 razy więcej tlenu i od 20 do 30 razy więcej dwutlenku węgla, niż gdyby gazy te rozpuszczały się jedynie w osoczu krwi. Powietrze, którym oddychamy, zawiera ok. 21 % tlenu. Wywiera on ciśnienie cząstkowe ok. 159 mm Hg (p. tab. 26). Powietrze, wchodząc do płuc, miesza się z gazami już obecnymi w pęcherzykach płucnych i nasyca się parą wodną, stąd ciśnienie cząstkowe tlenu w powietrzu pęcherzykowym obniża się do 101 mm Hg. Ciśnienie cząstkowe tlenu we krwi żylnej wchodzącej do włośniczek pęcherzykowych wynosi ok. 40 mm Hg. Różnica ciśnień wynosząca 64 mm Hg, powoduje szybką dyfuzję tlenu do krwinek czerwonych. Ciśnienia cząstkowe wyrównują się tu w ciągu mniej niż jednej sekundy, to jest w czasie, w którym przepływa krew przez naczynia włosowate płucne. Krwinki czerwone dochodzące do ścian pęcherzyka wysycone są tlenem w 70%, a opuszczające je w 97%. Ciśnienie cząstkowe dwutlenku węgla we krwi dochodzącej do pęcherzyków wynosi ok. 45 mm Hg, w powietrzu pęcherzykowym ok. 40 mm Hg. Różnica ciśnienia 5 mm Hg wystarcza do przechodzenia cząsteczek dwutlenku węgla z krwinek czerwonych przeż ścianki naczyń włosowatych do pęcherzyków. Z przedstawionych liczb wynika, że różnica ciśnień cząstkowych między krwią a powietrzem pęcherzykowym jest ok. 10 razy większa dla tlenu niż dla dwutlenku węgla. Z drugiej strony rozpuszczalność dwutlenku węgla w wodzie oraz zdolność jego dyfuzji jest ok. 20 do 30 razy większa niż rozpuszczalność tlenu. Tak więc w czasie 0,8 s dwutlenek węgla opuszcza krew pod działaniem Vi 0 części siły koniecznej dla przejścia tlenu. '266
Wymiana gazów między krwią a powietrzem jest największa na początku, kiedy różnica ciśnień jest duża, po czym maleje do zera w końcowym okresie zetknięcia się krwi ze ścianą pęcherzyka. Średni gradient ciśnień dla tlenu podczas prawidłowego oddychania wynosi 11 mm Hg.
Oddychanie tkankowe We włośniczkach tkankowych tlen odłącza się od hemoglobiny i przechodzi przez ściany naczyń włosowatych do płynu tkankowego,a następnie do komórek. Ciśnienie cząstkowe tlenu w początkowej części naczyń włosowatych wynosi 95 mm Hg, zaś w płynie międzykomórkowym 40 mm Hg. Różnica ciśnień 55 mm Hg powoduje przejście tlenu do przestrzeni międzykomórkowej. Ciśnienie cząstkowe tlenu w płynie tkankowym wynosi 40 mm Hg, a ciśnienie wewnątrz k o m ó r e k - 3 5 mm Hg, co pozwala na przenikanie do nich tlenu. Całkowita różnica między ciśnieniem tlenu w krwinkach czerwonych a ciśnieniem w komórkach tkanek wynosi 60 mm Hg i jest zbliżona do różnicy ciśnień cząstkowych w płucach (64 mm). Całkowita ilość tlenu związanego z hemoglobiną krwi tętniczej przy jej pełnym wysyceniu wynosi ok. 19,4 ml/100 ml krwi. Po przejściu przez naczynia włosowate tkanek spada do 14,4/100 ml krwi,a w tkankach pozostaje 5 ml tlenu uwolnionego z każdych 100 ml krwi. Następnie, gdy krew powróci do płuc, ok. 5 ml tlenu dyfunduje do każdych 100 ml krwi i hemoglobina uzyskuje ponownie wysycenie 97%. Podczas intensywnego wysiłku fizycznego komórki mięśniowe wykorzystują tlen z tak dużą szybkością, że jego stężenie w płynie międzykomórkowym spada do 15 mm Hg. Przy takim ciśnieniu tylko 4,4 ml może pozostać w połączeniu z hemoglobiną znajdującą się w 100 ml krwi. Ilość uwolnionego tlenu z każdych 100 ml krwi wynosi zatem 19,4-4,4 = 15. Podczas dużego wysiłku może być przetransportowane trzy razy więcej tlenu niż w warunkach życia codziennego. Z powyższego wynika, że wzrost zużycia tlenu przez tkanki zwiększa automatycznie ilość tlenu uwalnianegoz hemoglobiny. Liczbowy odpowiednik tej części hemoglobiny, która oddaje swój tlen przechodząc przez tkankowe naczynia włosowate,nazywa się współczynnikiem wykorzystania. W warunkach prawidłowych wynosi on ok. 27% hemoglobiny. Podczas dużego wysiku aż 77% hemoglobiny oddaje swój tlen. Jest to najwyższy współczynnik wykorzystania, nawet przy największym zapotrzebowaniu tlenowym tkanek. W celu obliczenia całkowitego zużycia w jednostce czasu zakładamy, że każde 100 ml krwi przenosi 5 ml tlenu. Jeżeli podstawowy minutowy rzut serca wynosi 5000 ml, to w ciągu minuty krew dostarcza 250 ml tlenu. Jak już mówiliśmy, wielkość ta może się zwiększać podczas dużego wysiłku i w niektórych chorobach nawet do 15 razy, z tego trzykrotnie wskutek większego wykorzystania tlenu i pięciokrotnie dzięki wzrostowi rzutu minutowego. Największa ilość transportowanego tlenu do tkanek wynosi zatem 250 x 15, tj. 3750 ml/min. u zdrowego dorosłego człowieka. '267
Adaptacja do dużego wysiłku, np. u sportowców, polegająca na wzroście ilości całkowitej hemoglobiny i rzutu minutowego serca może doprowadzić do zwiększenia transportu tlenu do 4,5 - 5 l/min.
Rola dwutlenku węgla w regulacji równowagi kwasowo-zasadowej Nieustanne wytwarzanie dwutlenku węgla w tkankach jest nieodłącznym następstwem przemiany materii - procesu, w którym węgiel zawarty w pokarmach utlenia się dostarczając energii koniecznej dla czynności komórek. Ponieważ usuwanie dwutlenku węgla z tkanek wymaga pewnego czasu, organizm funkcjonuje prawidłowo tak długo, dopóki jego poziom utrzymuje się w średnich wartościach. Duże odchylenia od tego poziomu są szkodliwe dla ustroju. Nadmiar dwutlenku węgla stanowi przy t y m większe niebezpieczeństwo niż jego niedobór. Istnieje kilka mechanizmów usuwania nadmiaru dwutlenku węgla, a mianowicie: 1) układ buforów kwasowo-zasadowych, 2) wydalanie przez płuca, 3) wydalanie przez nerki. Układ bufo rów tkankowych reaguje na zmiany stężenia C02 wciągu sekund. Układ oddechowy może dostosować się do nagłych zmian w ciśnieniu dwutlenku węgla w ciągu kilku minut. Nerki natomiast dopiero po kilku godzinach. Zarówno bufory tkankowe, jak i układ wydzielniczy nerek oddziałują na nadmiar dwutlenku węgla zobojętnieniem jonów wodorowych, uwalnianych przy rozpadzie kwasu węglowego. Przez płuca wydalany jestz ustroju molekularny dwutlenek węgla. Wwarunkach prawidłowych ilość jonów wodorowych w płynie pozakomórkowym wynosi ok. 0,00000001 w litrze. Liczbę tę można zapisać jako 107. Minus 7 jest logarytmem liczby 0,00000001. Mówimy, że liczba 0,00000001 (107) wyraża stężenie jonów wodorowych w roztworze, a jego pH wynosi 7. Ponieważ liczba ta stanowi logarytm o podstawie 10, to pH 7 wyraża dziesięciokrotnie wyższe stężenie jonów niż pH 8. Woda ma stężenie jonów wodorowych równe 7 i jest substancją obojętną. Roztwór kwaśny ma pH poniżej 7, a zasadowy powyżej 7. Prawidłowe pH płynu pozakomórkowego waha się w granicach 7,35 - 7,45, co świadczy o tym, że płyny ustrojowe są nieznacznie zasadowe. Gdy pH przekroczy 7,45, uważamy, że ustrój pozostaje w stanie zasadowicy (alkalozy), przy pH poniżej 7,35-kwasicy. Cały system układów buforowych przeciwdziała zmianom pH. Płyny ustrojowe rzadko stają się w znaczniejszym stopniu kwaśne lub zasadowe. Bufory można porównać do chemicznych gąbek wciągających lub uwalniających jony wodorowe. Istnieje szereg układów buforowych w organizmie: kwas węglowy, wodorowęglany, fosforany jednosodowe i dwusodowe, oksyhemoglobina i karbpksy'268
hemoglobina oraz białka osocza. Najważniejszym układem b u f o r o w y m jest układ kwas w ę g l o w y - w o d o r o w ę g l a n y . Wiele klinicznych zaburzeń równowagi kwasowo-zasadowej wynika ze zmian stosunki! kwasu w ę g l o w e g o H2C03 do jonu w o d o r o w ę g l a n o w e g o HC03. Gdy stosunek kwasu w ę g l o w e g o do w o d o r o w ę g l a n ó w ma się jak 1 : 20, pH płynów ustrojpwych jest prawidłowe. Krewtętnicza ma pH ok. 7,4; w miarę jak z krwią żylną dostaje się dwutlenkek węgla,powstaje kwas w ę g l o w y i pH krwi żylnej spada do ok. 7,35. Gdy dwutlenek węgla dyfunduje z naczyń włosowatych płucnych do pęcherzyków, pH krwi tętniczej wzrasta o 0,04 jednostki osiągając prawidłowe pH krwi tętniczej, tj. do 7,4. Za pomocą złożonych metod obliczono, że pH wnętrza komórki wynosi 7,0-7,2. Nasilona czynność komórki powoduje zwiększenie wytwarzania dwutlenku węgla i co za t y m idzie - obniża pH. Przy stałym wytwarzaniu C02 i prawidłowym krążeniu krwi p o d s t a w o w y m czynnikiem w p ł y w a j ą c y m na stężenie dwutlenku węgla są zmiany wentylacji pęcherzykowej. Ponieważ wzrost zawartości dwutlenku węgla powoduje spadek pR, zmiany wentylacji pęcherzykowej efektywnie zmieniają stężenie j o n ó w w o d o r o w y c h w ustroju. Dwukrotny wzrost wentylacji pęcherzykowej może podnieść pH płynów pozakomórkowych o 0,23 jednostki. Tak więc jeżeli pH płynów ustrojowych wynosiło 7,4 przy wentylacji prawidłowej, to jej podwojenie zmniejsza zawartość dwutlenku węgla i stężenia j o n ó w wodorowych, a pH wzrasta do 7,63. Odwrotnie - zmniejszenie wentylacji pęcherzykowej do 1 / 4 wartości prawidłowej powoduje zatrzymanie dwutlenku węgla i obniżenie pH z 7,4 do 7,0. Z powyższego wynika, w jaki sposób bufory ustrojowe przeciwstawiają się zmianom pH w warunkach zmian wentylacji płuc oraz jak niebezpieczne jest złe funkcjonowanie układu buforów.
Regulacja oddychania Oddychamy rytmicznie bez udziału świadomości. Można co prawda wstrzymać oddech, lecz jedynie na krótki czas. Mięśnie oddechowe otrzymują bodźce nerwowe z ośrodka oddechowego, leżącego w rdzeniu przedłużonym. W komórkach nerwowych tego ośrodka procesy fizykochemiczne wyzwalają regularne bodźce, które drogami n e r w o w y m i docierają do mięśni oddechowych. Przecięcie rdzenia przedłużonego poniżej ośrodka oddechowego prowadzi do bezdechu (aprtoe). W spoczynku człowiek dorosły oddycha ok. 16 razy na minutę. Liczba oddechów zmienia się w zależności od czynności ustroju, np. podwaja się w czasie forsownego marszu, a maleje we śnie. Podstawową rolę w procesie oddychania odgrywa pH krwi. Zakwaszenie (obecność dużej ilości C02) zwiększa liczbę oddechów, alkalizacja krwi zwalnia oddychanie. Zwiększenie zawartości dwutlenku węgla prowadzi do nasilenia ruchów oddechowych do czasu, aż zawartość C02 we krwi osiągnie poziom prawidłowy. Przeciwnie 18 - Anatomia i fizjologia
269
jeżeli w y k o n a m y kilka głębokich w d e c h ó w i stężenie C0 2 spadnie poniżej poziomu, występuje zahamowanie ruchów oddechowych, a nawet czasowy bezdech. Dużą rolę w regulacji oddychania odgrywa mechanizm odruchowy. Rozciągnięcie pęcherzyków płucnych przez powietrze podrażnia włókna dośrodkowe nerwu błędnego, po których pobudzenie dociera do ośrodka oddechowego. Ośrodek przestaje wysyłać bodźce do mięśni i wdech zostaje zahamowany. Stan ten trwa dopóty, dopóki pęcherzyki płucne nie zapadną się, gdyż wtedy znika działanie hamujące nerwu błędnego i ośrodek oddechowy wysyła bodźce przez nerwy odśrodkowe do mięśni oddechowych. W ten sposób wdech pociąga za sobą wydech i odwrotnie. Swoiste bodźce płyną do ośrodka oddechowego z miejsca, gdzie tętnica szyjna wspólna dzieli się na tętnicę szyjną wewnętrzną i zewnętrzną. W miejscu podziału znajduje się rozszerzenie, zwane zatoką tętnicy szyjnej (sinus caroticus), do której dochodzi gałąź nerwu językowo-gardłowego. Ściany zatoki są czułe na zmiany pH krwi. Wzrost C02 lub zmniejszenie ilości 0 2 we krwi wywołują zmiany wentylacji płuc.
'270
UKŁAD M O C Z O W Y
Układ moczowy ma do spełnienia dwojakie zadanie: 1) wydalanie z ustroju niepotrzebnych produktów przemiany materii i substancji szkodliwych oraz 2) utrzymanie stałości składu płynów ustrojowych. Do układu moczowego należą dwie nerki (prawa i lewa), wytwarzające mocz, wychodzące z nich moczowody, odprowadzające mocz z nerki do pęcherza moczowego, pęcherz moczowy oraz cewka moczowa. Nerka (ren, nephros) jest narządem parzystym o powszechnie znanym kształcie, ciężarze ok. 150 g, długości ok. 11 cm, szerokości ok. 5 cm i grubości
Nerka
Moczowód
m I Jajowód Macica • Jajnik
Ryc. 135. Budowa układu moczowo-płciowego żeńskiego.
'271
ok. 3 cm, konsystencji spoistej, barwy ciemnoczerwonej. Wyróżnia się w niej dwa bieguny - górny i dolny, dwa brzegi - boczny wypukły i przyśrodkowy wklęsły oraz dwie powierzchnie - przednią i tylną. Nerki - prawa i lewa - położone są na tylnej ścianie jamy brzusznej po obu stronach kręgosłupa na wysokości od XII kręgu piersiowego do II lędźwiowego. W położeniu t y m utrzymywane są dzięki obecności pokaźnego pokładu tkanki tłuszczowej tworzącej torebkę tłuszczową, połączoną tkanką łączną luźną z tylną ścianą jamy brzusznej. Przyczyniają się także do utrzymania nerek w ich pozycji duże pnie naczyniowe - tętnica i żyła nerkowa oraz ciśnienie panujące w jamie brzusznej. Dosyć luźne umocowanie nerek pozwala im na fizjologiczną r u c h o m o ś ć przesuwają się one przy głębokim wdechu oraz zajmują położenie nieco niższe w pozycji stojącej niż leżącej. Nerka prawa leży nieco niżej niż lewa.
BUDOWA NERKI Na przekroju podłużnym nerki widnieją dwie warstwy - zewnętrzna zwana korą nerki ( c o r t e x renis) i wewnętrzna - rdzeń nerki ( m e d u l l a renis). Rdzeń układa się w postaci trójkątnych pól, zwanych piramidami n e r k o w y m i (pyramides renales), pomiędzy które wchodzi kora tworząc tzw. słupy nerkowe. Wcięcie brzegu przyśrodkowego nadające nerce jej charakterystyczny
Ryc. 136
Ryc. 137
Ryc. 136. Nerka i nadnercze prawe (widok od przodu). Ryc. 137. Nerka i nadnercze lewe (widok od przodu).
'272
kształt nazywa się wnęką nerkową (hilus renalis). Przez wnękę wchodzą do nerki naczynia nerkowe i moczowód. We wnętrzu nerki bliżej wnęki znajduje się miedniczka nerkowa ( pelvis renalis), która na zewnątrz przechodzi w moczowód, a do wnętrza nerki rozgałęzia się na 2 - 3 kielichy nerkowe większe, które z kolei dzielą się na kielichy nerkowe mniejsze. Te ostatnie w liczbie ok. 10 obejmują zaokrąglone końce piramid nerkowych tworzących brodawki nerkowe.
CZYNNOŚĆ NEREK Jednostką czynnościową i strukturalną nerki jest nefron. Schemat nefronu przedstawiono na ryc. 138. Składa się on z ciałka nerkowego i wychodzącego zeń układu kanalikowego. Ciałko nerkowe zbudowane jest z pętli naczyń włosowatych tworzących kłębuszek nerkowy (Malpighiego) oraz otaczającej ten kłębuszek torebki ciałka nerkowego (Bowmana). Od torebki odchodzi kanalik kręty proksymalny, który następnie przechodzi w t z w . pętlę nefronu (Henlego) o dwóch ramionach-zstępującym i wstępującym. Ramię wstępujące pętli przechodzi w kanalik kręty dystalny (II rzędu), który uchodzi do cewki zbiorczej. W skład obu nerek człowieka wchodzi około 2,5 min w podobny sposób zbudowanych nefronów. Ponieważ budowa i czynności ich są podobne, można dla uproszczenia wyobrazić sobie czynność nerek jako zwielokrotnioną czynność pojedynczego nefronu, którą poniżej dokładniej rozpatrzymy. Kłębuszek naczyniowy ciałka nerkowego
Kanalik kręty
Kanalik kręty
Pętla nefronu
Cewka zbiorcza
Nacz^
odprc
Do torebki przesącza się woda, Na + , K t
Cr.HCOj,
glukoza
Zredukowanie przesączu kłębkowego do ok. 1/8 wskutek wchłaniania zwrotnego (reabsorpcji)
Zagęszczanie moczu
Oszczędzanie zasobu zasad na drodze wydalanie H + i NH 4 i reabsorpcji Na +
Ryc. 138. Schemat budowy i czynności nefronu.
'273
Nerki są bardzo obficie ukrwióYie. Przypływa przez nie około 25% krwi wyrzucanej przez serce z każdym skurczem, tj. około 1,2 l/min. Jest to znacznie więcej, niżby to wynikało z potrzeb metabolicznych samego narządu, którego ciężar nie przekracza 0,5% ciężaru ciała. Krew ta w nerkach ulega p e w n y m procesom fizykochemicznym,mającym na celu pozbycie się różnych substancji szkodliwych lub zbędnych dla ustroju drogą wydalenia ich z moczem na zewnątrz. Ponadto nerki pełnią bardzo ważną rolę w utrzymaniu stałości składu środowiska wewnętrznego ustroju - utrzymywania na mniej więcej stałym poziomie ciśnienia osmotycznego (osmolalności) płynów ustrojowych, stężenia j o n ó w w o d o r o w y c h (pH), a także funkcjonują jak narząd dokrewny, biorący udział w regulacji ciśnienia krwi, regulacji przemiany w a p n i o w e j oraz erytropoezie (wytwarzaniu krwinek czerwonych). Nadchodząca do nerek krew zostaje najpierw doprowadzona przez tętniczkę doprowadzającą (vas afferens) do splotu naczyniowego ciałka nerkowego. Tutaj mniej więcej jedna piąta objętości przepływającego osocza pod wpływ e m ciśnienia hydrostatycznego krwi przesącza się do torebki ciałka nerkowego, tworząc tzw. mocz pierwotny. Zatem z około 700 ml osocza, przepływającego w ciągu minuty przez obie nerki, ok. 120 ml, zostaje wyłączone z obiegu krwi przechodząc do torebek ciałek nerkowych, zaś pozostała część krwi w y p ł y w a z sieci kłębuszka naczyniowego przez tętniczkę odprowadzającą. Tętniczka ta ponownie rozpada się na sieć naczyń włosowatych oplatających t y m razem układ kanalików nefronu, które ostatecznie łączą się przechodząc w układ żylny. Przesączony w kłębuszku mocz pierwotny przechodzi do układu kanalikowego, gdzie ulega dalszym przemianom do moczu ostatecznego. Składają się na to dwa procesy - wchłanianie zwrotne, czyli reabsorpcja, oraz wydzielanie, czyli sekrecja. Nabłonek kanalikowy wchłania z powrotem do krwi w ciągu godziny około trzech czwartych w o d y i soli zawartych w całym osoczu. W kanalikach odbywa się także wydzielanie czynne, czyli sekrecja do moczu pierwotnego niektórych substancji, które podlegają wydalaniu z ustroju.
Przepływ krwi przez nerki Przepływu krwi przez nerki w całym nienaruszonym ustroju nie możemy zmierzyć bezpośrednio, istnieją jednak metody pośrednie pozwalające ten przepływ obliczyć. Są to tzw. techniki klirensowe, które mają duże znaczenie praktyczne i dlatego postaramy się je pokrótce przedstawić. Nerkowy przepływ krwi mierzysięza pomocą klirensu para-aminohipuranu. Klirens (clearance) po angielsku znaczy oczyszczanie. Para-aminohipuran (w skrócie PAH) jest substancją, która w ustroju nie występuje, odznacza s i ę t y m , że wprowadzona do krwi nie tylko przesącza się w kłębuszku, lecz także wydziela się czynnie w kanalikach tak wydajnie, że jeżeli dawka nie jest za wielka, to cała ilość PAH, jaka weszła z krwią do nerki, zostaje wydalona '274
T | v > 5
j
z moczem. Inaczej mówiąc, nerka oczyszcza z para-aminohipuranu w jednym pasażu całą krew, jaka przez nią przepływa. Zatem ilość PAH wchodząca do nerki w ciągu minuty równa się ilóści PAH opuszczającej nerki w tym samym czasie. Ilość PAH wydalonego z moczem w jednostce czasu (UV) można złatwością oznaczyć. W tym celu należy pomnożyć objętość moczu (V), wydalonego w jednostce czasu,np. w ciągu minuty, przez stężenie PAH w jednostce objętości moczu, np. w 1 ml (U). Z drugiej strony ilość PAH nadchodząca do nerki w jednostce czasu, np. w ciągu minuty, równa się objętości przepływającego w tym czasie osocza (przepływ) pomnożonej przez stężenie PAH we krwi (P) = przepływ x P. Możemy zatem napisać równanie: Przepływ x P = UV stąd: przepływ H Wykonanie klirensu polega na wstrzyknięciu badanej osobie PAH. Wartości U, V i P można łatwo oznaczyć pobierając próbki moczu i krwi. i wykonując odpowiednie analizy. W ten pośredni sposób można oznaczyć przepływ osocza przez nerki. Oznaczenie objętości krwinek czerwonych (hematokrytu) pozwala na obliczenie przepływu całej krwi.
Przesączanie kłębkowe
<
Przesączanie moczu pierwotnego przez błonę kłębkową następuje pod wpływem ciśnienia hydrostatycznego panującego w naczyniach włosowatych kłębkowych. Wynosi ono u człowieka ok. 60-70 mm Hg, jest więc znacznie większe niż w naczyniach włosowatych wszystkich innych tkanek, w których sięga tylko ok. 25 mm Hg. Jest także znacznie wyższe niż ciśnienie koloidoosmotyczne białek krwi, przeciwstawiające się przesączaniu płynu z naczyń włosowatych. Ciśnienie filtracyjne w kapilarach nerkowych jest zależne od ciśnienia tętniczego. Spadek ciśnienia tętniczego krwi, prowadzący do zmniejszenia ciśnienia filtracyjnego poniżej 60-70 mm Hg, powoduje ustanie przesączania kłębkowego - nerka przestaje wytwarzać mocz, następuje skąpomocz i bezmocz.
{
Pomiar przesączania kłębkowego - ^
i
*
Oznaczanie wielkości przesączania kłębkowego jest dziś podstawowym badaniem w całej fizjologii i patologii nerek służącym do oceny ich wydolności. Oczywiście także i tego pomiaru nie można wykonać bezpośrednio, również i tu stosuje się opisane wyżej metody klirensowe. Używa się do tego wielocukru - inuliny. Inulina jest substancją, która po podaniu do krwi swobodnie przesącza się w naczyniach włosowatych kłębkowych, a w kanalikach nie ulega ani reabsorpcji ani sekrecji. Zatem ilość inuliny przesączonej w kłębusz275
i
kach w jednostce czasu (np. w ciągu minuty) równa się ilości inuliny w t y m samym czasie wydalonej z moczem. Ilość inuliny przesączonej w ciągu minuty równa się iloczynowi wielkości filtracji w ml (F) pomnożonej przez stężenie tej substancji w osoczu (P), czyli Fx P Ilość inuliny wydalonej z moczem w ciągu minuty równa się stężeniu jej w moczu (U) pomnożonemu przez objętość wydalonego w tym czasie moczu (V). Można zatem napisać równanie: Fx P= U x V skądF
=
^
Rozważania powyższe ilustruje schematycznie ryc. 139. Naczynie doprowadzające
Naczynie odprowadzające
-Pętla naczyniowa kłębuszka Cząsteczki' inuliny
Torebka kłębka
Karfalik
Ryc. 139. Schemat wyjaśniający metodę klirensowych pomiarów przesączania kłębkowego. Kuleczki wyobrażają częsteczki inuliny lub innej substancji (kreatyniny), z której krew przy jednym pasażu przez kłębek zostaje oczyszczona drogą filtracji. Dla uproszczenia przyjmujemy np. że w 1 mm osocza przepływającego znajduje się jedna taka kuleczka. Jeżeli w jednostce czasu, np. w ciągu 1 min, w moczu znajduje się 100 takich kuleczek, to oznacza to, i ż w t y m czasie przesączyło się 100 ml moczu pierwotnego.
276
N
Inulina - niskomolekularny wielocukier - nie występuje w ustroju. Dla pomiaru przesączania kłębkowego musi być wprowadzona do krwi z zewnątrz - we wstrzyknięciu lub we wlewie dożylnym. Stosując tę metodę stwierdzono, że u człowieka przesączanie kłębkowe w obu nerkach wynosi ok. 120 ml/minutę. Dla codziennej praktyki klinicznej pomiar klirensu inuliny jest zbyt kłopotliwy i obciążający pacjenta, toteż w praktyce przesączanie kłębkowe ocenia się na podstawie klirensu kreatyniny endogennej. Ma ona tę przewagę, iż jest substancją naturalnie występującą we krwi, przez co znacznie upraszcza się technika wykonania całego badania.
Reabsorpcja kanalikowa Powiedzieliśmy wyżej, iż przesączanie kłębkowe, a więc ilość moczu pierwotnego, wynosi ok. 120 ml/min, tymczasem ostateczna ilość moczu wytworzonego zwykle sięga 1-1,5 ml/min. Wynika z tego, iż większość przesączonego w kłębuszkach płynu musi ulec reabsorpcji w kanalikach nerkowych. Razem z wodą wchłonięte zostają z powrotem różne substancje w niej rozpuszczone, które są cenne dla ustroju, jak np. glukoza, aminokwasy, sole mineralne. Inne substancje, takie jak kreatynina, mocznik, oraz różne wprowadzone do ustroju substancje obce, jak np. leki podlegające wydaleniu przez nerki, nie zostają w kanalikach wchłonięte z moczu pierwotnego i przechodzą do moczu ostatecznego. W ten sposób ustrój pozbywa się niektórych związków szkodliwych lub nadmiaru substancji normalnie w płynach ustrojowych występujących, które by się mogły w zbyt dużej ilości nagromadzić. Ponadto istnieje jeszcze jeden mechanizm wydalania różnych substancji przez nerki, polegający na aktywnym wydzielaniu ich przez nabłonek kanalikowy do światła kanalików. O tym, czy dana substancja przesączona w kłębuszkach ulega wchłanianiu zwrotnemu, czy wydzielaniu czynnemu w kanalikach, można przekonać się porównując jej ilość przesączoną z ilością w tym samym czasie wydaloną z moczem. Ilość jakiejś substancji, np. glukozy, przesączonej w kłębuszkach nerkowych równa się wielkości przesączania (np. 120 ml/min) pomnożonej przez stężenie tej substancji w osoczu (np. dla glukozy = 100 mg/100 ml). W t y m przypadku 120 m l / m i n x 100 mg/100 ml = 120 mg/min. Ponieważ w warunkach prawidłowych glukoza w moczu się nie pojawia, stąd wniosek, że cała przesączona ilość tego cukru uległa reabsorpcji w kanalikach. Jeżeli ilość jakiejś substancji przesączona w kłębuszkach równa się ilości w tym samym czasie wydalonej z moczem, to możemy powiedzieć, że substancja ta ani się nie wchłania, ani nie wydziela czynnie w kanalikach. Jak już wspomniano, w ten sposób zachowuje się inulina. Jeżeli natomiast ilość jakiejś substancji wydalonej z moczem jest większa niż ilość w t y m czasie przesączona w kłębuszkach, oznacza to, iż jest ona dodatkowo wydzielana w kanalikach. Sytuacja taka zdarza się rzadziej niż reabsorpcja, ale zachowują się w ten sposób np. jony potasu, amoniaku, jony wodorowe, para-aminohipuran. '277
Z tego co powiedzieliśmy widać, że główna czynność kanalików polega na wchłanianiu zwrotnym - wchłaniają one w ciągu doby ok. 1801 wody i ok. 1130 g soli. Niektóre substancje, takie jak glukoza, wodorowęglany i chlorek sodowy, są całkowicie lub prawie całkowicie reabsorbowane w kanalikach, inne - jak mocznik i siarczany ulegają reabsorpcji w stopniu znacznie mniejszym. Wchłanianie zwrotne (reabsorpcja) większości substancji ze światła kanalików do krwi jest procesem czynnym, związanym z wydatkowaniem energii. Dla wielu substancji ulegających wchłanianiu zwrotnemu, np. dla glukozy, istnieje pewien próg - pewna maksymalna ilość, jaka może ulec reabsorpcji, gdy znajdzie się w kanalikach. Jeżeli ilość glukozy przesączonej do kanalików jest mniejsza od tego maksimum, wówczas cała glukoza zostanie zreabsorbowana. Jeżeli jednak ilość glukozy przypadającej na układ kanalikowy przewyższa możliwości transportowe nabłonka kanalików, wówczas nadmiar nie zostaje zreabsorbowany i przechodzi do moczu. Dzieje się tak np. w nie leczonej cukrzycy, gdy poziom cukru we krwi przekroczy wartość progową i ładunek przesączonej glukozy do kanalików przekracza ich maksymalną zdolność transportową. Nieco inaczej wygląda wchłanianie zwrotne sodu - ten mechanizm nie ma wyraźnie ograniczonego maksimum zdolności transportowej i im większy jest ładunek sodu w kanalikach i im dłużej on tu przebywa, tym większe jest jego wchłanianie zwrotne.
Regulacja reabsorpcji Wielkość wchłaniania zwrotnego wielu substancji jest zmienna i zależy od aktualnych potrzeb ustroju. Nerki utrzymują stały skład płynów ustrojowych i reagują na jego zmiany zmniejszonym lub zwiększonym wydalaniem pewnych substancji, przede wszystkim wody i soli mineralnych. Wydalanie tych substancji zależy oczywiście od tego, jak wielka jest ich reabsorpcja w kanalikach. Reabsorpcja ta pozostaje pod kontrolą różnych czynników - przede wszystkim niektórych hormonów. W odniesieniu do wody i sodu wygląda to np. w ten sposób, że ok. 80% wody i soli ulega reabsorpcji przymusowej w kanaliku proksymalnym, natomiast wchłanianie pozostałych 20% pozostaje pod kontrolą hormonów, zależnie od potrzeb ustroju. Tak więc hormon kory nadnerczy - aldosteron, zwiększa reabsorpcję sodu w kanalikach, a więc i zatrzymywanie soli w ustroju. Hormon antydiuretyczny tylnego płata przysadki zwiększa reabsorpcję wody, zmieniając przepuszczalność cewek zbiorczych. Reabsorpcję fosforanów reguluje hormon przytarczyc. Natomiast reabsorpcja glukozy nie jest regulowana w nerkach i dlatego nie odgrywa roli w fizjologicznej regulacji poziomu cukru we krwi.
'278
Wydzielanie Czynne wydzielanie kanalikowe następuje wówczas, gdy ilość substancji wydalonej z moczem jest większa niż jej ilość przesączona w tym samym czasie w kłębuszkach. Dla ostatecznego składu moczu mechanizm ten jest mniej istotny niż reabsorpcja kanalikowa. Do substancji wydzielanych czynnie w kanalikach, jak już wspomniano, należą potas i jony wodorowe. Wydzielanie potasu pozostaje pod kontrolą aldosteronu, zaś sekrecja jonów wodorowych zmienia się zależnie od stanu równowagi kwasowo-zasadowej ustroju.
Regulacja równowagi kwasowo-zasadowej W żywym ustroju stale powstają w procesach przemiany kwaśne produkty (kwas węglowy, siarkowy, fosforowy, mlekowy, pirogronowy itd.), a mimo to odczyn krwi (pH) i płynów tkankowych pozostaje na stałym poziomie o pH ok. 7,4. Dwutlenek węgla jest usuwany z ustroju przez płuca. Inne kwasy (nielotne) zostają najpierw zobojętnione przez zawarte w ustroju bufory, przede wszystkim przez układ buforowy składający się z wodorowęglanów i kwasu węglowego. Następuje to według zwykłej reakcji chemicznej, w której mocny kwas wypiera z wodorowęglanu dwutlenek węgla, ten zaś może z łatwośpią zostać usunięty z ustroju przez płuca: HCI + NaHC0 3 - NaCI + H 2 C0 3 H2CO3 - H 2 0 + C0 2 Gdyby kwasy w tej postaci ulegały wydalaniu, to bardzo szybko nastąpiłoby wyczerpanie wszystkich buforów i doszłoby do zakwaszenia płynów ustrojowych, uniemożliwiającego dalszy przebieg procesów życiowych. Zapobiegają temu nerki, w których kanalik jest obdarzony zdolnością usuwania nadmiaru jonów wodorowych, co jest równoznaczne z usuwaniem kwasów. W uproszczeniu przedstawia to ryc. 140. W komórkach kanalika dystalnego powstaje w wyniku procesów metabolicznych, podobnie jak w całym ustroju, dwutlenek węgla. Komórki te odznaczają się jednak tym, iż zawierają specjalny enzym - anhydrazę węglanową, która przyspiesza ok. 300-krotnie reakcję C0 2 z wodą, w wyniku czego prawie cały C0 2 zamienia się w kwas węglowy wg reakcji: H2O + C0 2 anhydraza węglanowa H 2 C0 3 Kwas węglowy ulega częściowej dysocjacji na jon w o d o r o w y i anion wodorowęglanowy: H 2 C0 3 - H+ + HCOi '279
Komórki kanalika
Mocz pierwotny
Naczynie -włosowatej
H .0+C0 • f
T
Anhydrazą -.
H_C0,
NaHCO,
H'+HC0: N«i'HC0,
•E JS j§ o •) * 2 5
<0
•o g •JO o
o C
IM
-(O o
N;i'
-H
-
H.,COn ^ Ir
•• •
N.iK.PO,
E& .H (0 ta ro
i
NH,
!
Na i-CI - .
nh4ci^-' ' I H2C03
»- NnHCO.,
;-••»• NaHCO-^
SjtBs^B: i
Glutamina
NaCl + N.i
V
/
Nu' fNdHPOi -
—NaHCO.
; VV:y-^ĘĘj^^^^ĘM |
"sj 5- 'h. h,o+co2 ł i HCO? | | j p i f f l |
Nti..HP04
Na*
1 NiiHC0 ?
-
Ę fi? '•
*»
H'-HCO:! NdHCO^
s sr NaHCO.
ł
II NaH 2 P0 4 I
»inh4ci
i
Ryc. 140. Trzy podstawowe mechanizmy zakwaszania moczu.
Uwolniony j o n w o d o r o w y zostaje wydzielony do światła kanalika nerkowego w zamian za jon sodowy (Na + ), który wchodzi do komórki i razem z anionem w o d o r o w ę g l a n o w y m NaHCCV zostaje zwrócony do krwi. W ten sposób zostaje zaoszczędzony zapas sodu ustrojowego i zapas w o d o r o w ę g l a n ó w (zasób zasad). Drugi mechanizm polega na wydalaniu jonu w o d o r o w e g o za pośrednie t w e m fosforanów. Uwolniony w sposób wyżej opisany jon w o d o r o w y w y m i ę nia się na sód, zamieniając fosforan dwusodowy na fosforan jednosodowy W ten sposób może ulec wydaleniu duża ilość j o n ó w w o d o r o w y c h (kwasu) przy równoczesnym oszczędzaniu sodu. Kwas ten daje się w moczu zmiareczkować ługiem i dlatego ta część kwasów wydalonych z moczem nosi nazwę „kwaśności miareczkowej". Wreszcie trzeci mechanizm polega na wydzielaniu przez komórki kanalikowe amoniaku. Powstaje on z dezaminacji pewnych aminokwasów. Amoniak dyfunduje do światła kanalika, gdzie przy udziale jonu w o d o r o w e g o powstałego w sposób wyżej opisany powstaje j o n a m o n o w y NH 4 + , który łączy się z chlorem uwalniając sód. Są to trzy podstawowe mechanizmy wydalania jonu w o d o r o w e g o leżące u podstaw regulacji równowagi kwasowo-zasadowej organizmu przez nerki. '280
Regulacja gospodarki wodnej Woda jest bardzo ważnym składnikiem ustroju - wszystkie reakcje biochemiczne, będące podłożem procesów życiowych, mogą odbywać się tylko w środowisku w o d n y m . Woda stanowi 65% ciężaru ciała. Rozmieszczona jest w dwóch dużych „przedziałach" - wewnątrzkomórkowym i pozakomórkowym. Na ten ostatni składa się osocze krwi i płyn śród miąższowy. W ustroju woda nie występuje nigdzie w stanie czystym, zawsze stanowi roztwór różnych soli mineralnych i cząsteczek organicznych różnej wielkości. Rozpuszczone w wodzie substancje drobnocząsteczkowe wywierają działanie osmotyczne. W skrócie m ó w i m y o osmolarności lub osmolalności płynów ustrojowych, którą określamy w miliosmolach na litr lub na kilogram roztworu. Osmolalność osocza krwi wynosi ok. 300 miliosmoli na kg. Wspomniane przedziały wodne ustroju są oddzielone od siebie błonami komórkowymi, przez które woda dyfunduje według praw fizykochemicznych w kierunku większej osmolalności, dążąc do wyrównania ciśnienia osmotycznego po obu stronach błony. Osmolalność płynów ustrojowych waha się w bardzo wąskich granicach, gdyż większe zmiany w tym zakresie powodują zaburzenie podstawowych czynności komórek i tkanek. Głównym narządem regulującym osmolalność płynów ustrojowych są nerki. Po wypiciu dużej ilości wody nerki wydzielają zwiększoną ilość bardzo rozcieńczonego moczu. Przy wstrzymaniu dowozu wody nerki wydzielą małą ilość moczu bardzo zagęszczonego. Mechanizmy zagęszczania i rozcieńczania moczu są bardzo złożone i ostatecznie jeszcze nie we wszystkich szczegółach poznane. W reakcjach rozcieńczania i zagęszczania moczu istotną rolę odgrywa hormon antydiuretyczny tylnego płata przysadki. Zmniejszenie osmolalności płynu pozakomórkowego występujące przy nadmiernym dowozie wody do ustroju powoduje zahamowanie wydzielania hormonu antydiuretycznego i wzrost objętości wydalanego moczu, natomiast zwiększona osmolalność, wskutek np. wstrzymania dowozu wody lub nadmiernego spożycia soli, powoduje zwiększoną produkcję tego hormonu, zmniejszenie objętości wydalanego moczu i zatrzymanie wody w ustroju. Miejscem działania hormonu antydiuretycznego są kanaliki zbiorcze nefronów, w których hormon ten zwiększa przepuszczalność komórek dla wody. Kanaliki kręte proksymalne i dystalne leżą w obrębie warstwy korowej nerki, natomiast pętla nefronu i kanaliki zbiorcze przylegając do siebie znajdują się w obrębie istoty rdzeniowej. Na skrawkach mikroskopowych nerki człowieka osmolalność tkanki nerkowej wzrasta w kierunku od warstwy korowej ku brodawce nerkowej od ok. 300 mOsm/kg do 1200-1400 mOsm/kg wody. W nerce zwierząt przystosowanych do warunków pustynnych, które muszą maksymalnie oszczędzać wodę i wydzielać mocz możliwie najbardziej zagęszczony, maksymalna osmolalność warstwy rdzeniowej sięga 5000 mOsm/kg wody. Przez ten obszar zwiększonej osmolalności przechodzą cewki zbiorcze. Ich ściany są przepuszczalne dla '281
)
wody w różnym stopniu, zależnie od obecności lub braku hormonu antydiuretycznego. Przy braku hormonu antydiuretycznego ściany cewek zbiorczych są „szczelne" dla wody, która w postaci rozcieńczonego moczu spływa do miedniczki nerkowej. W ten sposób ustrój pozbywa się nadmiaru wody wydalając mocz hipotoniczny w stosunku do osocza. Obecność hormonu diuretycznego powoduje, iż woda może swobodnie dyfundować przez ścianę cewki zbiorczej do obszaru zwiększonej osmolalności i w ten sposób mocz zostaje zagęszczony, a woda zatrzymana w ustroju.
Mocz (urina) Dobowa ilość moczu wydalanego przez zdrowego człowieka waha się od 600 do 2500 ml i jest zależna od różnych czynników, głównie od spożytych płynów i temperatury otoczenia. Prawidłowy mocz ma zabarwienie żółte lub bursztynowe. Intensywność zabarwienia zależy od ilości wytworzonego moczu - mocz zagęszczony jest ciemniejszy wskutek większego stężenia barwnika moczu (urochromu). Ciężar właściwy moczu zależny jest od stężenia rozpuszczonych w nim substancji i waha się od 1,003 do 1,030. Mniej więcej połowę rozpuszczonych w moczu substancji stanowi mocznik, który jest głównym produktem końcowym przemiany białkowej. Kwas moczowy jest końcowym produktem przemiany purynowej. Do produktów przemiany azotowej wydalanych z moczem należy także kreatynina, której wydalanie dobowe jest dla danego osobnika wielkością dosyć stałą. Do soli mineralnych zawartych w moczu należą sole sodu, potasu, wapnia i magnezu w postaci chlorków, fosforanów i siarczanów. Największy procent soli wydalanych z moczem stanowi chlorek sodowy (44% wszystkich soli). W warunkach chorobowych w moczu mogą pojawiać się w większych ilościach takie substancje, jak białko, cukier, barwniki żółciowe i inne. Tabela
li
Wydalanie niektórych głównych składników w moczu
'282
Składnik moczu
Wydalanie (w g na dobę)
Woda Mocznik Kwas moczowy Kreatynina Siarczany Fosforany Chlorki Amoniak Potas Sód Wapń Magnez
600-2500 33,0 0,6 1,0 2,0 1,7 7,0 0,7 2,5 6,0 0,2 0,2
Ryc. 141. Areometr (urometr) do mierzenia ciężaru właściwego moczu.
Wwarunkach prawidłowych mocz produkowany jest przez nerki w ilości 1 - 2 ml/min, ale ilość ta może zwiększać się do 20 ml/min. Mocz gromadzi się najpierw w miedniczce nerkowej, skąd spływa do pęcherza moczowego przez moczowody. MOCZOWÓD (URETEF) Jest to długi parzysty przewód łączący miedniczkę nerkową z dnem pęcherza moczowego. Długość moczowodów wynosi 28-34 cm. Biegną one zaotrzewnowo ku dołowi do miednicy mniejszej, krzyżując się z naczyniami biodrowymi wspólnymi. Moczowody przebijają ścianę pęcherza skośnie tworząc ujście zamknięte fałdem błony śluzowej. Ściana moczowodu składa się z trzech warstw: 1) łączno-tkankowej błony zewnętrznej, 2) błony mięśniowej i 3) błony śluzowej pokrytej nabłonkiem przejściowym. Mocz przechodzi z miedniczki nerkowej do pęcherza częściowo pod wpływem ciśnienia i siły grawitacyjnej, przede wszystkim jodnak dzięki perystaltycznym skurczom warstwy mięśniowej moczowodójw. Wzrost ciśnienia moczu gromadzącego się w miedniczce nerkowej wyzwala skurcze perystaltyczne moczowodu, które przesuwają mocz z miedniczki do pęcherza moczowego. Skurcze te powstają automatycznie w warstwie mięśniowej w rytmie od I do 5 na minutę, posuwając się zawsze w jednym kierunku od nerki do pęcherza. '283
Skośny przebieg moczowodu przez ścianę pęcherza powoduje, że ujście moczowodu do pęcherza pokrywa fałd błony śluzowej działający jakzastawka, uniemożliwiająca cofanie się moczu do przewodu moczowego z pęcherza. Moczowody są obficie unerwione także przez włókna bólowe. Zablokowanie moczowodu, np. przez kamień nerkowy, powoduje natężenie fali perystaltycznej przed przeszkodą wywołujące jeden z najsilniejszych rodzajów bólu, jaki charakteryzuje tzw. kolkę nerkową.
PĘCHERZ MOCZOWY ( VESICA URINARIA) Jest to worek mięśniowy położony w miednicy mniejszej za spojeniem łonowym, służący jako zbiornik moczu. W stanie opróżnionym ma wielkość małej cytryny i nie wystaje ponad spojenie łonowe. Wypełniony przybiera
Nerka
-
"Moczowód
-Pęcherz moczowy NasieniowodPęcherzyk nasienny
v
Gruczoł krokowy
Najądrze
Jądro'
Ryc. 142. Budowa układu moczowo-płciowego męskiego.
'284
kształt gruszkowaty i zależnie od pojemności wystaje do j a m y brzusznej na dość znaczną wysokość. Pojemność pęcherza bywa różna, przeciętnie w y n o s i 700 cm 3 . Ściana pęcherza moczowego składa się z trzech w a r s t w : 1) błony śluzowej, 2) błony mięśniowej i 3) błony zewnętrznej. Błona śluzowa tworzy liczne fałdy, które przy rozciąganiu pęcherza ulegają rozprostowaniu. Możliwe to jest dzięki obecności tkanki podśluzowej zbudowane} z tkanki łącznej luźnej. Na dnie pęcherza znajduje się gładka przestrzeń w kształcie trójkąta, ograniczona ujściami m o c z o w o d ó w i cewki moczowej. Błona śluzowa pokryta jest nabłonkiem przejściowym. Błona mięśniowa składa się z niezbyt wyraźnie oddzielonych trzech w a r s t w włókien mięśniowych gładkich: wewnętrznej - podłużnej, ś r o d k o w e j - o k r ę ż nej i zewnętrznej - podłużnej. U podstawy pęcherza pęczki włókien okrężnych tworzą w o k ó ł ujścia wewnętrznego cewki moczowej pierścieniowaty mięsień zwieracz pęcherza. Zwieracz ten pozostaje stale napięty, nie pozwalając na o d p ł y w moczu z pęcherza. Nieco niżej od zwieracza pęcherza znajduje się mięsień zwieracz cewki zbudowany z mięśni poprzecznie prążkowanych. Jest on unerwiony przez nerwy somatyczne i wobec tego podlega woli. Błona mięśniowa pęcherza i zwieracz pęcherza są unerwione przez włókna układu wegetatywnego. Pobudzenie n e r w ó w przywspółczulnych powoduje skurcz mięśni pęcherza i rozkurczę zwieracza pęcherza, natomiast pobudzenie włókien współczulnych wywiera efekt odwrotny - rozkurczę błony mięśniowej i skurcz zwieracza pęcherza.
ODDAWANIE MOCZU Oddawanie moczu jest odruchem sterowanym z rdzenia kręgowego. Odruch ten może być jednak świadomie zainicjowany lub zatrzymany. Potrzeba oddania moczu pojawia się zwykle, gdy pęcherz wypełni się do objętości ok. 150 ml moczu. Gdy objętość ta wzrośnie do ok. 400 ml, parcie na mocz staje się silne i dłuższe zatrzymywanie odruchu oddawania moczu jest wówczas bardzo trudne.
CEWKA MOCZOWA Cewka moczowa żeńska ( u r e t h r a feminina) jest krótkim szerokim przewodem długości 3 - 5 cm, średnicy 5 - 7 m m , który rozpoczyna się ujściem wewnętrznym w pęcherzu moczowym i kończy ujściem zewnętrznym w przedsionku pochwy. Wysłana jest od wewnątrz błoną śluzową pokrytą w części początkowej nabłonkiem przejściowym, a w dalszej - w i e l o w a r s t w o w y m walcowatym, U kobiety cewka moczowa służy wyłącznie do wyprowadzania moczu z pęcherza na zewnątrz. 19 - Anatomia i fizjologia
285
U mężczyzny cewka moczowa jest przewodem wyprowadzającym zarówno mocz, jak i spermę, w związku ż czym budowa jej przedstawia się odmiennie. Cewka moczowa męska (urethra masculina) rozpoczyna się w ujściu pęcherza moczowego i kończy ujściem zewnętrznym na żołędzi prącia. Długość jej wynosi 17,5-20 cm, jest ona zatem znacznie dłuższa niż cewka moczowa żeńska. Przy wzwodzie prącia cewka się odpowiednio wydłuża. W cewce moczowej odróżniamy trzy części: 1) część sterczową - długości ok. 3 cm, przebijającą gruczoł krokowy, 2) część błoniastą, długości ok. 2 cm, przechodzącą przez przeponę moczowo-płciową oraz 3) część gąbczastą, długości 10-15 cm, przebiegającą w ciele gąbczastym prącia. Średnica cewki moczowej męskiej nie jest jednakowa na całym jej przebiegu - najszersza jest część sterczową, najwęższa - część błoniasta. W części błoniastej włókna mięśniowe przechodzące z przepony moczowo-płciowej na cewkę moczową tworzą w jej ścianie mięsień zwieracz cewki (m. sphincter urethrae). Część gąbczasta cewki tworzy w części końcowej rozszerzenie zwane dołem ł ó d k o w a t y m ( fossa navicularis urethrae). Najwęższą część cewki stanowi jej ujście zewnętrzne. Ma ona kształt pionowej szpary. Cewkę moczową wyścieła błona śluzowa pokryta nabłonkiem w i e l o w a r s t w o w y m walcowatym.
'286
UKŁAD PŁCIOWY Zadaniem czynności płciowych jest zachowanie gatunku przez wytworzenie potomstwa. Jest to uzależnione od współdziałania osobników dwóch odrębnych płci - kobiety i mężczyzny. Na całość tych czynności składa się: 1) wytwarzanie komórek rozrodczych, czyli gamet: p l e m n i k ó w - u mężczyzny i komórek jajowych - u kobiety, 2) dokonanie aktu płciowego (kopulacji) oraz 3) regulacja hormonalna tych czynności. Przeznaczone do tych zadań narządy płciowe u mężczyzny i u kobiety są zbudowane odmiennie i dlatego musimy je omawiać oddzielnie. NARZĄDY PŁCIOWE MĘSKIE Dzielą się one na w e w n ę t r z n e i zewnętrzne. Do pierwszych zalicza się gruczoł płciowy męski produkujący plemniki - jądro, gruczoł krokowy (stercz), pęcherzyki nasienne oraz drogi wyprowadzające nasienie. Do narządów płciowych zewnętrznych należą - prącie, moszna i cewka moczowa męska.
Moczowód
Pęcherz moczowy
-Odbytnica
Gruczoł krokowy
Pęcherzyk nasienny
Prącie
•i Cewka -' moczowa Jądr<•ar*--. Ryc. 143. Narządy płciowe męskie (schemat).
'287
Narządy płciowe męskie wewnętrzne Jądro Jądro (testis, testimonium virile, orchis) jest parzystym złożonym gruczołem c e w k o w y m , w którym, jak w s p o m n i a n o wyżej, wytwarzane są gamety męskie - plemniki. Poza produkcją plemników jądro pełni rolę gruczołu dokrewnego, którego h o r m o n y regulują czynność pozostałych narządów płciowych oraz są odpowiedzialne za rozwój i utrzymanie szeregu wtórnych cech płciowych męskich. Jądro ma wielkość i kształt nieco spłaszczonego orzecha włoskiego, zawieszonego luźno w w o r k u m o s z n o w y m na powrózku nasiennym, dzięki czemu łatwo się przesuwa we wszystkich kierunkach. Od tyłu do jądra przylega najądrze oraz wchodzą do niego naczynia i nerwy, a wychodzą przewodziki odprowadzające jądra. Jądro lewe często bywa większe niż prawe i jest zawieszone niżej. Zewnętrzną warstwę jądra tworzy gruba torebka włóknista zbudowana z tkanki łącznej zbitej, zwanej błoną białawą, obejmującą miąższ jądra. Zgrubienie błony białawej wpuklające się do wnętrza jądra od tyłu zwane jest śródjądrzem. Odchodzą od niego promienisto cienkie blaszkowate przegródki jądra (septula testis), dzielące całe jądro na około 200 stożkowatych płacików (lobuli testis). Płaciki te są wypełnione przez cewki nasienne kręte ( tubuli seminiferi contorti). Na każdy płacik przypada od jednej do czterech cewek. Długość takiej cewki wynosi ok. 30-80 cm, grubość 0,1-0,3 m m . Ogólną długość cewek krętych w jądrze oblicza się na ok. 150-300 m. Z każdego płacika wychodzi jedna cewka nasienna prosta ( t u b u l u s seminiferus rectus). Cewki proste zdążają do śródjądrza, gdzie łączą się ze sobą, tworząc rodzaj sieci zwanej siecią jądra. Cewki proste są bardzo krótkie (0,2-0,4 m m ) i nieco szersze od cewek krętych. Cewki nasienne kręte są miejscem wytwarzania plemników, natomiast cewki proste stanowią pierwszy odcinek dróg odprowadzających nasienie. Cewki kręte jądra są wyłożone nabłonkiem nasieniotwórczym, w którym odróżniamy dwa rodzaje komórek: 1) plemniki i ich komórki macierzyste oraz 2) komórki podporowe (Sertolego). Na poprzecznych przekrojach cewek krętych występuje zasadniczo pięć generacji komórek płciowych, a mianowicie: 1. Spermatogonie (spermiogonie). 2. Spermatocyty I rzędu (spermiocyty). 3. Spermatocyty II rzędu (prespermidy, prespermatydy). 4. Spermatydy (spermidy). 5. Spermie, czyli plemniki. Stanowią one kolejne stadia rozwoju plemników. Najmłodsze elementy spermatogonie, znajdują się najbliżej błony podstawnej cewki, najdojrzalszew pobliżu światła cewki. '288
Spermatogeneza rozpoczyna się od podziału komórek macierzystych spermatogonii. Część spermatogonii pozostaje na miejscu przy błonie podstawnej cewki, stanowiąc rezerwę dla rozplemu następnych pokoleń komórek płciowych, pozostałe odsuwają się w kierunku światła cewki, ulegając przemianie na spermatocyty I rzędu. Podziały spermatocytów I rzędu różnią się od zwykłego podziału mitotycznego t y m , że w jego trakcie następuje zredukowanie c h r o m o s o m ó w do połowy liczby t y p o w e j dla danego gatunku, tj. do 23 u człowieka. Jest to tzw. podział redukcyjny (p. str. 10). Następny podział dojrzewania jest zbliżony do mitozy. Spermatyda ma początkowo kształt komórki nabłonkowatej, która wydłuża się, przekształcając się w plemnik. Dojrzały plemnik człowieka (spermium s. spermatozoon) składa się z główki, szyjki i witki. Główka, mająca kształt gruszkowaty, zawiera zagęszczony
'289
w małej objętości materiał jądrowy, otoczony obkurczoną błoną komórkową. Od przodu główka zawiera specjalną strukturę, tzw. akrosom. Gdy plemnik znajdzie się w bezpośrednim zetknięciu z komórką jajową, akrosom jak gdyby eksploduje, wyzwalając enzym hialuronidazę. Enzym ten prawdopodobnie otwiera drogę umożliwiając plemnikowi penetrację komórki jajowej. Witka jest narządęm kinetycznym plemnika zbudowanym podobnie jak rzęska. Zawiera ona włókienka obdarzone zdolnością kurczenia się. Skurcze ich wprawiają w ruch plemniki. W drogach rodnych żeńskich plemniki poruszają się z szybkością ok. 30 cm/h. Obok pokoleń komórek płciowych w cewkach krętych znajdują się duże komórki o kształcie smukłym, kolumnowym, rozciągające się od błony podstawnej prawie do osi cewki, tzw. komórki podporowe. Stanowią one element podporowy i odżywczy. Do każdej takiej komórki przyczepiają się liczne dojrzewające plemniki. Komórki podporowe dostarczają im prawdopodobnie materiału odżywczego, hormonów oraz enzymów niezbędnych dla procesu dojrzewania plemnika. Tkanka śródmiąższowa i wydzielanie wewnętrzne jądra Przestrzenie pomiędzy cewkami krętymi w zrazikach jądra wypełnia delikatna tkanka łączna wiotka, zwana tkanką śródmiąższową. Składa się ona z cienkich włókien kolagenowych, naczyń krwionośnych i limfatycznych, nerwów oraz kilku rodzajów komórek, wśród których najważniejsze są swoiste komórki śródmiąższowe jądra (Leydiga). Komórki te tworzą wewnątrzwydzielniczą część jądra, produkują bowiem hormony płciowe męskie, czyli androgeny. Głównym hormonem płciowym męskim jest testosteron. Androgeny wytwarzane są także w korze nadnerczy. Wydzielany przez jądra testosteron szybko (15-30 min) ulega przemianie w wątrobie i zostaje wydalony z moczem w postaci glukuronianów lub siarczanów. Pod względem chemicznym związki te są 17-ketosterydami (17-KS), ponieważ przy węglu 17 mają grupę ketonową. Wydalanie 17-ketosterydów z moczem jest wskaźnikiem sekrecji androgenów w ustroju, które u mężczyzny pochodzą z jąder i z kory nadnerczy, u kobiety - t y l k o z kory nadnerczy. Dorosły mężczyzna wydala z moczem 8-20 mg 17-KS na dobę. Dorosła kobieta - 5-15 mg. Testosteron jest odpowiedzialny za rozwój pierwotnych i wtórnych cech płciowych męskich. Niewielkie ilości testosteronu są wytwarzane w okresie rozwoju płodowego pod wpływem gonadotropiny kosmówkowej. W okresie chłopięcym do 10-12 r.ż. jądra nie produkują testosteronu prawie wcale. Wytwarzanie jego zwiększa się szybko w okresie dojrzewania płciowego i utrzymuje mniej więcej na stałym poziomie do 40 r.ż. po czym zaczyna się stopniowo zmniejszać. Wzmożona sekrecja testosteronu, jaka następuje po okresie pokwitania, powoduje powiększenie się prącia, moszny i jąder trwające do ok. 20 r.ż. '290
Testosteron powoduje także rozwój charakterystycznych dla mężczyzny wtórnych cech płciowych: włosy zarastają okolicę od spojenia łonowego w górę wzdłuż linii białej do pępka, pojawia się zarost na twarzy oraz wyrastają włosy w okolicy klatki piersiowej, rzadziej - także w innych okolicach ciała (na grzbiecie). Wzmożona produkcja testosteronu, jeśli skojarzy się z predyspozycją genetyczną, prowadzi do powstawania łysiny. Mężczyzna z nieczynnymi jądrami nie łysieje. Testosteron powoduje powiększenie się krtani i zgrubienie strun głosowych, co prowadzi do zmiany tembru głosu. Skóra pod w p ł y w e m testosteronu ulega zgrubieniu, zwiększa się w niej zawartość barwnika, wzmaga się czynność gruczołów potowych i łojowych. Trądzik pospolity (zapalenie gruczołów łojowych), będący jednym z najczęstszych objawów towarzyszących okresowi pokwitania, jest związany ze wzmożoną czynnością tych gruczołów pod wpływem testosteronu. Testosteron wywiera silny w p ł y w anaboliczny na przemianę białkową, wyrażający się dodatnim bilansem azotowym. Powoduje to duży przyrost masy mięśniowej, jaki ma miejsce bezpośrednio po okresie pokwitania. Pod wpływem testosteronu następuje silny rozrost kości i wzmożone odkładanie się w nich soli wapniowych i fosforowych. Testosteron powoduje także zrastanie się nasad kości długich z ich trzonami, co doprowadza do zahamowania wzrostu. Osobnicy wcześnie trzebieni odznaczają się wysoką smukłą sylwetką. Wydzielanie testosteronu pozostaje pod kontrolą hormonu gonadotropowego przysadki, zwanego hormonem pobudzającym komórki śródmiąższowe (ICSH). Ilość wydzielanego testosteronu jest mniej więcej proporcjonalna do ilości ICSH. Jądra nie stymulowane gonadotropiną nie produkują testosteronu. ICSH jest identyczny z LH kobiety. Prawie takie same właściwości jak ICSH ma gonadotropina kosmówkowa wytwarzana przez łożysko podczas ciąży. Hormon ten pobudza rozwój komórek śródmiąższowych i produkcję testosteronu u płodu, co ma decydujące znaczenie dla rozwoju narządów płciowych męskich. FSH (hormon pobudzający pęcherzyki) z przysadki stymuluje dojrzewanie spermatocytów w cewkach nasiennych. Bez FSH spermatogeneza nie zachodzi, jednakże dla ostatecznego wytworzenia się dojrzałych plemników konieczna jest także niewielka ilość testosteronu, a zatem także ICSH. Wytwarzanie gonadotropin przez przedni płat przysadki jest sterowane z podwzgórza. Regulacja ta odbywa się za pośrednictwem swoistych neurohormonów, zwanych czynnikami wyzwalania gonadotropin. Są to substancje polipeptydowe, których miejsce powstawania w podwzgórzu nie zostało dokładnie ustalone. Hormony te drogą układu krążenia wrotnego - podwzgórze-przysadka - zostają przeniesione do przysadki. Wydzielanie gonadotropin ulega zahamowaniu pod w p ł y w e m wstrzykiwania testosteronu. W p ł y w ten zaznacza się bardziej w stosunku do ICSH niż FSH. Pomiędzy przysadką i jądrem istnieje zatem mechanizm ujemnego sprzężenia zwrotnego utrzymujący na stałym poziomie sekrecję testosteronu - nadmiar testosteronu powoduje zahamowanie wydzielania ICSH, co z kolei zmniejsza produkcję testosteronu w jądrach.
W ciągu pierwszych dziesięciu lat życia chłopca przedni płat przysadki nie wydziela prawie wcale gonadotropin i w związku z t y m jądra nie wytwarzają androgenów. Około 10 r.ż. przysadka zaczyna produkować wzrastające ilości '291
( gonadotropin i ok. 13 r.ż. jądra osiągają pełną wydolność płciową. Poczynając od okresu pokwitania, przysadka wydziela h o r m o n y gonadotropowe przez resztę życia. Spermatogeneza, przynajmniej w postaci resztkowej, utrzymuje się zwykle do końca życia mężczyzny. U większości mężczyzn następuje jednak pomiędzy 40 i 50 r.ż. stopniowe słabnięcie czynności płciowych, związane z obniżeniem sekrecji testosteronu. W okresie t y m dochodzi do wzmożonego wydzielania FSH, ponieważ przysadka przestaje być hamowana przez testosteron. Zanik potencji płciowej przeważnie następuje bezobjawowo, niekiedy jednak w okresie t y m występują u mężczyzny zaburzenia somatyczne i psychiczne podobne do okresu przekwitania u k o b i e t y - t z w . climacterium virile.
Najądrze Najądrze ( e p i d i d y m i s ) jest układem przewodzików stanowiących magazyn plemników, w którym osiągają one pełną dojrzałość. Najądrze przylega do jądra wzdłuż jego tylnego brzegu, głównie przy g ó r n y m biegunie, tworząc t w ó r w kształcie fajki z w y g i ę t y m cybuchem. Rozróżnia się w nim głowę, trzon i ogon najądrza. Wnętrze najądrza stanowi w s p o m n i a n y już układ przewodzik ó w ; w obrębie głowy najądrza przewodziki te tworzą liczne pętle i skręty wypełniające zraziki, czyli stożki najądrza. Łącząc się ze sobą, tworzą one pojedynczy bardzo kręty i długi (4-6 m) kanał, zwany p r z e w o d e m najądrza. Przewód ten wraz z otaczającą go tkanką łączną tworzy trzon i ogon najądrza. Ogon najądrza, zaginając się ostro ku górze i ku tyłowi, przechodzi w nasieniowód. Przewodziki odprowadzające są wysłane charakterystycznym nabłonkiem, który miejscami tworzy festonowate wzniesienia składające się z wielu warstw komórek walcowatych. Na wolnej powierzchni komórek znajdują się rzęski, których ruch wytwarza prąd skierowany ku najądrzu. Pomiędzy t y m i komórkami znajdują się kubkowate zagłębienia wysłane jasnymi komórkami sześciennymi, tworzącymi tzw. gruczoły śródbłonkowe. Zewnętrzną warstwę przewodzika stanowi cienka błona podstawna oraz ułożone na niej nieliczne włókna mięśniówki gładkiej. Przewód najądrza (ductus epididymidis) ma na przekroju poprzecznym regularny zarys okrągły. Wyścieła go wysoki nabłonek walcowaty tworzący dwie warstwy. Plemniki, straciwszy kontakt z komórkami p o d p o r o w y m i , dostają się do przewodzików głowy najądrza i przewodu najądrza. Przechodzeniu ich sprzyja wzmożone ciśnienie panujące w jądrze. W najądrzu plemniki osiągają pełną dojrzałość. Plemniki przebywające w drogach najądrza zachowują zdolność zapłodnienia do 42 dni, chociaż przy normalnej aktywności płciowej rzadko okres ten bywa tak długi. Przy często powtarzanych stosunkach płciowych zapasy dojrzałych plemników ulegają szybko wyczerpaniu i już trzeci ejakulat w ciągu 12 h jest praktycznie pozbawiony plemników. Ponowne wypełnienie '292
zbiorników trwa około dwóch dni, a okres ich przebywania w najądrzu w tych warunkach może się ograniczyć do kilku godzin. Liczba plemników w jednym ejakulacie w y n o s i ok. 200-300 milionów.
Nasieniowód Nasieniowód ( ductus deferens) stanowi bezpośrednie przedłużenie przew o d u najądrza. Jest to przewód długości 50-60 cm służący do szybkiego przeprowadzania nasienia do cewki moczowej. Biegnie on początkowo po przyśrodkowej stronie jądra, kierując się ku kanałowi pachwinowemu, przez który wchodzi do wnętrza miednicy małej w powrózku nasiennym. W miednicy małej nasieniowód zdąża ku tylnej ścianie pęcherza moczowego. Po skrzyżowaniu się z moczowodem zbliża się do nasieniowodu strony przeciwnej, przylegając do dna pęcherza. Ściana nasieniowodu jest gruba i składa się z trzech błon: zewnętrznej zbudowanej z tkanki łącznej włóknistej, zawierającej liczne włókna sprężyste, środkowej - błony mięśniowej, oraz w e w n ę t r z n e j - błony śluzowej. Najgrubsza (ok. 1 m m ) jest warstwa mięśniowa; dzięki niej nasieniowód daje się wyraźnie wyczuć w miejscu, gdzie leży powierzchownie pod skórą, tj. przed wejściem do kanału pachwinowego. Średnica nasieniowodu w y n o s i ok. 3 m m , światło - 0,5 m m . W końcowej części nasieniowód tworzy wrzecionowate rozszerzenie, zwane bańką nasieniowodu ( a m p u l l a ductus deferentis). Wydzielina bańki nasieniow o d u pobudza plemniki do ruchu. Następnym odcinkiem dróg wyprowadzających nasienie jest krótki, 20-milimetrowy, przewód w y t r y s k o w y ( ductus ejaculatorius), który przebija gruczoł krokowy i otwiera się m a ł y m ujściem do części sterczowej cewki moczowej. Ujścia przewodów wytryskowych znajdują się po obu stronach łagiewki sterczowej. Pęcherzyki nasienne ( vesiculae seminales) - prawy i lewy - stanowią wypuklenie nasieniowodu w kształcie gruszkowatego płaskiego t w o r u długości ok. 4 - 5 cm. Są to gruczoły pomocnicze, wytwarzające zasadową wydzielinę, która zawiera białko zasadowe (protaminy), wielocukry oraz fruktozę (ok. 300 mg%). Ta ostatnia stanowi materiał odżywczy i źródło energii dla plemników. Wydzielina pęcherzyków nasiennych jest żółtawym gęstym płynem o odczynie zasadowym, który pobudza plemniki do ruchu. Usunięcie pęcherzyków nasiennych przeważnie pociąga za sobą niepłodność.
Gruczoł k r o k o w y Gruczoł k r o k o w y , stercz (prostata), jest nieparzystym narządem wielkości kasztana, położonym w jamie miednicy mniejszej pod pęcherzem m o c z o w y m za spojeniem ł o n o w y m . Ma on kształt spłaszczonego kasztana, podstawą '293
V zwróconego ku górze. Przeciętne rozmiary gruczołu krokowego wynoszą: w wymiarze strzałkowym - 2 cm, wysokość ok. 3 cm, szerokość - ok. 4 cm. Przednią część gruczołu krokowego przebija cewka moczowa; przez część tylną biegną skośnie ku dołowi i przodowi przewody wytryskowe. Gruczołten w wieku starczym ulega często powiększeniu i uciskając cewkę moczową stwarza przeszkodę w oddawaniu moczu. Od zewnątrz gruczoł krokowy otacza torebka łącznotkankowa zawierająca także włókna mięśniowe. Wydzielina gruczołu krokowego stanowi około 20% płynnej części nasienia. Zawiera ona m.in. kwas cytrynowy, substancje białkowe oraz szereg enzym ó w , jakfosfataza kwaśna, beta-glukurohidaza i fibrynolizyna. Nasienie, które bardzo szybko po wytrysku krzepnie, pozostawione w temperaturze ok. 37° ulega pod w p ł y w e m fibrynolizyny upłynnieniu. Gruczoł krokowy wydziela także substancje o właściwościach hormonalnych, tzw. prostaglandyny, które po dostaniu się do pochwy powodują zwolnienie skurczów macicy i rozluźnienie końca brzusznego jajowodu.
Gruczoły o p u s z k o w o - c e w k o w e Gruczoły opuszkowo-cewkowe ( g l a n d u l a e bulbourethrales) przeważnie mają kształt i wielkośćziarna grochu, barwężółtawobrązową i leżą przy t y l n y m końcu opuszki prącia na dnie miednicy mniejszej. Gruczoły te produkują jasną, podobną do śluzu, wydzielinę o odczynie lekko zasadowym. Substancja ta wydziela się obficie podczas pobudzenia płciowego; gruczoły opuszkowo-cewkowe opróżniają się już podczas wzwodu prącia przygotowując drogę dla nasienia.
Powrózek nasienny Powrózek nasienny (funiculus spermaticus) składa się z nasieniowodu, naczyń krwionośnych i splotów nerwowych oraz osłonek takich samych, jakie otaczają jądro, z wyjątkiem blaszek surowiczych. Długość powrózka nasiennego w y n o s i ok. 15-20 cm, średnica dorównuje grubości małego palca. Powrózek nasienny daje się wymacać przez skórę moszny oraz w dole p a c h w i n o w y m przy wejściu do kanału pachwinowego.
Przewody w y t r y s k o w e Następny odcinek drogi nasienia od bańki nasieniowodu do ujścia cewki moczowej, położony w obrębie gruczołu krokowego, nosi nazwę przewodu w y t r y s k o w e g o (ductus ejaculatorius). Długość jego w y n o s i ok. 2 cm, światło od ok. 1 mm w części początkowej, zwężając się do 0,2 mm przy ujściu cewki 294
moczowej. Obydwa przewody wytryskowe uchodzą do cewki moczowej, każdy na m a ł y m wzniesieniu błony śluzowej, zwany w z g ó r k i e m nasiennym (colliculus seminalis), po obu stronach niewielkiego podłużnego zagłębienia łągiewki sterczowej (utriculus prostaticuś).
Narządy płciowe męskie zewnętrzne Prącie Prącie, członek męski (pen/s, membrum virile) jest narządem o p o d w ó j n y m zadaniu - służy do wprowadzenia nasienia do dróg rodnych kobiety w trakcie aktu płciowego oraz do wyprowadzania moczu z pęcherza na zewnątrz. Czynnościom t y m odpowiada jego budowa. Prącie ma kształt podłużnego stożkowato zakończonego walca. Część tylna prącia (pars fixa) ukryta pod skórą moszny i krocza jest nieruchomo przytwierdzona do kości łonowych i spojenia łonowego. Trzon prącia (corpus penis) wraz z żołędzią (glans penis) w stanie spoczynku zwisa nad workiem mosznowym jako tzw. część wisząca (pars pendula penis). Podczas wzwodu prącie usztywnia się i powiększa bardzo znacznie swe w y m i a r y . Prącie w stanie w z w o d u , czyli phallus, przybiera kierunek zwrócony ku górze i ku przodowi. W stanie zwiotczałym długość części ruchomej prącia wynosi 8,5-10,5 cm, przeciętnie - 9,5 cm. W stanie w z w o d u trzon prącia wydłuża się do ok. 15-17 cm, a o b w ó d - do ok. 12 cm. Trzon prącia zakończony jest stożkowatym zgrubieniem zwanym żołędzią (glans penis). Na wierzchołku żołędzi znajduje się szczelinowate ujście zewnętrzne cewki moczowej. Na granicy trzonu z żołędzią widnieje wyraźne rynienkowate zwężenie - szyjka żołędzi (collum glandis). Zgrubiały brzeg podstawy żołędzi tworzy wypukły pierścień zwany koroną żołędzi (corona glandis). Skóra pokrywająca trzon prącia jest bardzo cienka, łatwo przesuwalna; z podłożem łączy się ściśle wzdłuż korony żołędzi. Na granicy z żołędzią w stanie zwiotczałym skóra tworzy głęboki fałd pokrywający żołądź zwany napletkiem (praeputium). Napletek stanowi zapasowy materiał skórny, który przy powiększeniu się prącia podczas w z w o d u cofa się ku t y ł o w i obnażając całkowicie żołądź. Niekiedy ujście napletka bywa tak wąskie, iż nie może się cofnąć do tyłu po wzwodzie tworząc tzw. stulejkę (phimosiś). W kieszonce napletka gromadzi się mazista t r e ś ć - m a s t k a (smegma praeputii). Stulejka sprzyja gromadzeniu się mastki i zanieczyszczeń stwarzając warunki sprzyjające stanom zapalnym. Szczególnie duże ilości mastki gromadzą się w kieszonce napletka u mężczyzn nie przestrzegających higieny osobistej. Jeśli się jej nie usuwa regularnie, powstają kwasy tłuszczowe wydzielające w o ń zjełczałego tłuszczu. Napletek nie tworzy pełnej kieszonki na całym obwodzie żołędzi. Od dołu od blaszki wewnętrznej fałdu skórnego odchodzi tzw. wędzidełko napletka uniemożliwiające nadmierne jego cofanie się. Trzon prącia jest utworzony z trzech cylindrycznych t w o r ó w , zwanych ciałami jamistymi. Dwa ciała jamiste prącia (corpora cavernosa penis) leżą po '295
\ Ujście zewnętrzne cewki Wędzidełko napletka ik1L_Żołądź §| prącia Korona żołędzi -Trzon prącia Ciało jamiste ,
m
Ciało
" Cjdlicz.isll! pian.i
K. kulszowa
M. kulszowo-jamisty- - y—
Korzeń prącia
_—-M. poprzeczny & głęboki fc krocza '
M. opuszkowo-jamisty
^
1
Ryc. 145. Ciała jamiste prącia i ich mięśnie (po zdjęciu skóry).
stronie grzbietowej (górnej) prącia, zaś nieparzyste- - ciało gąbczaste prącia (corpus spongiosum penis) przylega do nich od dołu leżąc w płytkim rowku. Każde z ciał jamistych prącia ma kształt walca o zaostrzonym końcu. Końce tylne, zwane odnogami prącia (crura penis), rozchodząc się do tyłu i od dolnego punktu spojenia łonowego, ku tyłowi, dołowi i bocznie przytwierdzają się do kości kulszowej i łonowej w ten sposób, że ich łącznotkankowa pokrywa zrasta się z okostną. Ciała jamiste prącia są zrośnięte ze sobą w linii środkowej, tworząc przegrodę prącia (septum penis). Przegroda ta nie jest szczelna, lecz posiada liczne otwory łączące ze sobą komory obu ciał jamistych. Na przedni koniec zrośniętych ciał jamistych nasadzona jest na kształt czepka lub naparstka żołądź. Podczas wzwodu prącia koniec ten tworzy dla żołędzi mocne oparcie. Ciało gąbczaste prącia (corpus spongiosum penis) obejmuje cewkę moczową. Jest ono dłuższe i cieńsze niż ciała jamiste prącia. Na obu swych końcach, rozszerza się tworząc z przodu wspomniane już zgrubienie w kształcie kołpaczka nasadzonego na koniec przedni ciał jamistych prącia zwane żołędzią, zaś '296
w końcu tylnym - zgrubienie zwane opuszką prącia (bulbus penis). Opuszka wielkości orzecha laskowego leży w okolicy krocza na przeponie moczowo-płciowej pokryta mięśniem opuszkowo-gąbczastym. Skurcz tego mięśnia powoduje wyrzucenie nasienia podczas wytrysku z cewki moczowej. Cewka moczowa wchodzi do ciała gąbczastego mniej więcej w odległości 1 cm ku przodowi od tylnego końca opuszki. Każde z ciał jamistych zbudowane jest z bardzo mocnej osłonki łącznotkankowej z tkanki łącznej zbitej, grubości ok. 1 mm, zwanej błoną białawą (tunica albuginea). Tkanka jamista stanowi układ nieregularnych przestrzeni naczyniowych jamek, pomiędzy licznymi beleczkami odchodzącymi do wewnętrznej powierzchni błony białawej. Układ przestrzeni naczyniowych otrzymuje krew z końcowych odgałęzień odchodzących od tętnicy głębokiej prącia i w mniejszym stopniu z tętnicy grzbietowej prącia. Ciało gąbczaste prącia jest zbudowane podobnie jak ciała jamiste. W z w ó d prącia Wzwód prącia następuje wskutek wypełnienia ciał jamistych i ciała gąbczastego krwią tętniczą pod dużym ciśnieniem. Ośrodek nerwowy wzwodu znajduje się w odcinku krzyżowym rdzenia kręgowego. Wychodzące stąd impulsy przywspółczulne dochodzą do prącia przez nn. erigentes powodując rozkurcz wszystkich mm. gładkich w tętnicach i beleczkach ciał jamistych, zwiększając w ten sposób wielokrotnie napływ do prącia krwi, która wypełnia zatoki ciał jamistych i ciała gąbczastego. Większe żyły prącia są zaopatrzone w lejkowate zastawki, które utrudniają odpływ krwi żylnej. Przestrzenie jamiste pod wpływem napływającej pod dużym ciśnieniem krwi ulegają rozciągnięciu powodując napięcie błony białawej. Ciśnienie w przestrzeniach naczyniowych ciał jamistych jest zbliżone do ciśnienia w układzie tętniczym. Wynikiem tego jest znaczne powiększenie i usztywnienie prącia, umożliwiające dokonanie aktu płciowego. Wzwód prącia u seksualnie wydolnego młodego mężczyzny następuje w ciągu 3 - 8 s. w odpowiedzi na efektywne pobudzenie płciowe. U mężczyzn starszych (po 40 r.ż.) wzwód dochodzi do skutku nieco wolniej. Podczas wzwodu prącia przestrzenie jamiste ciała gąbczastego są wypełnione krwią, podobnie jak w ciałach jamistych, jednakże odpływ krwi nie jest tu zahamowany w takim stopniu i nawet przy najsilniejszym wzwodzie ciało gąbczaste poddaje się uciskowi, a światło cewki moczowej nie ulega zamknięciu.
Moszna Jądra leżą w odzielnym worku - mosznie (scrotum), utworzonym przez wypuklenie przedniej ściany jamy brzusznej. W okresie płodowym zawiązki gruczołów płciowych powstają na tylnej ścianie jamy brzusznej na wysokości '297
\ dolnych kręgów lędźwiowych. W trakcie rozwoju płodowego jądra odbywają wędrówkę zwaną „zstępowaniem jąder" (descensus testiculorum) z jamy brzusznej przez kanał pachwinowy do moszny. W procesie tym uczestniczy także przednia ściana jamy brzusznej, tworząc jak gdyby kieszonkę ze wszystkich swoich warstw z otrzewną włącznie, wędrującą przed jądrem do worka mosznowego. Jądro ma zatem zawczasu przygotowaną drogę i nie powoduje wypuklenia ściany brzusznej, lecz rzeczywiście zstępuje do gotowego worka mosznowego. Proces zstępowania jąder zostaje zakończony przed urodzeniem i jama surowicza moszny traci połączenie z jamą otrzewnej. Jeżeli połączenie takie zachowa się, wówczas powstaje wrodzona przepuklina pachwinowa, przez którą mogą wchodzić do moszny pętle jelitowe. Zatrzymanie się jądra w trakcie zstępowania w jamie brzusznej lub w kanale pachwinowym jest określone jako w n ę t r o s t w o (cryptorchismus). W środku worek mosznowy jest przedzielony przegrodą (septum scroti) na dwie jamy. Skóra moszny odznacza się pigmentacją silniejszą, małą zawartością tkanki tłuszczowej oraz obecnością szczególnej warstwy mięśni gładkich tworzących tzw. błonę kurczliwą (tunica dartos). Dzięki owej błonie skóra moszny może się silnie rozciągać dostarczając części pokrywy dla prącia przy jego wzwodzie. Brak podściółki tłuszczowej ułatwia wymianę ciepła i temperatura w worku mosznowym jest o 2,5-4° niższa niż w jamie brzusznej.
NARZĄDY PŁCIOWE ŻEŃSKIE Czynności płciowe żeńskie są bezpośrednio związane z rozmnażaniem i dlatego można je podzielić na dwie zasadnicze fazy: pierwsza polega na przygotowaniu ustroju kobiety do zapłodnienia i c i ą ż y - d r u g a - obejmuje okres samej ciąży. Narządy płciowe żeńskie dzielimy na wewnętrzne i zewnętrzne. Do wewnętrznych należą: jajniki, jajowody, macica i pochwa. Do zewnętrznych zalicza się srom niewieści.
Narządy płciowe żeńskie wewnętrzne Jajnik Jajnik (ovarium), czyli gruczoł płciowy żeński, jest odpowiednikiem jądra u mężczyzny. Jest to twór parzysty wielkości małego orzecha włoskiego, barwy białej lub lekko różowej o matowej powierzchni, konsystencji twardej, sprężystej, bardzo wrażliwy na ucisk. Jajnik leży w miednicy mniejszej, gdzie jest umocowany w więzadle szerokim macicy za pośrednictwem bardzo krótkiej krezki (mesovarium). Ponadto w umocowaniu jajnika bierze udział więzadło wieszadłowe jajnika (ligamentum suspensorium ovarii) oraz więzadło właściwe jajnika (ligamen'298
u
Pęcher moczo' Przepon moczow -płciow
Ryc, 146. Budowa narządów płciowych żeńskich.
tum ovarii proprium). Wewnątrz więzadła wieszadłowego biegną do jajnika naczynia i nerwy idące do wnęki jajnika od ściany miednicy. Wspomniane więzadło jajnika właściwe łączy biegun dolny narządu z ujściem macicznym jajowodu. Zawiera ono włókna sprężyste oraz pasma mięśni gładkich, dzięki czemu może się skracać i wydłużać. Pomimo wymienionych wyżej więzadeł jajnik jest narządem ruchomym, mogącym zmieniać swe położenie w dosyć szerokim zakresie. W odróżnieniu od jądra, w którym pasma nabłonka płciowego wrastają do wnętrza powstającego jądra, przekształcając się w cewki nasienne, w jajniku nabłonek płciowy pozostaje na powierzchni. Pod nim znajduje się warstwa łącznotkankowa tworząca delikatną błonę białawą (tunica albuginea), która jest jednak znacznie słabiej rozwinięta niż analogiczna błona w jądrze. Pod nią leży warstwa tkanki łącznej luźnej, która tworzy tu zrąb jajnika (stroma ovarii). W zrębie jajnika znajdują się pęcherzyki jajnikowe. Jajnik noworodka żeńskiego zawiera bardzo liczne pęcherzyki jajnikowe pierwotne (folliculi ovarii primarii). Każdy taki pęcherzyk składa się z dużej centralnie położonej komórki jajowej - oogonii, otoczonej pojedynczą warstwą komórek nabłonka pęcherzykowego. Komórki te w stosunku do jaja odgrywają rolę podobną jak komórki podporowe dla plemników, tj. biorą udział w jego odżywianiu. Wiele z tych pęcherzyków pierwotnych ulega w okresie dzieciństwa i pokwitania zanikowi. Ogólną liczbę pęcherzyków pierwotnych szacuje się na około 200 000. W okresie płodności kobiety, tj. od pokwitania do menopauzy, dojrzewa z tej liczby tylko około 400, a praktycznie szansę spełnienia swej misji biologicznej, tj. przekształcenia się w organizm potomny, ma tylko około 10 komórek jajowych. W zestawieniu z liczbą plemników w ejakulacie, z których szansę '299
\ spełnienia misji biologicznej ma tylko 1 spośród 200-300 milionów widać jak zapobiegliwie rozrzutna jest natura, gdy idzie o sprawę zachowania gatunku. Z powyższegq wynika także, że wiek komórek jajowych wydalanych z jajnika waha się zatem od 14 do ok. 45 lat. Być może w związku z tym pozostaje większa ilość wad wrodzonych u dzieci starszych matek. Po uzyskaniu dojrzałości płciowej, mniej więcej co 28 dni pewna ilość pęcherzyków (przypuszczalnie około 20) zaczyna ulegać zmianom, doprowadzającym do powstania tzw. pęcherzyków wtórnych. Płaskie komórki pęcherzykowe otaczające komórkę jajową zaczynają się rozmnażać i przekształcać w komórki sześcienne i walcowate tworząc wiele warstw. Powoduje to powiększenie się całego pęcherzyka. Stosunkowo niewielka liczba pęcherzyków wtórnych przekształca się w pęcherzyki jajnikowe dojrzewające. Wtrakcie procesu dojrzewania pęcherzyka komórki pęcherzykowe zaczynają wytwarzać płyn pęcherzykowy (liquor folliculi), który gromadzi się w jedną powiększającą się kroplę i rozsuwa komórki tworząc jamę pęcherzyka (antrum folliculi). W ten sposób pęcherzyk zostaje rozdzielony na dwie częścr- część odsuniętą
Największa możliwość zapłodnienia
Ryc. 147. Schemat cyklu miesiączkowego.
300
1
od jaja i część związaną z jajem. Część komórek skupionych wokół komórki jajowej tworzy tzw. wzgórek jajonośny (cumulus oophorus) wpuklający się do wnętrza pęcherzyka. Rozwój pęcherzyka pozostaje pod kontrolą hormonu gonadotropowego przysadki folikulostymuliny (FSH). Całkowicie rozwinięty pęcherzyk ma ok. 10 mm średnicy. Gotowy do pęknięcia pęcherzyk położony tuż pod powierzchnią jajnika osiąga jeszcze większe rozmiary. Równocześnie rośnie także komórka jajowa, która osiąga średnicę ok. 0,2 m m , stając się dostrzegalna dla gołego oka. Jest to oocyt I rzędu. W dniu poprzedzającym pękanie pęcherzyka oocyt I rzędu dzieli się nierównomiernie wydalając pierwsze ciałko kierunkowe. W wyniku tego podziału powstaje oocyt II rzędu o zredukowanej do połowy liczbie chromosomów. Gdy dojrzały pęcherzyk pęka, wydalony zostaje oocyt II rzędu. Dalszy podział dojrzewania i wydalenie drugiego ciałka kierunkowego następuje w chwili zapłodnienia. Oocyt II rzędu otoczony wieńcem komórek pęcherzykowych wydostaje się z pękniętego pęcherzyka do jamy otrzewnej, skąd zostaje przejęty przez strzępki jajowodowe i przeprowadzony do jajowodu. W każdym cyklu miesięcznym z reguły dojrzewa i pęka jeden pęcherzyk jajnikowy (Graafa). Proces ten nosi nazwę jajeczkowania, czyli owulacji. Owulacja następuje zasadniczo w środku cyklu (intermenstruum), zwykle pomiędzy 13 i 16 dniem cyklu miesięcznego. W cyklach nieprawidłowo długich lub krótkich wahaniom ulega okres przedowulacyjny, natomiast okres pomiędzy owulacją a dniem wystąpienia krwawienia miesięcznego przeważnie jest stały i zazwyczaj wynosi 14-16 dni. Ponieważ zdolność jaja do zapłodnienia ocenia się na 24 do 48 h od chwili owulacji, to zapłodnienie może nastąpić tylko w jajowodzie, gdyż wędrówka jaja do macicy trwa około 72 h. Wynika z tego, że okres, w którym jajo może ulec zapłodnieniu, jest ograniczony i wynosi 24-36 h. Nie zapłodnione jajo ulega zwyrodnieniu i wydaleniu podczas miesiączkowania. Przyjmuje się, że plemniki zachowują zdolność zapłodnienia w ciągu 18-24 h, ponieważ jednak żywe plemniki znajdowano w drogach rodnych kobiety do trzech dni, to okres, w którym jajo może ulec zapłodnieniu, rozciąga się na ok. 4 dni. Zapłodnienie może zatem nastąpić, jeżeli plemniki zostaną złożone w drogach rodnych na 1 - 2 dni przed owulacją. Największa możliwość zapłodnienia przypada więc na okres około 4 dni w środku cyklu miesiączkowego. W niektórych cyklach owulacja może nie dochodzić do skutku w zwykłym terminie. Nie należy ignorować doniesień opisujących przypadki owulacji i zapłodnienia w niezwykłych okresach cyklu, nawet podczas miesiączki. Po owulacji ściana pęcherzyka zapada się i ulega pofałdowaniu. Do jamy pęcherzyka następuje niewielkie krwawienie. Otwór wielkości łebka od szpilki, pozostający po pęknięciu pęcherzyka, zostaje zaczopowany przez"skrzep. Komórki warstwy ziarnistej zaczynają się intensywnie rozmnażać i powiększać. W ich cytoplazmie gromadzi się żółty barwnik, dzięki czemu pierwotnie '301
\ szaroróżowy pęcherzyk przeistacza się w twór o zabarwieniu żółtawym, noszący teraz nazwę ciałka żółtego (corpus luteum). Jeżeli nie nastąpi zapłodnienie, okres czynności ciałka żółtego trwa ok. 12-14 dni, po czym zaczyna ono zanikać. Jest to tzw. ciałko żółte miesiączkowe. Komórki luteinowe ulegają zwyrodnieniu tłuszczowemu, ich miejsce zajmuje tkanka łączna, a cały twór przekształca się w tzw. ciałko białawe (corpus albicans), które z czasem zanika i po upływie ok. 2 miesięcy pozostaje na jego miejscu tylko mała włóknista blizna. Jeżeli nastąpi zapłodnienie i implantacja jaja, to pod wpływem hormonów produkowanych przez trofoblast rozwija się ciałko żółte ciążowe, które powiększa się, osiągając około 4 miesiąca ciąży średnicę do 5 cm. Od 6 miesiąca ciałko żółte zaczyna powoli zanikać. Hormony płciowe żeńskie Prawidłowe życie płciowe kobiety podlega cyklicznym zmianom sterowanym z podwzgórza za pośrednictwem przysadki. Ośrodki w podwzgórzu wytwarzają dwie substancje polipeptydowe o charakterze neurohormonów, zwane czynnikiem uwalniającym, oznaczone pierwszymi literami angielskiej ich nazwy-FRF (Follicle Stimulating Hormone Releasing Factor) oraz LRF (Luteinizing Hormone Releasing Factor), które trafiają do przedniego płata przysadki. Cykliczny charakter wyzwalania tych hormonów jest specyficzną cechą mózgu żeńskiego. U mężczyzny wydzielanie hormonów gonadotropowych ma charakter mniej więcej stały. Obecność w ustroju żeńskim swoistego „zegara biologicznego" zlokalizowanego w podwzgórzu jest prawdopodobnie zależna od tego, że w chwili narodzenia nie ma w nim męskich hormonów płciowych. Wstrzykiwanie noworodkowi żeńskiemu testosteronu zapobiega powstawaniu ośrodka cyklicznego i wyzwalanie hormonów gonadotropowych u takich osobników odbywa się w sposób ciągły bez cyklicznych fluktuacji. Za pośrednictwem podwzgórza i przedniego płata przysadki ośrodkowy układ nerwowy może wywierać wpływ na aktywność gonad. Wiadomo np., że czynniki emocjonalne i środowiskowe mogą zaburzać i zmieniać rytm miesiączkowy. Cykl jajnikowy zależy całkowicie do hormonów gonadotropowych przysadki. Jajniki, które nie są stymulowane przez te hormony, jak to jest np. w dzieciństwie, pozostają nieczynne. Wycięcie przedniego płata przysadki u kobiety dojrzałej prowadzi do zaniku czynności jajników. Przedni płat przysadki wydziela trzy różne hormony gonadotropowe istotne dla czynności jajnika. Są to: 1) hormon pobudzający pęcherzyki (follikulostymulina)-FSH, 2) hormon luteinizujący (LH) oraz 3) luteotropina (LTH). FSH i LH działają wyłącznie na jajnik, natomiast LTH poza wpływem na jajniki pobudza także gruczoły sutkowe do wydzielania mleka i dlatego bywa nazywany prolaktynę (PRL) lub hormonem laktogennym. Sama prolaktyna ma niewielki wpływ na gruczoł sutkowy; dla przygotowania gruczołu do pełnej laktacji (rozrostu przewodów i pęcherzyków mlecznych) potrzebne jest jeszcze współdziałanie estrogenów i progesteronu, a także hormonów korowo-nadnerczowych. LH u mężczyzny bywa nazywany hormonem pobudzającym komórki śródmiąższowe (ICSH). Na początku cyklu zaznacza się niewielki wzrost poziomu FSH we krwi powodujący dojrzewanie pęcherzyków jajnikowych oraz stymulujący komórki pęcherzykowe do wydzielania estrogenów. Z osocza krwi wyizolowano w większych ilościach trzy naturalne estrogeny: beta-estradiol, estron i estriol. Wzrost poziomu estrogenów we krwi jest odpowiedzialny za fazę wzrastania błony śluzowej macicy w pierwszej połowie cyklu miesięcznego. Gdy poziom estrogenów we krwi zaczyna spadać, narasta szybko wydzielanie LH. Wzmożone wydzielanie LH przypuszczalnie powoduje pękanie dojrzałego pęcherzyka jajnikowego i rozwój ciałka żółtego, które wytwarza progesteron. W drugiej połowie cyklu jajnikowego prawie cały progesteron pochodzi z jajnika. Podczas ciąży, zwłaszcza po czwartym miesiącu ciąży, progesteron jest wytwarzany w dużych ilościach w łożysku.
'302
Przysadka
Ryc. 148. Schemat regulacji wydzielania hormonów płciowych.
Progesteron przyczynia się do zmian w błonie śluzowej macicy, przygotowując ją do implantacji zapłodnionego jaja. Progesteron zmniejsza także częstość skurczów macicy. Progesteron jest hormonem sterydowym, który w wątrobie bardzo szybko ulega przemianie na związki pozbawione właściwości progestacyjnych. Wytwarzanie estrogenów zwiększa się podczas pokwitania ponad dwudziestokrotnie. Pod ich w p ł y w e m następuje przekształcanie się narządów płciowych z f o r m y dziecięcej w postać charakterystyczną dla dojrzałej kobiety oraz rozwijają się wtórne cechy płciowe żeńskie. Następuje powiększenie macicy, jajowodów i pochwy, rozwija się podściółka tłuszczowa w obrębie wzgórka łonowego i warg sromowych większych, powiększają się wargi sromowe mniejsze. Nabłonek
'303
pochwy z sześciennego zmienia się na wielowarstwowy płaski, znacznie bardziej odporny na zakażenia i urazy. W macicy rozpoczynają się cykliczne zmiany w błonie śluzowej polegające na przeroście i rozwoju gn iczołów. W jajowodach zwiększa się liczba komórek rzęskowych i wzmaga ruch rzęsek wytwarzających prąd w kierunku światła macicy, co sprzyja przemieszczaniu się zapłodnionego jaja do jamy macicy. Estrogeny powodują rozwój tkanki łącznej i zrębu sutka, nadając mu charakterystyczny wygląd zewnętrzny. Inicjują one także wzrost pęcherzyków i zrazików gruczołowych, nie doprowadzając jednak do przemiany sutka w narząd produkujący mleko - odpowiedzialny za rozwój elementów gruczołowych sutka jest bowiem progesteron, natomiast estrogeny nawet hamują produkcję mleka. Estrogeny przyśpieszają wzrost kości, dzięki czemu w okresie pokwitania zaznacza się u dziewcząt wzmożone wzrastanie. Doprowadzają one jednak także do zrastania się nasad z trzonami kości długich i w konsekwencji - zatrzymania dalszego wzrostu. W p ł y w ten jest silniejszy niż analogiczny w p ł y w testosteronu w organizmie męskim, toteż kobiety pozbawione jajników przed okresem pokwitania osiągają wzrost o kilka centymetrów większy niż trzebieńcy męscy. Ubocznym w p ł y w e m estrogenów na rozwój kośćca jest charakterystyczne ukształtowanie się żeńskiego typu miednicy. Podobnie jaktestosteron, estrogeny zwiększają zatrzymywanie wapnia ifosforu oraz wywierają słaby w p ł y w anaboliczny w przemianie białkowej. Estrogeny zwiększają ukrwienie skóry, dzięki czemu skóra kobiety jest cieplejsza niż u mężczyzny. Z tej samej przyczyny krwawienia z powierzchownych ran ciętych skóry u kobiety bywają większe. Estrogeny nadają skórze kobiety charakterystyczną miękkość i gładkość. Dzięki budowie chemicznej zbliżonej do sterydów nadnerczowych estrogeny zwiększają wchłanianie zwrotne sodu i w o d y w cewkach dystalnych nefronu, powodując nieznaczną retencję soli i wody. Hormon ciałka żółtego - progesteron, poza działaniem na błonę śluzową macicy i jajowodów, wywiera także w p ł y w na rozwój gruczołów sutkowych, przygotowując je do aktywnego wytwarzania mleka, które jednak następuje dopiero pod w p ł y w e m pobudzenia prolaktynę z przysadki. Progesteron zwiększa także wchłanianie zwrotne sodu, chloru i w o d y w nerkach, prowadząc do zatrzymania tych substancji w ustroju. Progesteron wywiera niewielki w p ł y w kataboliczny na białka, co prawdopodobnie ma znaczenie w okresie ciąży, gdy następuje uruchamianie zasobów białkowych matki na potrzeby rosnącego płodu. W przebiegu prawidłowego cyklu jajnikowego w p ł y w ten jest znikomy.
Jajowód Jajowody (tuba uterina, salpinx), w liczbie dwóch (prawy i lewy), są to cewkowate przewody długości 14-20 cm służące do przyjęcia i przeprowadzenia komórki jajowej z jajnika do jamy macicy. Każdy jajowód ciągnie się od końca jajowodowego jajnika do górnego kąta macicy, biegnąc w górnym brzegu więzadła szerokiego macicy pod otrzewną. Koniec jajowodu otwierający się do jamy otrzewnej w sąsiedztwie jajnika jest rozszerzony w kształcie trąbki - zwanej lejkiem jajowodu; z brzegu lejka zwisają na kształt frędzli wypustki zwane strzępkami jajowodu (fimbriae tubae). Powierzchnię wewnętrzną strzępków pokrywa błona śluzowa, tworząca na większych strzępkach podłużne fałdy, będące przedłużeniem fałdów śluzówki lejka. Jeden ze strzępków, zwany strzępkiem jajnikowym, wyróżnia się tym, że jest dłuższy, głębiej wyżłobiony i przylega do bieguna górnego jajnika. Lejek jest najbardziej ruchomą częścią jajowodu, zazwyczaj skierowaną ku dołowi. Strzępki prawdopodobnie ułatwiają wprowadzenie komórki jajowej '304
z pękniętego pęcherzyka do ujścia brzusznego jajowodu, którego średnica jest tak mała, że przepuszcza tylko grubszą szczecinkę. Lejek przchodzi w bańkę (ampulla), która stanowi ponad połowę całej długości jajowodu. Bańka ma przebieg kręty. Średnica jej wynosi 8 - 9 mm. Ściany są cienkie. Za bańką jajowód zwęża się tworząc cieśń (isthmus tubae) długości 3 - 6 cm, średnicy 2-3 mm o przebiegu prostolinijnym i grubych ścianach, dzięki czemu ma on konsystencję twardego powrózka. Ostatni odcinek jajowodu przebijający ścianę macicy (część maciczna - pars uteńna), długości ok. 1,5 cm, jest najwęższy (średnica wynosi 0,5-1 mm). Odcinek ten kończy się ujściem macicznym (ostium uterinum). Ściana jajowodu składa się z trzech warstw: błony surowiczej, błony mięśniowej oraz błony śluzowej. Komórka jajowa po wydostaniu się z pękniętego pęcherzyka na powierzchnię jajnika zostaje przeprowadzona, prawdopodobnie przy udziale strzępków, głównie strzępka jajnikowego do jajowodu. Dzięki luźnemu zawieszeniu jajnika, pozwalającemu na zmiany jego położenia w szerokim zakresie, oraz ruchliwości końca jajnikowego jajowodu strzępki jajowodu mogą się znaleźć nawetj przy jajniku strony przciwnej, co niejednokrotnie udało się zaobserwować podczas operacji. Możliwe jest zatem przeniesienie komórki jajowej np. z jajnika lewego do jajowodu po stronie prawej. Przesuwanie się jaja w jajowodzie odbywa się pod wpływem rytmicznych skurczów jego mięśniówki. Zapłodnienie komórki jajowej następuje najprawdopodobniej podczas jej wędrówki w bańce jajowodu. Macica Macica (uterus, metra, hystera) jest swoistą wylęgarnią, w której zapłodnione jajo, zarodek, a następnie płód, rozwija się i w ciągu około 280 dni osiąga zdolność do samoistnego życia poza ustrojem matki. W okresie t y m macica dostarcza rozwijającemu sie nłodowi substancji odżywczych oraz materiału na Więzadło Jdjowod wieszadłowe I jajnika Krezka jajnika ,
Jajnik (przekrój)
>-
p
Więzadło obłe macicy
Marszczki pochwowe Ryc. 149. Pochwa, macica, jajowód i jajnik.
'305
budowę jego tkanek. Z chwilę zakończenia rozwoju płodowego macica jako narząd mięśniowy odgrywa główną rolę w wydaleniu płodu na zewnątrz w czasie porodu. Macica ma kształt nieco spłaszczonej gruszki, długości 7 - 9 cm, cieńszym końcem zwróconej ku dołowi. Rozróżnia się w niej dwie zasadnicze części grubszą i szerszą, zwaną trzonem (corpus uteri), oraz węższą - szyjkę (cervix uteri). Zaokrąglona górna część macicy, będąca faktycznie jej sklepieniem, nazywa się dnem (fundus uteri). Po obu stronach dna uchodzą do macicy jajowody. Powierzchnia przednia macicy zwrócona jest ku pęcherzowi moczowemu, powierzchnia tylna - ku odbytnicy. Powierzchnie te łączą się ze sobą wzdłuż lekko zaokrąglonych brzegów bocznych, prawego i lewego, wzdłuż których odchodzi od macicy jej więzadło szerokie (ligamentum latum uteri). Część pochwowa szyjki macicy ma kształt półkulistego czopu, po środku którego widnieje ujście macicy (ostium uteri). U kobiet, które nie rodziły, ma ono kształt okrągłego otworka, a u wieloródek-wąskiej szpary, ograniczonej dwiema wargami - przednią i tylną. Wewnątrz trzonu znajduje się szczelinowata przestrzeń - jama macicy (cavum uteri). Ma ona kształt płaskiego trójkąta podstawą zwróconego ku górze. W kątach bocznych jamy macicy znajdują się ujścia maciczne jajowodów, w kącie dolnym - otwór prowadzący do kanału szyjki macicy (canaiis cervicis uteri), zwanej dawniej ujściem wewnętrznym macicy. Kanał ten otwiera się do pochwy wspomnianym wyżej ujściem macicy. Długość całej jamy (kanału i jamy macicy), mierzona zgłębnikiem, wynosi ok. 6-7 cm. Na granicy między jamą i kanałem szyjki macicy leży zwężony (szerokości ok. 1 cm) odcinek cieśni macicy. Ścianę macicy tworzy gruba (ok. 2 cm) warstwa mięśniowa (myometrium), pokryta od zewnątrz otrzewną zwaną omaciczem (perimetrium), od wewnątrz wysłana błoną śluzową (endometrium). Szyjka macicy zbudowana jest głównie ze zbitej tkanki włóknistej, zawierającej pewną ilość włókien mięśniowych gładkich o układzie przeważnie okrężnym. . Komórki mięśni gładkich macicy mają długość około 50 [xm. W macicy ciężarnej komórki te ulegają przerostowi i mogą wydłużać się do 500 jim i więcej, a masa całego narządu powiększa się około 2Ó-krotnie (od 50 do 1000 g). Poza okresem ciąży i porodu mięśniówka macicy nie ma prawie żadnych zadań i tylko przy obfitszej menstruacji wykonuje odczuwane boleśnie silniejsze skurcze, opróżniające jamę macicy. Bodźcem troficznym zapobiegającym zanikowi macicy są hormony jajnikowe wywołujące w niej cykliczne zmiany. Pod w p ł y w e m tych hormonów w okresie przedmiesiączkowym następuje zmniejszenie się ilości tkanki łącznej i powiększenie się włókien mięśniowych. Zwiększa się przy tym ich rozciągliwość i zmienia reaktywność na środki farmakologiczne. W ciąży macica powiększa się, by dostarczyć miejsca rosnącemu płodowi. Następuje przy t y m nie tylko znaczny przerost komórek mięśniowych, lecz '306
także rozkręcanie się i rozsuwanie spiralnie przebiegających pęczków mięśniowych w obrębie trzonu macicy. Szyjka macicy nie ulega przy tym większym zmianom i zamyka jamę macicy od dołu, natomiast trzon powiększa się ku górze. Następujące przy tym rozluźnienie struktury ściany w obrębie cieśni pomiędzy trzonem i szyjką macicy daje się wymacać przy badaniu oburęcznym (objaw Hegara). Błona śluzowa macicy (tunica mucosa s. endometrium) przylega bezpośrednio do błony mięśniowej, gdyż nie ma w macicy podśluzówki. Powierzchnię błony śluzowej pokrywa nabłonek jednowarstwowy walcowaty, poprzetykany grupami komórek urzęsionych. W obrębie trzonu macicy nabłonek tworzy gruczoły cewkowe proste, które zmieniają swój wygląd zależnie od fazy cyklu miesiączkowego i prawie wcale nie wytwarzają żadnej wydzieliny. W kanale szyjki gruczoły są silniej rozgałęzione (glandulae cervicales uteri). Produkują one galaretowato-śluzową wydzielinę, która tworzy śluzowy czop wypełniający kanał szyjki i chroniący jamę macicy przed wtargnięciem zarazków chorobotwórczych. Podstawowa warstwa błony śluzowej jest ukrwiona przez tętniczki tworzące liczne rozgałęzienia oraz gęstą sieć naczyń włosowatych zalegających w błonie śluzowej i przyległej warstwie mięśniowej. Powierzchowna warstwa błony śluzowej jest ukrwiona przez sieć naczyń włosowatych pochodzących z tzw. tętnic spiralnych. U dojrzałej płciowo, nie ciężarnej kobiety błona śluzowa macicy podlega okresowym zmianom, które są sterowane czynnością jajnika. Zmiany te zwane cyklem miesiączkowym powtarzają się w odstępach od 21 do 35 dni, przeważnie co 28 dni, począwszy od okresu pokwitania (ok. 14 r. ż.) aż do menopauzy (ok. 45-50 r. ż.). W przebiegu cyklu miesiączkowego rozróżnia się w błonie śluzowej macicy cztery okresy lub fazy: 1. Faza wzrastania. 2. Faza wydzielnicza. 3. Faza niedokrwienia. 4. Faza złuszczania, czyli krwawienie miesiączkowe. 1. Faza wzrastania (stadium proliferationis) sterowana hormonem pęcherzykowym jajnika rozpoczyna się z końcem krwawienia miesiączkowego i trwa do utworzenia ciałka żółtego, tj. około 11 dni, od piątego do szesnastego dnia cyklu (licząc od dnia wystąpienia krwawienia miesiączkowego). Charakteryzuje się licznymi podziałami mitotycznymi wszystkich komórek błony śluzowej, która pogrubia się prawie czterokrotnie (do 2-3 cm). 2. Faza wydzielnicza (stadium secretionis) rozpoczyna się w drugiej połowię cyklu miesiączkowego pod wpływem hormonu ciałka żółtego - progesteronu, i trwa w przybliżeniu od szesnastego do dwudziestego siódmego dnia cyklu. Charakteryzuje się obrzmieniem błony śluzowej i wzmożoną sekrecją jej gruczołów, które zaczynają wydzielać śluz. Komórki zawierają dużo glikogenu, który przechodzi także do wydzieliny śluzowej. Zwiększone ukrwieme wzmaga '307
aktywność wydzielniczą błony śluzowej. Grubość błony śluzowej zwiększa się podczas fazy wydzielniczej prawie dwukrotnie. Ostatecznym celem wszystkich tych zmian jest przygotowanie błony śluzowej do przyjęcia zapłodnionego jaja i stworzenie warunków dla jego implantacji. Wydzielina błony śluzowej jajowodu i macicy dostarcza zapłodnionej komórce jajowej materiału odżywczego w okresie pierwszych jego podziałów. Z chwilą gdy nastąpi implantacja zapłodnionego jaja, komórki trofoblastu zaczynają trawić błonę śluzową macicy, dostarczając substancji odżywczych dla szybko rozwijającego się zarodka. 3. Faza niedokrwienia (stadium ischemicum). Jeżeli w drugiej połowie cyklu nie nastąpi zapłodnienie i zagnieżdżenie się jaja, to w 27 lub 28 dniu cyklu (w 13-14 dniu po owulacji) tętnice spiralne błony śluzowej ulegają obkurczeniu, wskutek czego błona śluzowa staje się na wiele godzin blada. Skurcz jest prawdopodobnie następstwem ustania produkcji estrogenów, które mają działać rozkurczająco na naczynia błony śluzowej. W głębszych jej warstwach przepływ krwi nie zostaje zakłócony. 4. Faza złuszczania (stadium desquamationis), czyli krwawienie miesiączkowe (menstruatio), następuje wskutek ponownego otwarcia się obkurczonych tętnic. Pod naporem napływającej krwi naczynia krwionośne położone blisko powierzchni błony śluzowej pękają, krew wylewa się do zrębu błony śluzowej i przedostaje się do światła macicy. Martwiczo zmienione wskutek uprzedniego niedokrwienia powierzchowne warstwy śluzówki oddzielają się od ściany macicy i po upływie około 48 h cała ta część śluzówki ulega złuszczeniu. Oddzielone tkanki pomieszane z krwią i wydzieliną gruczołów zostają wydalone na zewnątrz. Podczas menstruacji następuje utrata około 35 ml krwi i ok. 35 ml płynu surowiczego. Dzięki obecności fibrynolizyny wyzwolonej z martwiczych tkanek krew miesiączkowa w warunkach prawidłowych nie krzepnie. Jeżeli krwawienie z otwartych naczyń jest nadmierne, ilość fibrynolizyny może być niedostateczna, a wówczas pojawiają się skrzepy. Obecność skrzepów we krwi miesiączkowej zwykle wskazuje na proces chorobowy. Krwawienie miesiączkowe zwykle trwa nie dłużej niż pięć dni, po czym rozpoczyna się proces odnowy czynnościowej warstwy błony śluzowej (endometrium). Zewnętrzną warstwę ściany macicy tworzy pokrywająca ją otrzewna zwana omaciczem (perimetrium). Ponieważ macica zajmuje w miednicy położenie centralne, otrzewna od przodu przechodzi z pęcherza moczowego na szyjkę macicy, tworząc tu szczelinowate zagłębienie pęcherzowo-maciczne (excavatio vesico-uterina), zaś od tyłu schodzi na macicę z odbytnicy znacznie niżej. Pokrywając częściowo także tylne sklepienie pochwy, otrzewna tworzy tu zagłębienie odbytniczo-maciczne (excavatio rectouterina). W prawidłowym ustawieniu macicy, w przodopochyleniu i przodozgięciu, w zagłębieniu t y m znajdują się pętle jelita cienkiego. Wzdłuż obu brzegów bocznych otrzewną schodząc z macicy tworzy podwójny fałd, zwany więzadłem szerokim macicy (ligamentum latum uteri), który dochodzi do ściany bocznej miednicy mniejszej. Wygląda to tak, jak gdyby '308
macica wraz z jajowodem została wpuklona ku górze od dna miednicy, tworząc przed sobą worek otrzewnowy, co wszakże nie odpowiada faktycznemu rozwojowi tych tworów. Razem z macicą więzadło szerokie przegradza poprzecznie jamę miednicy mniejszej na dwie części - przednią i tylną. Macica tkwi zatem w więzadle szerokim jakwfałdziefartucha,ztym że część owego fartucha, pokrywająca macicę, nazywa się omaciczem (perimetrium). Pomiędzy blaszkami więzadła szerokiego znajduje się niewielka ilość tkanki łącznej. Więcej tkanki łącznej luźnej znajduje się przy brzegach bocznych macicy, gdzie przebiegają zdążające ku niej naczynia i nerwy. U podstawy więzadła szerokiego jego blaszki rozchodzą się, a zawarta pomiędzy nimi tkanka łączna wraz z leżącymi w niej tworami nazywa się przymaciczem (parametrium). Silniej rozwinięte pasma łącznotkankowe przymacicza, zawierające wiązki mięśniówki gładkiej, noszą nazwę więzadeł (ligamenta). Tworzą one aparat więzadłowy macicy. Największe znaczenie mają pasma biegnące od szyjki macicy ku ścianom miednicy mniejszej (ligamenta cardinalia uteri). Ku przodowi biegnie z szyjki macicy więzadło pęcherzowo-maciczne (ligamentum vesicouterinum), a ku tyłowi - więzadło odbytniczo-maciczne (ligamentum rectouterinum). Od bocznego brzegu trzonu macicy odchodzi więzadło obłe macicy (ligamentum teres uteri), które w postaci łącznotkankowego powrózka długości 12-14 cm biegnie łukowato ku przedniej ścianie jamy brzusznej, którą przebija skośnie w miejscu odpowiadającym kanałowi pachwinowemu i kończy się w tkance podskórnej wargi sromowej większej i wzgórka łonowego. Wymienione wyżej więzadła przyczyniają się w sposób istotny do utrzymania macicy w jej prawidłowym położeniu, jednakże główną rolę ogrywają tu mięśnie i powięzie dna miednicy. Macica, jak wspomniano, leży po środku miednicy mniejszej, mniej więcej w jej osi. Takiemu położeniu zawdzięcza ona ogromną ruchomość - macica daje się przesuwać we wszystkich kierunkach. W prawidłowym położeniu macica jest pochylona do przodu (przodopochylenie- anteversió). Powierzchnia przednia trzonu przylega do pęcherza moczowego, tylna - styka się z pętlami jelitowymi. Pomiędzy trzonem a szyjką macica jest ponadto zgięta ku przodowi pod kątem rozwartym (przodozgięcie - anteflexió). W pozycji leżącej kobiety część pochwowa szyjki macicy styka się z tylną ścianą pochwy. Oś macicy nie stanowi przedłużenia osi pochwy, lecz tworzy z nią kąt rozwarty ku przodowi. Układ taki przyczynia się do utrzymania macicy w jej centralnym położeniu w miednicy mniejszej. Gdyby cały kanał rodny przebiegał w jednej prostej osi, będącej przedłużeniem otworu miednicy mniejszej, wówczas pod naporem tłoczni brzusznej mogłoby łatwo dojść do wypadnięcia macicy. W ginekologii oprócz położenia prawidłowego rozróżnia się jeszcze różne rodzaje przesunięcia macicy w stosunku do osi miednicy - ku przodowi (antepositio), ku tyłowi (retropositio) lub bocznie (laterpositio). Poza tym macica może być uniesiona ponad płaszczyznę wejścia do miednicy (elevatio uteri) lub opuszczona głęboko do miednicy (depressio s.descensus uteri). '309
Najczęstszą anomalią położenia macicy jest jej tyłozgięcie (retroflexio), przeważnie skojarzone z tyłopochyleniem (retroversio). Przy takim położeniu zwykle bywają obfite krwawienia miesiączkowe oraz utrudnione jest zapłodnienie. Pochwa Pochwa (vagina, colpos) jest przewodem mięśniowo-błoniastym, który łączy przedsionek pochwy z macicą. Głównym zadaniem pochwy jako narządu płciowego jest przyjęcie nasienia wprowadzonego do niej podczas stosunku płciowego przez członek męski. W czasie porodu pochwa służy jako kanał wyprowadzający dla płodu, poza tym stanowi drogę odpływu krwi menstruacyjnej i innych produktów wydzielania macicy. Pochwa prawie w całości leży w miednicy mniejszej pomiędzy pęcherzem i cewką moczową od przodu, a odbytnicą od tyłu. Krótki odcinek końcowy przebija przeponę moczowo-płciową. Oś pochwy ma kierunek prawie prostolinijny i w pozycji pionowej skierowana jest ku górze i ku tyłowi, tworząc z płaszczyzną poziomą kąt 50-70 . W pozycji leżącej na grzbiecie pochwa ma kierunek zbliżony do poziomego. Z osią macicy pochwa tworzy kąt nieco większy od prostego rozwarty ku przodowi. Ściany pochwy przylegają do siebie i na przekroju poprzecznym pochwa w górnym odcinku ma kształt poprzecznej szczeliny, w dolnym - kształt litery H. Tylko górny koniec pochwy obejmujący szyjkę macicy ma przekrój cylindryczny. Długość pochwy bywa zmienna i może się wahać między skrajnymi wartościami od 5 do 14 cm. Przeciętnie długość pochwy mierzona wzdłuż ściany przedniej wynosi ok. 7,5 cm, wzdłuż ściany tylnej - 9 cm, a to wskutek tego, iż ściana tylna przyczepia się nieco wyżej do szyjki macicy niż ściana przednia. Dzięki takiemu umocowaniu górna część pochwy tworzy zachyłki, zwane sklepieniami pochwy: najgłębsze - sklepienie tylne (fornix vaginae posterior), następnie dwa sklepienia boczne i sklepienie przednie. Średnica pochwy nie jest jednakowa na całej długości. W części środkowej wynosi ona przeciętnie 2-3 cm. Najwęższa jest pochwa w części przeponowej, najszersza - w części obejmującej szyjkę macicy. Przednia ściana pochwy jest zrośnięta ściśle z podstawą pęcherza i cewką moczową. Przemieszczenie jednego narządu pociąga za sobą przesunięcie drugiego, np. obniżenie przedniej ściany pochwy, pociągając dno pęcherza, może spowodować powstanie torbieli pęcherza i przetokę. Ściana tylna pochwy jest związana z odbytnicą znacznie luźniej. Przestrzeń pomiędzy pochwą a odbytnicą wypełnia masa tkanki mięśniowej i włóknistej krocza. Po bokach pochwy znajdują się dźwigacze odbytu i mięśnie przepony miedniczej. Ściana pochwy, grubości ok. 3 mm, składa się z trzech warstw: 1) błony zewnętrznej, 2) błony mięśniowej i 3) błony śluzowej. 1. Błonę zewnętrzną stanowi cienka warstwa zbitej tkanki łącznej oraz tkanka łączna luźna, łącząca pochwę z narządami sąsiednimi. Znajduje się tu także obfity splot żylny oraz włókna i skupienia komórek nerwowych. '310
2. Środkowa warstwa mięśniowa składa się z włókien mięśniowych gładkich ułożonych w dwie warstwy - zewnętrzną podłużną i wewnętrzną okrężną. Warstwa podhiżna stanowi u góry przedłużenie mięśniówki macicy, u dołu wplata się w utkanie włókien poprzecznie prążkowanych mięśni dna miednicy. W części dolnej pochwę otacza pierścień włókien mięśniowych prążkowanych, określany jako m. opuszkowo-gąbczasty (m. bulbospongiosus). 3. Błona śluzowa jest ściśle zrośnięta z warstwą mięśniową. Składa się z warstwy właściwej pokrytej wielowarstwowym nabłonkiem płaskim, podobnym do naskórka, lecz nie rogowaciejącym. Błona śluzowa pochwy nie zawiera gruczołów i jest zwilżana przez śluz pochodzący z gruczołów szyjki macicy oraz płyn surowiczy przesiąkający ze splotu naczyniowego otaczającego jej ścianę. Po osiągnięciu dojrzałości płciowej nabłonek pochwy grubieje, zwiększa się w nim także zawartość glikogenu. Zawartość glikogenu w komórkach ulega cyklicznym wahaniom - zwiększa się w fazie owulacji i zmniejsza pod koniec cyklu. Fermentacyjne działanie pewnych bakterii (Bacillus Dóderleini) powoduje wytwarzanie z glikogenu kwasu mlekowego; kwaśny odczyn pochwy chroni macicę przed inwazją drobnoustrojów chorobotwórczych. U kobiet młodych błona śluzowa pochwy tworzy na ścianie przedniej i tylnej podłużne wzniesienie, od którego odchodzą liczne poprzeczne f a ł d y - m a r s z czki pochwowe (rugae vaginales), silniej rozwinięte na tylnej ścianie i w dolnej części pochwy. Po kilku porodach oraz w wieku starszym fałdy te zanikają i powierzchnia śluzówki wygładza się. Wspomniane wzniesienia wraz z fałdami poprzecznymi noszą nazwę słupów marszczek (columnae rugarum) przedniego i tylnego. U dziewicy dolny koniec pochwy jest częściowo zamknięty poprzecznym fałdem śluzówki o cienkim zrębie łącznotkankówym, zwanym błoną dziewiczą (hymen). W nabłonku śluzówki pochwy zachodzą cykliczne zmiany zależne od wpływu hormonów jajnikowych, głównie estrogenów. Wykorzystuje się je w praktyce dla określenia stadiów cyklu jajnikowego oraz stopnia aktywności estrogenowej. Badanie cytologiczne rozmazów pochwowych jest najprostszą, najszybszą i najtańszą metodą określania tych czynników. Metodą tą w o b u przypadkach można określić sześć stadiów cyklu jajnikowego: 1. Wczesną fazę pęcherzykową (6-7 dni), charakteryzującą się w y s t ę p o w a n i e m mniej więcej jednakowej liczby komórek nabłonkowych z jądrem średniej wielkości i leukocytów. 2. Zaawansowaną fazę pęcherzykową (8-11 dzień), w której liczba komórek kwasochłonnych zwiększa się, leukocyty znikają. 3. Późną fazę pęcherzykową (12-14 dzień) z przewagą komórek kwasochłonnych. Jądra komórek są małe. Obfity śluz. Komórki pojedyncze oddzielone od siebie. 4. Wczesną fazę luteinową (15-17 dzień). Komórki są pomarszczone lub o zawiniętych brzegach; komórki w skupieniach. Wzrost liczby komórek zasadochłonnych. Pojawiają się leukocyty. Jądra skurczone, małe. 5. Zaawansowaną (18-24 dzień) i późną fazę luteinową (25-28 dzień). Komórki kwasochłonne pojedyncze, dalszy wzrost J
'311
V 6. Menstruację (1-5 dzień). Silne złuszczenie nabłonka, gęste skupienia komórek. Krwinki czerwone, leukocyty, histiocyty, strzępki komórek błony śluzowej macicy.
Wskaźnikiem aktywności estrogenowej jest także zawartość glikogenu w nabłonku pochwowym. Dla orientacyjnej oceny tej aktywności wysuszone lub utrwalone rozmazy ogląda się pod mikroskopem po zadziałaniu roztworem lub parami jodu.
Zewnętrzne części płciowe żeńskie Do zewnętrznych części płciowych żeńskich, obejmowanych także nazwą sromu niewieściego (pudendum femininum), należą: 1) wzgórek łonowy, 2) wargi sromowe większe, 3) wargi sromowe mniejsze, 4) łechtaczka, 5) przedsionek pophwy, 6) opuszka przedsionka oraz 7) gruczoły przedsionkowe większe. Wzgórek łonowy Spoidło przednie warg Napletek łechtaczki Żołądź łechtaczki Wędzidełko łechtaczki
Warga sromowa większa Warga sromowa mniejsza
Ujście zewnętrzne cewki
Przedsionek, pochwy
Marszczki pochwowe Krocze
Odbyt Ryc. 150. Srom.niewieści.
Wzgórek ł o n o w y Wzgórek łonowy (mons pubis) jest to zaokrąglona wyniosłość przed spojeniem łonowym, utworzona przez większe skupienie tkanki tłuszczowej. Ma on kształt trójkąta, którego wierzchołek skierowany jest ku dołowi, gdzie łączy się z wargami sromowymi większymi. Wysokość (grubość) wzgórka łonowego wynosi przeciętnie ok. 2 - 3 cm, u kobiet otyłych może sięgać nawet do 8 cm. W okresie pokwitania wzgórek łonowy pokrywa się szorstkimi włosami ł o n o w y m i (pubes). Granica obszaru pokrytego włosami ma przebieg poziomy. 312
Wargi sromowe większe Wargi sromowe większe (labia majora pudendi) są to dwa podłużne silnie rozwinięte fałdy skórne, będące odpowiednikiem moszny. Każda z nich ciągnie się od wzgórka łonowego ku dołowi i tyłowi ograniczając szparę sromową. W tyle wargi sromowe zbliżając się do odbytu stają się coraz bardziej płaskie i gubią się w skórze krocza w niewielkiej odległości od siebie. Każda warga ma dwie powierzchnie - zewnętrzną, pokrytą ciemno zabarwioną, owłosioną skórą, obficie zaopatrzoną w gruczoły łojowe i potowe, oraz powierzchnię wewnętrzną, znacznie delikatniejszą, w głębi wilgotną, przypominającą błonę śluzową. Najgrubsze są wargi sromowe od przodu - ich przyśrodkowe brzegi łączą się tutaj ze sobą tworząc spoidło przednie przebiegające nad łechtaczką. Od tyłu wargi są połączone szerszą listewką skórną, zwaną spoidłem tylnym, położonym ok. 2,5 cm od odbytu. U młodej, nie podnieconej płciowo, kobiety wargi sromowe większe ściśle przylegają do siebie w linii środkowej, chroniąc położone głębiej s t r u k t u r y wargi sromowe mniejsze, ujście pochwy i cewki moczowej. Urazy porodowe mogą osłabić konstrukcję uciskową zrębu warg, wskutek czego tracą one napięcie, vyiotczeją i mogą luźno zwisać. Wargi sromowe mniejsze Wargi sromowe mniejsze (labia minora pudendi) ukazują się w szparze sromowej po rozchyleniu warg większych, jako dwa cienkie różowoczerwone fałdy skórne ciągnące się od łechtaczki skośnie w dół i ku tyłowi. Od przodu każda z tych warg dzieli się na dwa ramiona. Ramiona górne obu stron łączą się ze sobą nad łechtaczką, tworząc fałd skórny pokrywający ją na kształt kapturka, zwany napletkiem łechtaczki. Ramiona dolne łączą się na dolnej powierzchni łechtaczki tworząc wędzidełko (frenulum clitoridiś).\N ten sposób trzon łechtaczki jest zwykle całkowicie schowany pomiędzy wspomnianymi ramionami warg mniejszych i występuje tylko jej żołądź. Silniej rozwinięty napletek może pokrywać także całą żołądź, która uwidacznia się dopiero po odciągnięciu napletka lub podczas wzwodu łechtaczki. Przyśrodkowe powierzchnie warg sromowych mniejszych ograniczają boczne ujście pochwy (ostium vaginae). W tyle wargi sromowe mniejsze łączą się za ujściem pochwy tworząc delikatny fałd, zwany wędzidełkiem warg sromowych (frenulum labiorum pudendi), który przeważnie rozrywa się przy pierwszym porodzie. Wędzidełko to ogranicza od tyłu płytkie zagłębienie, zwane dołem przedsionka pochwy (fossa vestibuli vaginae). Wargi sromowe mniejsze pokryte są nabłonkiem wielowarstwowym płaskim spoczywającym na zrębie łącznotkankowym zaopatrzonym obficie w naczynia krwionośne, ale pozbawionym tkanki tłuszczowej. Na obu powierzchniach warg mniejszych znajdują się liczne gruczoły łojowe.
'313
V Przedsionek pochwy Przedsionek pochwy (vestibulum vaginae) ukazuje się po rozchyleniu warg sromowych większych i mniejszych jako eliptyczna płytka przestrzeń, ograniczona z boków wargami sromowymi mniejszymi; od przodu - wędzidełkami łechtaczki, od tyłu - wędzidełkiem warg sromowych mniejszych. W szparze tej znajduje się ujście zewnętrzne cewki moczowej i ujście pochwy. Ujście zewnętrzne cewki moczowej (ostium urethrae externum) leży w przedsionku pochwy, mniej więcej 2,5 cm poniżej łechtaczki. Ma ono postać okrągłego lub szczelinowatego otworu na niewielkiej brodawce cewkowej. Poniżej brodawki cewki moczowej znajduje się ujście pochwy (ostium vaginae). U dziewicy ujście to jest częściowo zamknięte cienkim fałdem błony śluzowej, zwanym błoną dziewiczą. t
Błona dziewicza Błona dziewicza (hymen) ma najczęściej kształt pierścienia (szerszego od tyłu) lub półksiężycowatego fałdu wklęsłością zwróconego ku przodowi (hymen semilunaris s. falciformis). Niekiedy błona dziewicza ma kształt regularnego pierścienia z otworem pośrodku (hymen annularis). Przy silniej rozwiniętych częściach bocznych błony dziewiczej ujście pochwy ma kształt owalny. Czasami wolny brzeg tej błony bywa od samego początku karbowany (hymen denticulatus), postrzępiony (hymen fimbriatus) lub płatowaty (hymen lobatus). Przy pierwszym stosunku płciowym kobiety błona dziewicza, jeśli nie było w niej większego otworu, zostaje zwykle naddarta (defloratio), z czym związane bywa niewielkie krwawienie. Błona pęka najczęściej w jednym lub kilku miejscach w części tylnej, wskutek czego powstaje z niej kilka płatków (lobi hymenales), Łechtaczka Łechtaczka (clitoris) jest nieparzystym narządem, pełniącym rolę receptora i transformatora bodźców czuciowych inicjujących i wzmagających pobudzenie płciowe. Jest ona odpowiednikiem członka męskiego i podobnie jak on składa się z dwóch ciał jamistych łechtaczki, otoczonych mocną łącznotkankową błoną białawą. Utworzony przez ciała jamiste trzon łechtaczki (corpus clitoridiś) ma kształt cylindra długości ok. 2 cm, zakończonego zaokrąglonym wierzchołkiem dokoła powleczonym skórą, zwanym żółędzią łechtaczki. Średnica łechtaczki wynosi przeciętnie 4-5 mm, ale łechtaczka o średnicy 10 mm nie należy do nieprawidłowości anatomicznych. Żołądź łechtaczki jest bardzo wrażliwa na dotyk. Pod naskórkiem jej znajdują się bardzo liczne nerwy oraz ciałka czuciowe (ciałka Pacinego). W odróżnieniu od żołędzi prącia, łechtaczki nie przebija cewka moczowa. 314
Łechtaczka reaguje na pobudzenie płciowe wzwodem, którego mechanizm jest podobny jak w prąciu. Następuje przy tym przeważnie wyraźne powiększenie żołędzi. Najszybciej reaguje łechtaczka na bezpośrednie drażnienie. Opuszki przedsionka Opuszki przedsionka (bulbi vestibuli) są to parzyste twory zbudowane z tkanki łącznej gąbczastej, zawarte w ścianie sromu u podstawy warg sromowych mniejszych po obu stronach ujścia pochwy. Odpowiadają one opuszce prącia. Jeżeli sobie wyobrazimy cewkę moczową męską od dołu rozszczepioną i szeroko rozsuniętą na boki, przez co zostanie otwarta zatoka moczowo-płciowa, to ciała gąbczaste cewki znajdą się w miejscu opuszek przedsionka kobiety. Mają one kształt migdała długości 3 - 4 cm. Od przodu łączą się ze sobą wąskim pasmem, zwanym spoidłem opuszek. Powierzchnia górna przylega do przepony moczowo-płciowej, dolną pokrywa mięsień opuszkowo-gąbczasty. Koniec tylny opuszki styka się z gruczołem przedsionkowym większym. Wypełnienie zatok żylnych opuszki tworzy z niej rodzaj poduszki wyściełającej wejście do pochwy. Opuszki, podobnie jak ciało jamiste prącia, nie ulegają silniejszemu wzwodowi. Gruczoły przedsionkowe większe Gruczoły przedsionkowe większe (Bartoliniego) (glartdulae vestibulares majores) są odpowiednikiem gruczołów cewkowych męskich. Są to dwa małe okrągławe lub owalne twory wielkości małego ziarnka fasoli, barwy czerwonawożółtawej, położone po obu stronach ujścia pochwy, bocznie od części tylnej przedsionka pochwy. Każdy z tych gruczołów styka się z opuszką przedsionka odpowiedniej strony, a częściowo jest przez nią pokryty. Przewód wyprowadzający każdego gruczołu, długości ok. 2 cm, ptwiera się bocznie od błony dziewiczej w rowku pomiędzy brzegiem przytwierdzonym błony a wargami sromowymi mniejszymi. Gruczoły przedsionkowe produkują wydzielinę śluzową. Sutek Sutek (mamma) jest parzystym narządem o kształcie półkulistym lub stożkowatym, który rozwija się u kobiety w okresie pokwitania. Sutki są położone symetrycznie na przedniej ścianie klatki piersiowej, gdzie zajmują okolicę pomiędzy trzecim i szóstym żebrem oraz między mostkiem i dołem pachowym. Szerokość zawartej między sutkami przestrzeni (sinus mammarum) jest indywidualnie zmienna. Nieco poniżej środka sutka, zazwyczaj na poziomie czwartej przestrzeni międzyżebrowej lub piątego żebra, wznosi się na powierzchni narządu cylindryczna lub stożkowata, ciemniej zabarwiona brodawka sutkowa {papilla mammae). U kobiet, które rodziły, pigmentacja brodawki jest silniejsza. '315
Fahl p;idiowy przedni Ramię—i„Brodawka sutkowa Otoczka -brodawki sutkowej Ryc. 151. Sutek 18-letniej dziewczyny.
Płaciki gruczołu sutkowego
\ Otoczka, brodawki sutkowej
Żebra
M. piersiowy większy
Brodawka, sutkowa Przewody mleczne Płaty gruczołu sutkowego Ryc. 152. Gruczoł sutkowy w przekroju (schemat).
Brodawka leży na środku okrągłego silniej zabarwionego pola, zwanego otoczką brodawki (areola mammae). Na wierzchołku brodawki znajduje się 12-15 drobnych otworków stanowiących ujścia przewodów mlecznych. Kształt i wielkość brodawek bywają zmienne. Niekiedy brodawki są tak słabo rozwinięte, że utrudniają lub uniemożliwiają noworodkowi ssanie. Brodawka jest zaopatrzona we włókna mięśniowe gładkie o układzie okrężnym i spiralnym. U podstawy brodawki włókna te przechodzą w utkanie otoczki. Skurcz mięśniówki powoduje erekcję brodawki. Wielkość sutka jest indywidualnie zmienna i zależy przede wszystkim od ilości zawartej w nim tkanki tłuszczowej. W tkance tej zalegają gruczoły sutkowe skupione w 15-20 płatów ułożonych promienisto dokoła brodawki. W czasie ciąży gruczoły sutkowe rozrastają się i pod koniec ciąży zaczynają '316
produkować mleko. Zmiany w gruczołach sutkowych zachodzące podczas cyklu miesięcznego dotyczą głównie tkanki łącznej międzyzrazikowej. Obrzmienie sutka przed menstruacją następuje wskutek przekrwienia tkanki międzyzrazikowej. AKT PŁCIOWY, SPÓŁKOWANIE (COITUS) Warunkiem dokonania aktu płciowego przez mężczyznę jest skuteczny wzwód prącia i wytrysk nasienia. Wzwód prącia (którego mechanizm opisano w innym miejscu) u młodego seksualnie sprawnego mężczyzny następuje odruchowo w ciągu 3 - 8 s od zadziałania odpowiednich bodźców inicjujących. Najważniejszym źródłem tych bodźców jest żołądź prącia. Seksualne bodźce czuciowe mogą pochodzić także z innych okolic przyległych do prącia. Jedną z przyczyn wzmożonego popędu płciowego u mężczyzny jest prawdopodobnie przepełnienie wydzielinami narządów płciowych. Działanie leków wzmagających popęd płciowy (aphrodisiaca) typu kantarydyny polega na podrażnieniu śluzówki pęcherza i cewki mocźowej. Wzwód prącia może nastąpić także pod wpływem bodźców psychicznych, takich jak erotyczne myśli lub senne marzenia o spółkowaniu. U wielu mężczyzn, zwłaszcza w okresie pokwitania, czynniki psychiczne mogą doprowadzić do wytrysku nasienia. W późniejszym wieku mechanizm psychicznego pobudzenia staje się mniej czuły i działa wolniej. Kulminacyjnym punktem i zarazem zakończeniem aktu płciowego jest u mężczyzny wytrysk nasienia (e/acu/af/o). Jest to proces odruchowy przebiegający w postaci serii skurczów perystaltycznych przewodów jądra, najądrza i głównie nasieniowodów powodujących wyrzucenie plemników do cewki moczowej. Równocześnie skurcze pęcherzyków nasiennych mięśniówki gruczołu krokowego dołączają wydzieliny tych narządów, które razem ze śluzem z gruczołów opuszkowo-cewkowych tworzą w cewce płynną masę, zwaną nasieniem (sperma). Proces powyższy jest wstępem do właściwego wytrysku, który następuje wskutek rytmicznych skurczów mięśni prążkowanych otaczających opuszkę ciała gąbczastego cewki. Skurcze wytryskowe obejmują cewkę moczową na całej jej długości, powodując wyrzucanie nasienia pod dosyć dużym ciśnieniem. Skurcze te powtarzają się w rytmie 0,8 s. Po pierwszych trzech-czterech skurczach ich częstość i siła wyrzutu zmniejsza się. U kobiety; podobnie jak i mężczyzny, skuteczne dokonanie aktu płciowego zależne jest od odpowiedniego pobudzenia psychicznego i lokalnego w obrębie narządów płciowych. Najwcześniejszym wskaźnikiem pobudzenia płciowego u kobiety jest pojawienie się na ścianach pochwy wydzieliny zwilżającej (lubricatió). Obfite zwilżenie pochwy, które u młodej kobiety jest reakcją prawie natychmiastową na skuteczne pobudzenie płciowe, sprawia że ruchy podczas spółkowania mają charakter masażu, który dostarcza optymalnych bodźców czuciowych dla odruchów kulminujących u obu płci w orgazmie. Orgazm żeński jest odpowiednikiem wytrysku u mężczyzny. U obu płci następuje podczas stosunku płciowego przyspieszenie czynności serca do 100-175 uderzeń na minutę. Wzrasta ciśnienie krwi skurczowe 21 - Anatomia i fizjologia
317
-
•
V
u mężczyzny o 40-100 mm Hg i rozkurczowe o 20-50 mm Hg, u kobiety o 30-80 mm Hg ciśnienie skurczowe i o 20-40 mm Hg ciśnienie rozkurczowe. Przyspieszenie oddechów i hiperwentylacja towarzyszą stosunkowi płciow e m u u obu płci, dochodząc do 40 oddechów na minutę. Skurcze mięśniowe podczas aktu płciowego mają początkowo charakter dowolny. Biorą w nim udział mięśnie tułowia, miednicy i kończyn. W fazie orgazmu u obu płci często występują mimowolne skurcze spastyczne całych grup mięśniowych, np. mięśni twarzy, mięśnia prostego brzucha, mięśnia mostkowo-obojczykowo-sutkowego.
ZAPŁODNIENIE. CIĄŻA. PORÓD. W normalnych warunkach tylko jeden plemnik może zapłodnić komórkę jajową, ponieważ osłonka przejrzysta {zona pellucida) jaja ma postać kratownicy, która z chwilą przebicia błony komórkowej przez pierwszy plemnik wypełnia się wychodzącą z jaja substancją uniemożliwiającą dostęp innym plemnikom. Głowa plemnika wnosi do komórki jajowej 23 chromosomy i zapłodnione jajo, zwane zygotą, uzyskuje pełny garnitur 46 chromosomów. Płeć przyszłego dziecka zależy od rodzaju plemnika, jaki wniknął do komórki jajowej, a ściślej od zawartego w nim chromosomu płciowego. Jeżeli będzie to plemnik niosący chromosom płciowy X, wówczas rozwinie się płeć żeńska; chromosom Y - determinuje płeć męską. Ponieważ komórka jajowa jest wyposażona zawsze w chromosom płciowy X, to w pierwszym przypadku garnitur chromosomów będzie miał postać 42 + X + X, w d r u g i m - 4 2 + X + Y. Podczas owulacji komórka jajowa wydostaje się z jajnika bezpośrednio do jamy otrzewnej, skąd musi wejść do jajowodu. Umożliwiają to strzępki jajnikowe, których wewnętrzna powierzchnia jest pokryta nabłonkiem migawkowym. Jego rzęski wytwarzają ruch kierujący jajo do ujścia brzusznego jajowodu. Zapłodnienie jaja następuje zwykle jeszcze wtedy, zanim trafi ono do jajowodu lub wkrótce po tym, jak się w nim znajdzie. Na przejście jaja przez jajowód do jamy macicy potrzeba około trzech dni. W tym czasie zapłodniona komórka jajowa przechodzi szereg podziałów czerpiąc substancje odżywcze z wydzieliny gruczołów błony śluzowej jajowodu.
Implantacja Po znalezieniu się w macicy zygota przez dalsze 4 - 5 dni czerpie substancje odżywcze z wydzielin błony śluzowej macicy, zwanych „mlekiem maciczn y m " . Wszczepienie (implantacja) dzielącego się jaja do śluzówki macicy następuje w 7 - 8 dni po zapłodnieniu. Kluczową rolę w procesie implantacji '318
Matka
Ojciec
Komórki jajo\A|e
Dziewczynka Ryc. 153. Determinacja płci. Komórka jajowa zawiera zawsze chromosom płciowy X. Plemniki są dwóch rodzajów - z chromosomem płciowym X lub Y. Zapłodnienie plemnikiem X da w wyniku potomka płci żeńskiej, plemnik Y da potomka męskiego.
odgrywają komórki trofoblastu rozwijające się na powierzchni pęcherzyka zarodkowego. Wydzielane przez te komórki enzymy proteolityczne trawią stykającą się z nim błonę śluzową macicy, a równocześnie inne komórki trofoblastu tworzą pomosty przyczepiające trofoblast do brzegów wytrawionego zagłębienia. '319
V W ten sposób zarodek jak gdyby „wyżera" sobie w śluzówce macicy jamkę, do brzegów której jednocześnie się przymocowuje. Komórki trofoblastu szybko-się mnożą, by razem z elementami błony śluzowej macicy utworzyć łożysko i inne błony płodowe.
Ciąża Okres rozwoju płodu w łonie matki od zapłodnienia do porodu nazywamy ciążą (graviditaś). Ciąża trwa u człowieka 10 miesięcy księżycowych, czyli około 280 dni, i kończy się porodem (partus). Rosnący w macicy płód stanowi duże obciążenie dla organizmu matki, który musi mu dostarczyć niezbędnych dla wzrostu i życia substancji. Przez łożysko przepływa ok. 3 / 4 I krwi na minutę. Krążenie w łożysku działa jak przetoka tętniczo-żylna, zmniejszająca opór obwodowy w układzie tętniczym. Gdy w układzie naczyniowym zmniejsza się opór, następuje spadek ciśnienia krwi. Aby utrzymać ciśnienie krwi na odpowiednim poziomie przy tak zmniejszonym oporze, serce musi zwiększyć objętość wyrzucanej krwi o ok. 30-40% powyżej normy. Zwiększa się również u ciężarnej objętość krwi krążącej o ok. 30%. Wskutek zwiększonego zużytkowania substratów metabolicznych zwiększa się zwykle u kobiety w ciąży łaknienie, prowadzące do wzrostu ciężaru ciała. Dodaje się do tego także wzrost ciężaru ciała płodu, ciężar płynu owodniowego, błon płodowych, powiększonej macicy i gruczołów sutkowych. W następstwie wzmożonej aktywności wielu gruczołów dokrewnych z tarczycą włącznie, zwiększa się, zwłaszcza w drugiej połowie ciąży, podstawowa przemiana materii o ok. 15%. v Największy wzrost płodu odbywa się w ostatnim trymestrze ciąży. Ciężar płodu podwaja się wciągu ostatnich dwóch miesięcy. W t y m okresie zapotrzebowanie płodu na materiał budulcowy zwiększa się tak bardzo, że w ostatnim miesiącu ciąży organizm matki nie potrafi wchłonąć i przyswoić z przewodu pokarmowego dostatecznej dla tego celu ilości białka. Wapnia,fosforu, żelaza. Organizm matki pokrywa te potrzeby z zapasów, które gromadzą się w nim od początku ciąży. Jeżeli jednak dieta kobiety ciężarnej nie zawiera odpowiedniej ilości elementów odżywczych, wówczas dochodzi do niedoboru w organizmie matki takich składników, jak wapń, fosfor, żelazo, witaminy. Dieta kobiety w ciąży musi być zatem pełnowartościowa, a niektóre składniki, takie jak żelazo i wapń, powinny być doprowadzone nawet w pewnym nadmiarze.
Poród Proces wydalenia dojrzałego płodu nazywamy porodem (partus). Rozpoczyna się on od rytmicznych skurczów macicy o narastającej sile, które ostatecznie doprowadzają do wydalenia płodu. 320
Przyczyny skurczów porodowch macicy są dwojakie: .1) wpływy hormonalne i 2) czynniki mechaniczne. 1. Progesteron zmniejsza kurczliwość mięśnia macicy, natomiast estrogeny wpływają na jej obkurczanie się. Z postępem ciąży obydwa te hormony są wydzielane we wzrastającej ilości, z t y m że od siódmego miesiąca wydzielanie estrogenów zaczyna przeważać nad progesteronem, przy czym bezpośrednio przed porodem pojawia się we krwi duża ilość nie związanych (wolnych) estrogenów. Hormonem swoiście pobudzającym macicę do skurczów jest oksytocyna z tylnego płata przysadki. 2. Mechanicznym czynnikiem pobudzającym macicę do skurcżów jest rozciąganie jej włókien mięśniowych. Bliźnięta rodzą się zwykle ok. 19 dni wcześniej niż pojedynczy płód, co także przemawia za rolą czynnika rozciągania macicy.
Mechanizm porodowy Skurcze macicy rozpoczynające poród pojawiają się najpierw rzadko i trwają ok. 1 min, następnie ich częstotliwość i intensywność wzrasta i w końcu występują co 1 - 3 min. Równocześnie odruchowo zaczynają się kurczyć mięśnie brzucha wytwarzając tzw. tłocznię brzuszną. W 19 na 20 przypadków płód jest ułożony w jamie macicy w ten sposób, że przy porodzie pierwsza wydostaje się na zewnątrz głowa. Pierwszą przeszkodą, jaką musi pokonać wydalany płód, jest szyjkamacicy. Pod koniec ciąży ulega ona rozpulchnieniu, co pozwala na jej rozciąganie podczas skurczów trżonu macicy. Rozwarcie szyjki macicy następuje zwykle w ciągu 8-24 h w pierwszej ciąży i znacznie krócej, bo niekiedy nawet tylko w kilka minut, w ciążach powtórnych. Jest to pierwszy okres porodu. Drugi okres porodu obejmuje czas przechodzenia płodu przez drogi rodne po rozwarciu szyjki i trwa prawidłowo od jednej minuty u wieloródek do ok. V 2 h lub dłużej u pierwiastek. W tym okresie pękają błony płodowe i odchodzą wody poprzedzające; wody następowe wypływają z chwilą, gdy noworodek opuszcza kanał rodny. Drugi okres porodu kończy się urodzeniem płodu. Skurczom porodowym macicy towarzyszy znaczny ból, spowodowany prawdopodobnie niedokrwieniem mięśniówki macicy wskutek uciskania naczyń krwionośnych przez kurczącą się macicę. Po urodzeniu się noworodka i ustaniu tętnienia pępowiny podwiązuje się ją dwiema mocnymi tasiemkami iprzecina między nimi. Przecięcie pępowiny bez założenia podwiązki spowodowałoby upływ krwi noworodka. Trzeci okres porodu trwa od chwili urodzenia się noworodka do urodzenia się łożyska z błonami płodowymi. Po upływie 10-45 min od urodzenia się noworodka macica obkurcza się, bardzo się przy tym zmniejszając. Następuje '321
T
3 I f -Owodnia
Szyjka macicy
flSt jĘm^S:??, 1*1 P i - i i ^ B M w a M I 1 '
Pochwa
-—-—Wody płodowa
(płyn dniow\)
wm Wtm
Ryc. 154. I okres porodu.
Okres II
Okres I Ryc. 155. Poród.
'322
przy tym oddzielenie łożyska. Towarzyszy temu krwawienie ze stratą ok. 350 ml krwi. Obkurczając się macica zaciska krwawiące naczynia. Po porodzie jama macicy jest podatna na zakażenie, dlatego konieczne jest zachowanie jak największej czystości zarówno podczas porodu, jak i w ciągu najbliższych dni po nim. Okres od ukończenia porodu do całkowitego powrotu narządów rodnych do stanu pierwotnego nazywa się połogiem.
Termin porodu Zakładając, że okres ciąży trwa 280 dni od początku ostatniej miesiączki, datę porodu obliczamy dodając do daty ostatniej miesiączki 7 dni i odejmując trzy miesiące.
UKŁAD KRĄŻENIA
Każda komórka ustroju, aby utrzymać się przy życiu, musi mieć zapewnioną odpowiednią ilość pokarmu i tlenu. Organizmy jednokomórkowe pobierają pożywienie i tlen bezpośrednio z otoczenia. Ustrój człowieka ma wysoko wykształcony układ narządów trawienia, które trawią pokarm, jak również narządów oddechowych, które pobierają tlen z powietrza. W przenoszeniu substancji odżywczych i tlenu z tych układów do tkanek bierze udział krew i limfa, które uczestniczą również w usuwaniu z komórek produktów przemiany materii, przenosząc je do narządów wydalających. Krew krąży w ustroju wzamkniętym układzie naczyń i jest wprawiona w ruch przez serce. Naczynia, którymi płynie krew od serca do obwodu, nazywają się tętnicami (arteriae). Tętnice, początkowo grube, dzielą się na coraz drobniejsze gałązki, które wreszcie rozpadają się na bardzo cienkie naczynia włosowate (kapilary), tworzące w tkankach gęstą sieć. W naczyniach włosowatych krew płynie powoli, co sprzyja wymianie produktów odżywczych. Naczynia włosowate, łącząc się ze sobą, tworzą żyły (venae), którymi krew odpływa do serca.
Ż. główna górna
Przedsionek, prawy
Przedsionek lewy
Pień _ płucny
Ż. główna— dolna
Komora lewa
Ryc. 156. Budowa serca. Strzałki pokazują kierunki przepływu krwi.
'324
Serce składa się z czterech jam, z których dwie noszę nazwę przedsionków (atria) - prawego i lewego, dwie zaś komór (ventriculi) - prawej i lewej. Odpowiednie przedsionki są połączone z komorami, natomiast prawa i lewa połowy serca są od siebie całkowicie oddzielone. Do przedsionka uchodzą żyły, z komór biorą początek tętnice. Z komory lewej (ventriculus sinistei) wychodzi tętnica główna (aorta). Jej gałęzie zaopatrują tkanki ustroju w krew natlenioną i zawierającą substancje odżywcze. Krew po przejściu przez naczynia włosowate odtlenia się i pobiera z tkanek dwutlenek węgla, po czym żyłami dopływa do przedsionka prawego. Drogę krwi z lewej komory do przedsionka prawego nazywamy krwiobiegiem albo krążeniem wielkim. Z przedsionka prawego krew dostaje się do komory prawej, a stąd drogą pnia płucnego i tętnic płucnych - do płuc. W płucach krew oddaje dwutlenek węgla, pobiera tlen i wraca żyłami płucnymi do przedsionka lewego. Drogę krwi z komory prawej do przedsionka lewego nazywamy krążeniem małym lub płucnym. Z powyższego wynika, że przez prawą połowę serca przechodzi krewżylna (odtleniona), przez lewą połowę - krew tętnicza (natleniona). Obieg krwi przedstawia schematycznie ryc. 156. SERCE Serce (cor) jest narządem zbudowanym ze swoistego mięśnia poprzecznie prążkowanego. Ma ono kształt spłaszczonego stożka, podstawą zwróconego ku górze. Wielkość serca równa się w przybliżeniu wielkości pięści osobnika. Dziecko w wieku 2 - 4 lat ma stosunkowo duże serce i wykazuje dużą ruchliwość bez zmęczenia. W okresie pokwitania (14-16 lat) serce jest zbyt małe w stosunku do całego organizmu i przez to słabe; dopiero w wieku 18-20 lat osiąga normę. Serce mężczyzny waży ok. 300 g, kobiety - ok. 270 g.
Kształt serca W sercu wyróżniamy: podstawę zwróconą ku górze, ku tyłowi i ku stronie prawej oraz koniuszek skierowany ku dołowi, przodowi i ku stronie lewej. Na przeponie spoczywa powierzchnia przeponowa serca. Wypukła powierzchnia mostkowo-żebrowa zwrócona jest ku przodowi. Obydwie powierzchnie oddzielone są od siebie brzegiem prawym i leżącą po stronie lewej powierzchnią płucną serca. Na zewnętrznej powierzchni serca znajduje się szereg bruzd, w których leżą naczynia wieńcowe i ich gałęzie, odżywiające mięsień sercowy. Bruzda wieńcowa oddziela przedsionki od komór. Nie ma jej w miejscu odejścia pnia tętnicy płucnej i tętnicy głównej. Bruzdy międzykomorowe, przednia i tylna, leżą na granicy między prawą a lewą komorą. Łączą się one na prawo od '325
Ryc. 157. Widok serca z przodu.
T. główna Ż riłówna górna
Ż. główna dolna T. wieńcowa prawa
Zatoka wieńcowa
Gałąź międzykomorowa tylna
Ryc. 158. Serce od strony tylno-dolnej.
koniuszka, przez co sam koniuszek należy do komory lewej. Powyżej bruzdy wieńcowej znajdują się przedsionki serca. Od każdego z nich odchodzi po stronie bocznej uchyłek o kształcie bardzo spłaszczonego stożka, zwany uszkiem przedsionka. Wewnątrz uszka znajduje się jama komunikująca się z jamą przedsionka. W górnej części przedniej powierzchni serca widzimy główne pnie tętnicze. Skośnie od strony prawej ku lewej i ku górze przebiega pień płucny. Od strony lewej przylega do niego uszko przedsionka lewe, od tyłu graniczy on z aortą. Początkowy odcinek aorty krzyżuje się z pniem płucnym, biegnąc skośnie od strony lewej i dołu ku stronie prawej i ku górze. Nieco od tyłu, góry i strony prawej widoczna jest wpadająca do przedsionka prawego żyła główna górna. Żyła główna dolna wpada do przedsionka prawego bezpośrednio po przejściu przez przeponę. Od tyłu do przedsionka lewego uchodzą trzy lub częściej cztery żyły płucne, Wspomnieliśmy już wyżej o dwóch przedsionkach i dwóch komorach serca. Przedsionek prawy od lewego oddziała przegroda międzyprzedsionkowa, komory zaś - przegroda międzykomorowa. Przedsionek prawy łączy się z komorą prawą ujściem przedsionkowo-komorowym prawym, przedsionek lewy z komorą lewą - ujściem przedsionkowo-komorowym lewym. Ujścia są otoczone zbudowanymi z tkanki łącznej pierścieniami włóknistymi, do których przymocowują się zastawki. W ujściu przedsionkowo-komorowym prawym znajduje się zastawka przedsionkowo-komorowa prawa albo trójdzielna, złożona z trzech płatków; w ujściu przedsionkowo-komorowym lewym zastawka przedsionkowo-komorowa lewa albo dwudzielna, zbudowana z dwóch płatków. Zastawki nie pozwalają na cofanie się krwi do przedsionków w czasie skurczu komór.
Przedsionek prawy Przez ścianę tylną przedsionka prawego (atrium dextrum) uchodzą do niego żyły główne - górna i dolna. W sąsiedztwie ujścia żyły głównej dolnej widzimy ujście zatoki wieńcowej serca (sinus coronarius cordis), zamykane półksiężycowatego kształtu zastawką zatoki wieńcowej (valvula sinus coronarii). Na przedniej ścianie przedsionka leży wejście do uszka prawego. Na powierzchni wewnętrznej uszka widzimy przebiegające zazwyczaj równolegle mięśnie grzebieniaste (musculi pectinati). Na przegrodzie międzyprzedsionkowej znajduje się zagłębienie, zwane dołem owalnym (fossa ova//s). Wżyciu płodowym istnieje w t y m miejscu o t w ó r owalny (foramen ovale), łączący oba przedsionki.
Komora prawa Komora prawa (ventriculus dexter) na przekroju poprzecznym ma kształt półksiężyca. Światło komory zwęża się stopniowo w kierunku koniuszka serca. Ku przodowi od ujścia przedsionkowo-komorowego prawego widzimy począ'327
tek pnia płucnego. W ujściu pnia płucnego widoczna jest zastawka tego pnia, złożona z trzech płatków półksiężycowatych. Każdy z płatków jedną swoją krawędzią przymocowuje się do ściany tętnicy, a brzegiem w o l n y m styka się z płatkami sąsiednimi, zamykając w tej sposób światło tętnicy. Zastawka pnia płucnego (valva trunci pulmonalis) pozwala na swobodne przejście krwi do tętnicy podczas skurczu komory, zamyka natomiast p o w r ó t do komory w czasie jej rozkurczu. Jak w s p o m n i a n o , w ujściu przedsionkowo-komorowym p r a w y m znajduje się zastawka przedsionkowo-komorowa prawa albo trójdzielna (valva tricuspidalis). Każdy z jej płatków ma kształt zbliżony dotrójkąta. Jeden bok płatka jest przytwierdzony do pierścienia włóknistego,otaczającego ujście, dwa pozostałe są wolne. Do wolnych brzegów i dolnej powierzchni każdego płata przymocowane są struny ścięgniste, które d r u g i m końcem łączą się z wierzchołkami mięśni b r o d a w k o w a t y c h . Czynność strun ścięgnistych i mięśni brodawkowatych polega na t y m , że nie pozwalają one na wywinięcie się •> zastawek w kierunku przedsionków podczas skurczu komór. Na ścianie wewnętrznej komory widzimy przebiegające w rozmaitych kierunkach beleczki mięśniowe, z których wychodzą wspomniane wyżej mięśnie brodawkowate. Ich liczba przeważnie odpowiada liczbie płatków zastawek.
Przedsionek lewy Przez o t w o r y tylnej ściany przedsionka lewego ( a t r i u m sinistrum) uchodzą do niego cztery, rzadziej trzy żyły płucne. Uszko lewe jest mniejsze od prawego i, podobnie jak prawe, ma na powierzchni wewnętrznej mięśnie grzebieniaste. W ścianie dolnej przedsionka widzimy ujście przedsionkowo-komorowe lewe.
Komora lewa Komora lewa (ventriculus sinister) jest dłuższa od prawej. Na przekroju poprzecznym ma ona kształt koła. Między przegrodą międzykomorową a ujściem przedsionkowo-komorowym l e w y m bierze początek tętnica główna, W ujściu aorty znajduje się zastawka aorty, zbudowana z trzech półksiężycowatych płatków.
Ujście przedsionkowo-komotówe lewe zamyka zastawka dwudzielna (valva mitralis). Jej struny ścięgniste przymocowane są do dwóch silnie rozwiniętych mięśni brodawkowatych. Na powierzchni wewnętrznej komory lewej znajdują się beleczki mięśniowe. Budowa serca
Ściana serca składa się z trzech w a r s t w ; są t o : 1) nasierdzie (epicardium), 2) mięsień sercowy ( m y o c a r d i u m ) , 3) wsierdzie ( e n d o c a r d i u m ) . Nasierdzie jest cienką błoną surowiczą pokrywającą zewnętrzną powierzchnię mięśnia sercowego wraz z leżącymi na jego powierzchni naczyniami '328
Przedsionek lewy
Zastawka półksięży cowata aorty Odnoga pęczka przedsionkowokomorowego
Zastawka dwudzielna
Mm. brodawkowate
Ryc. 159. Przekrój przez lewę komorę serca.
wieńcowymi. Na początkowych odcinkach wielkich naczyń nasierdzie przechodzi w zewnętrzną blaszkę osierdzia (p. Worek osierdziowy). Mięsień sercowy zbudowany jest z tkanki mięśniowej poprzecznie prążkowanej. Różni się swą budową od mięśni szkieletowych tym, że poszczególne komórki łączą się odgałęzieniami z komórkami leżącymi w sąsiedztwie, tworząc w ten sposób swoistą sieć mięśniową. Włókienka kurczliwe przechodzą przez odgałęzienia do komórek sąsiednich. Wnętrze komórki zajęte jest przez sarkoplazmę z leżącymi w niej jądrami. Na przebiegu włókien mięśniowych widoczne są cienkie, poprzecznie przebiegające, twory, zwane wstawka^ mi. Warstwa mięśnia sercowego jest najgrubsza w obrębie komory lewej, cieńsza w ścianie komory prawej, najcieńsza w przedsionkach. Mięśniówka komór, z wyjątkiem tzw. pęczka przedsionkowo-komorowego (p. niżej), nie łączy się bezpośrednią z mięśniówką przedsionków. Na granicy między przedsionkami i komorami znajdują się pierścienie włókniste. Dwa z nich otaczają ujścia przedsionkowo-komorowe, dwa inne ujście aorty i pnia płucnego. Mięśniówka przedsionków składa się z.dwóch warstw. Warstwa powierzchowna przebiega mniej więcej równolegle do bruzdy wieńcowej serca. Warstwa głęboka jest oddzielna dla każdego przedsionka. Jedne z włókien tej warstwy zaczynają się i kończą na pierścieniach włóknistych, tworząc pętlę. '329
inne gromadzą się okrężnie wokół ujść żył do przedsionków. Mięśniówka komór składa się z trzech warstw. Włókna mięśniowe warstwy powierzchownej zbiegają od pierścieni włóknistych skośnie ku dołowi. Na koniuszku vvłókna zawijają się ósemkowato. W tym miejscu powstaje ich charakterystyczny układ, zwany w i r e m serca. Włókna biegną w dalszym ciągu w kierunku odwrotnym, wchodząc w skład warstwy wewnętrznej ściany komory. Warstwa środkowa jest najgrubsza w komorze lewej. Jej włókna rozpoczynają się i kończą na pierścieniach włóknistych, mając przebieg w kształcie nie zamkniętego koła. W okolicy ujścia żyły głównej do przedsionka prawego, na granicy między przedsionkiem a komorą prawą, oraz w przegrodzie międzykomorowej spotykamy skupienia komórek ubogich w miofibryle, przypominające włókna mięśniowe gładkie. Są to komórki mięśniowe przewodzące serca (tzw. włókna Purkinjego). Pierwsze skupienie włókien tworzy węzeł zatokowo-przedsionkowy, drugie - to węzeł przedsionkowo-komorowy. W przegrodzie międzykomorowej przebiega pęczek przedsionkowo-komorowy. Komórki mięśniowe przewodzące serca tworzą tzw. układ autonomiczny serca, w którym powstają bodźce do jego skurczów. Wsierdzie jest cienką, przezroczystą błoną. Składa się ono z cienkiej warstwy tkanki łącznej, pokrytej warstwą płaskich komórek. Wsierdzie pokrywa ściany przedsionków i komór, przechodząc bezpośrednio w błonę wyściełającą wnętrze naczyń. Zastawki zbudowane są z tkanki łącznej włóknistej, pokrytej po obu stronach przez wsierdzie.
Worek osierdziowy Serce umieszczone jest w worku osierdziowym. Osierdzie (pericardium) składa się z dwóch blaszek: zewnętrznej, czyli ściennej, i z wewnętrznej, czyli trzewnej, lub nasierdzia. Między nimi znajduje się szczelinowata przestrzeń, zawierająca niewielką ilość płynu. Blaszka ścienna tworząca worek wsierdziowy przylega do opłucnej śródpiersiowej, do tylnej powierzchni mostka, do przepony i od tyłu do przełyku. Blaszka trzewna osierdzia (nasierdzie) pokrywa mięsień sercowy wraz z naczyniami wieńcowymi otoczonymi niewielką warstwą tłuszczu, który grupuje się w sąsiedztwie naczyń. Blaszka trzewna przechodzi w blaszkę ścienną na początkowych odcinkach wielkich naczyń serca. W worku osierdziowym znajduje się niewielka ilość płynu, który zapobiega tarciu serca podczas jego pracy.
Położenie serca Serce leży w śródpiersiu; od tyłu sąsiaduje z przełykiem, od przodu przylega na pewnej przestrzeni do ściany klatki piersiowej. Po bokach osierdzie ścienne sąsiaduje z opłucną. Po obu stronach między osierdziem a opłucną przebiegają nerwy przeponowe. '330
Serce leży asymetrycznie, przy czym jego jedna trzecia znajduje się po stronie prawej, zaś dwie trzecie po stronie lewej. Rzut prawego brzegu serca biegnie wzdłuż prawego brzegu mostka łukiem lekko wygiętym w prawo między II prawym międzyżebrzem a przyczepem do mostka chrząstki VI żebra prawego. Rzut lewego brzegu serca biegnie łukiem wypukłym w lewo. Rozpoczyna się w II lewym międzyżebrzu, kończy w miejscu rzutu koniuszka, tj. w V lewym międzyżebrzu, ok. 1,5 cm na prawo od linii środkowo-obojczykowej lewej. Łącząc ten punkt z przyczepem mostkowym chrząstki VI prawego żebra otrzymamy dolną granicę serca. Większą część przedniej powierzchni serca pokrywają płuca, dlatego też do przedniej ściany klatki piersiowej przylega tylko niewielka część serca. W klinice metodą opukiwania można wyznaczyć granice całego serca albo tylko tej jego części, która nie jest przykryta przez płuca. W pierwszym przypadku m ó w i m y o stłumieniu względnym, w drugim o stłumieniu bezwzględnym (tab. 28). T a b e l a 28 Granice serca Granica
Stłumienie bezwzględne
Stłumienie względne
Prawa
linia mostka lewa
linia mostka prawa
Lewa
1,5 cm przyśrodkowo od linii środkowo-obojczykowej lewej
1,5 cm przyśrodkowo od linii środkowo-obojczykowej lewej
Górna
żebro IV lewe
górny brzeg III żebra
Dolna
trudne do oznaczenia ze względu na stłumienie pochodzące od wątroby
Unaczynienie i unerwienie serca Serce unaczyniają tętnice wieńcowe prawa i lewa (aa. coronariae dextra et sinistra). Tętnice wieńcowe odchodzą ód wstępującej części aorty tuż powyżej płatków półksiężycowatych. Leżą one pod nasierdziem, otoczone tkanką tłuszczową. Tętnica wieńcowa prawa biegnie w bruździe wieńcowej między uszkiem a przedsionkiem prawym, dostaje się na tylną powierzchnię serca i w bruździe międzykomorowej tylnej podąża w kierunku koniuszka serca. Tętnica wieńcowa lewa dzieli się na dwie gałęzie: gałąź międzykomorowa przednia kieruje się ku dołowi, biegnąc w bruździe międzykomorowej przedniej; gałąź okalająca biegnie w lewo w bruździe wieńcowej między przedsionkiem lewym a uszkiem. Żyły sęrca uchodzą do zatoki wieńcowej, która biegnie na tylnej powierzchni serca. Żyła wielka serca biegnie początkowo w bruździe międzykomorowej przedniej, zawraca na lewo wzdłuż bruzdy wieńcowej, a końcowy jej odcinek rozszerza się w zatokę wieńcową. Żyła średnia serca biegnie w bruździe międzykomorowej tylnej. Żyła mała serca przebiega w prawej części bruzdy '331
wieńcowej. Nerwy serca pochodzą od nerwu błędnego i pnia współczułnego. Gałęzie tych n e r w ó w tworzą splot sercowy. O znaczeniu tych n e r w ó w dla pracy serca będzfe mowa w rozdz. Fizjologia układu krążenia. NACZYNIA KRWIONOŚNE Powiedzieliśmy już, iż naczynia krwionośne dzielimy na tętnice, żyły i naczynia włosowate. Czynność naczyń w p ł y w a na ich budowę. W tętnicach płynie krew pod dużym ciśnieniem, dlatego ściany ich są grube i sprężyste. Naczynia włosowate mają ściany bardzo cienkie, co jest niezbędne przy w y m i a n i e produktów pomiędzy krwią a tkankami. Grubość ścian żył jest niewielka, gdyż krew płynie w nich pod m a ł y m ciśnieniem.
Tętnice Rozróżnia się tętnice małe, średnie i duże. Przejście między nimi jest stopniowe. W ścianie tętnic małych wyróżniamy trzy w a r s t w y : 1) błonę w e w n ę t r z n ą , 2) błonę środkową i 3) błonę zewnętrzną. Błona wewnętrzna składa się ze śródbłonka i cienkiej warstewki tkanki łącznej. Śródbłonek tworzą płaskie wydłużone komórki, ułożone w jedną warstwę. Nadają one gładkość ścianie wewnętrznej tętnic. Najgrubsza warstwa środkowa zbudowana jest z włókien mięśniowych, przebiegających okrężnie. Błona zewnętrzna składa się z tkanki łącznej, przy czym włókna klejodajne i sprężyste biegną wzdłuż osi długiej naczynia. Tętnice średniego kalibru zbudowane są podobnie. Na granicy między błoną wewnętrzną a środkową oraz środkową a zewnętrzną znajduje się nagromadzenie włókien sprężystych. Ilość włókien sprężystych wzrasta w miarę powiększania się grubości naczynia. Tętnice grube zbudowane są nieco odmiennie. IMa granicy pomiędzy warstwą wewnętrzną a środkową skupia się dużo włókien sprężystych. Znajdują się one również w obrębie w a r s t w y środkowej i zewnętrznej. W ścianach dużych tętnic spotykamy cienkie naczynia, umożliwiające odżywianie grubszych ścian naczyń. Tętnice biorą czynny udział w procesie krążenia. Wskutek skurczu mięśniówki tętnic może się rytmicznie zmieniać ich światło powodując tętnienie naczynia.
Żyły Podobnie jak w przypadku tętnic w skład ściany żył wchodzą trzy w a r s t w y : błona wewnętrzna, błona środkowa i błona zewnętrzna. Błona w e w n ę t r z n a składa się ze śródbłonka i warstewki tkanki łącznej. Błona środkowa zawiera nieliczne włókienka mięśniowe i jest w żyłach bardzo cienka. Błona zewnętrzna - w przeciwieństwie do tętnic - jest najgrubsza. Zawiera ona włókna '332
klejodajne oraz włókna mięśni gładkich o przebiegu podłużnym. Żyły średnie i duże są zbudowane podobnie, z tym że znacznie wzrasta grubość ich warstwy zewnętrznej.
Naczynia włosowate Naczynia włosowate (kapiiary) mają ściankę zbudowaną z jednej warstwy komórek śródbłonka. Na zewnętrznej powierzchni tych naczyń spotyka się komórki z wypustkami (tzw. perycyty). Mają one zdolność kurczenia się, przez to mogą zmieniać światło naczyń. Krew z tętnic do żył nie zawsze płynie przez naczynia włosowate. W pewnych miejscach istnieją bezpośrednie połączenia między nimi (tzw. anastomozy tętniczo-żylne), mające duże znaczenie w regulacji krążenia.
Naczynia tętnicze krążenia wielkiego Tętnica główna (aorta) Tętnica główna (aorta) jest największym naczyniem tętniczym ustroju, średnica jej wynosi przeciętnie 28 mm. Jej odgałęzienia zaopatrują w krew narządy całego ciała. Aorta bierze początek w lewej komorze serca. Jak wspomniano, w miejscu odejścia aorty widnieje zastawka, złożona z trzech płatków, które od swojego kształtu noszą nazwę półksiężycowatych. Tuż ponad nimi aorta jest zgrubiała i to miejsce nosi nazwę opuszki aorty. Tętnicę główną dzielimy na trzy części. Są to: 1) aorta wstępująca, 2) łuk aorty, 3) aorta zstępująca. W części zstępującej wyróżniamy aortę piersiową i aortę brzuszną. Aorta wstępująca kieruje się początkowo ku górze i ku stronie prawej, od przodu sąsiaduje z pniem płucnym. W dalszym przebiegu zagina się ku tyłowi i ku stronie lewej, przechodząc w łuk aorty. Od części wstępującej, tuż powyżej zastawek, biorą początek tętnice wieńcowe serca. Łuk aorty przechodzi ponad lewym oskrzelem głównym i lewą tętnicą płucną, po czym biegnie w kierunku tylnej ściany klatki piersiowej, gdzie na wysokości IV kręgu piersiowego rozpoczyna się część zstępująca aorty. Od łuku aorty odchodzą trzy gałęzie, zaopatrujące w krew kończyny górne i głowę. Są to: a) pień ramienno-głowowy, b) tętnica szyjna wspólna lewa, c) tętnica podobojczykowa lewa. Od wklęsłej powierzchni łuku aorty odchodzą cienkie gałęzie oskrzelowe. Aorta zstępująca ma kształt wydłużonej litery S. Rozpoczyna się ona na wysokości IV kręgu piersiowego, kończy się na poziomie IV kręgu lędźwiowego. Aorta piersiowa (część piersiowa aorty zstępującej) znajduje się powyżej jej przejścia przez przeponę leżącą na wysokości XII kręgu piersiowego. Początkowy i końcowy odcinek aorty piersiowej przebiega po lewej stronie kręgosłupa, środkowy leży na powierzchni przedniej, granicząc od przodu 22 - Anatomia i fizjologia
333
T. szyjna wspólna IPWb
T. podobojczykowa lewa
Pień ramienno -głowowy
Aorta piersiowa
T. nerkown
T. biodrowa wspólna
T. biodrowa zewnętiznn
T. biodrowa wewnętrzna
T. udowa
Ryc. 160. Gałęzie aorty.
z przełykiem. Leżąc na tylnej ścianie klatki piersiowej, aorta wygina się zgodnie z wygięciami kręgosłupa. Od aorty piersiowej odchodzą cienkie gałęzie oskrzelowe, przełykowe, śródpiersiowe oraz 10 par tętnic międzyżebrowych tylnych. Aorta brzuszna biegnie od rozworu tętnicy głównej w przeponie do wysokości IV kręgu lędźwiowego, gdzie dzieli się na naczynia końcowe. Są to: dwie tętnice biodrowe wspólne i nieparzysta tętnica krzyżowa pośrodkowa. Aorta brzuszna biegnie pozaotrzewnowo i sąsiadując z kręgosłupem leży po jego lewej stronie. Na przedniej powierzchni trzonów kręgowych po stronie prawej aorty znajduje się żyła główna dolna. Od przodu aorta graniczy z trzustką i dolną częścią dwunastnicy. Aorta brzuszna oddaje gałęzie ścienne i trzewne. Z ostaną one opisane w odpowiednim rozdziale. Gałęzie aorty dzielimy na:' 1) gałęzie łuku aorty, 2) gałęzie aorty piersiowej, 3) gałęzie aorty brzusznej. '334
Gałęzie łuku aorty Tętnica szyjna wspólna Po stronie prawej tętnica szyjna wspólna (a. carotis communis) odchodzi od pnia ramienno-głowowego, po stronie lewej bezpośrednio od tuku aorty. Tętnica szyjna wspólna prawa powstaje z podziału pnia ramienno-głowowego, poza stawem mostkowo-obojczykowym prawym. Tętnica szyjna wspólna lewa opuszcza otwór górny klatki piersiowej, przechodząc poza stawem mostkowo-obojczykowym lewym. Od tego miejsca stosunki topograficzne obydwu tętnic są jednakowe. Tętnica szyjna wspólna leży na powięzi przykrywającej mięśnie głębokie szyi, od przodu jest przykryta mięśniem mostkowo-obojczykowo-sutkowym. Przyśrodkowo od tętnicy leży u dołu tchawica i przełyk, w górze - krtań i gardło. Bocznie od tętnicy znajduje się żyła szyjna wewnętrzna. Między żyłą a tętnicą przebiega nerw błędny. Na wysokości górnej krawędzi chrząstki tarczowatej tętnica szyjna wspólna dzieli się na dwie gałęzie równej wielkości: tętnicę szyjną zewnętrzną i tętnicę szyjną wewnętrzną.
T. skroniowa powierzchowna T. szczękowa
—T. twarzowa
Ryc. 161. Tętnice głowy i szyi.
'335
Tętnica szyjna zewnętrzna W początkowym odcinku tętnica szyjna zewnętrzna (a. carotis externa) leży przyśrodkowo od żyły szyjnej wewnętrznej, następnie przechodzi na jej stronę boczną. Leżąc w trójkącie tętnicy szyjnej, pokryta jest powięzią, mięśniem szerokim szyi i skórą i tu można wyczuć na niej tętno. Na wysokości szyjki wyrostka kłykciowego żuchwy tętnica szyjna zewnętrzna dzieli się na dwie gałęzie końcowe: tętnicę skroniową powierzchowną i tętnicę szczękową. Tętnica skroniowa powierzchowna (a. temporalis superficialis) jest przedłużeniem tętnicy szyjnej zewnętrznej. Biegnie ku górze, leżąc nieco ku przodowi od otworu słuchowego zewnętrznego. Na wysokości górnego brzegu oczodołu dzieli się na dwie gałęzie: końcowe przednią - czołową i tylną ciemieniową. Tętnica ta często jest widoczna u osób starszych, tętno zaś na niej jest zawsze wyczuwalne. Oddaje ona gałęzie do mięśni twarzy, do ślinianki przyusznej, małżowiny usznej i mięśnia skroniowego. Tętnica szczękowa (a maxillaris) jest drugim końcowym odgałęzieniem tętnicy szyjnej zewnętrznej. Odchodzi od niej pod kątem prostym na wysokości szyjki wyrostka kłykciowego żuchwy. Początkowo biegnie ku przodowi i przyśrodkowo, dalej między mięśniem skrzydłowym bocznym i przyśrodkowym dostaje się do dołu skrzydłowo-podniebiennego. Do ważniejszych odgałęzień tętnicy szczękowej należą: a. Tętnica zębodołowa dolna (a. alveolaris interior) wchodzi do kanału żuchwy i oddaje gałązki do zębów. Zakończenie jej wychodzi przez otwór bródkowy. b. Tętnica zębodołowa górna tylna (a. alveolaris superior posterior) występuje często w postaci kilku gałęzi. Tętniczki te wnikają przez drobne otworki znajdujące się w szczęce unaczyniając zęby trzonowe. Od tętnicy szyjnej zewnętrznej odchodzą również tętnice do tarczycy, języka, gardła, skóry twarzy i okolicy potylicznej. Gałęzie tętnicy szyjnej zewnętrznej 1. Tętnica tarczowa górna (a. thyreoidea superior) odchodzi w pobliżu miejsca podziału tętnicy szyjnej wspólnej i zaginając się łukiem wypukłym ku dołowi osiąga górny brzeg tarczycy. Na jej przebiegu odchodzą odgałęzienia do krtani i mięśnia mostkowo-obojczykowo-sutkowego. 2. Tętnica językowa (a. lingualis) odchodzi od tętnicy szyjnej zewnętrznej na wysokości większego rogu kości gnykowej. Kieruje się ku przodowi i przyśrodkowo, podążając do języka. Odgałęzienia tej tętnicy zaopatrują w krew mięśnie i błonę śluzową języka, częściowo migdałki, nagłośnię i śliniankę podżuchwową. 3. Tętnica twarzowa (a. facialis) odchodzi nieco powyżej tętnicy językowej. Biegnie ku przodowi i ku górze, wchodząc pod mięsień rylcowo-gnykowy i brzusiec tylny mięśnia dwubrzuścowego, następnie przechodzi obok ślinianki podżuchwowej i krzyżuje trzon żuchwy od zewnątrz, na przednim brzegu żwacza. Od tego miejsca wyginając się wężowato osiąga kąt wewnętrzny szpary powiekowej, otrzymując nazwę tętnicy kątowej (a. angularis). Na swym przebiegu oddaje odgałęzienia do warg, okolicznych mięśni, ślinianki podżuchwowej i skóry twarzy. 4. Tętnica gardłowa wstępująca (a. pharyngea ascendens) rozpoczyna się na przyśrodkowej ścianie tętnicy szyjnej zewnętrznej i po bocznej powierzchni ściany gardła wznosi się ku górze, zaopatrując gardło.
'336
5. Tętnica potyliczna (a. occipitalis) odchodzi od tylnej powierzchni tętnicy szyjnej zewnętrznej; kierując się ku tyłowi i ku górze, leży w bruździe tętnicy potylicznej na kości skroniowej. Unaczynia ona okoliczne tkanki i skórę aż do ciemienia. 6. Tętnica uszna tylna (a. auricularis posterioi) rozpoczyna się nieco powyżej tętnicy potylicznej i biegnie ku tyłowi między wyrostkiem sutkowatym a małżowiną uszną, oddając odgałęzienia do kanału nerwu twarzowego, jamy bębenkowej, komórek sutkowych i małżowiny usznej. W szczęce unaczynia zęby trzonowe szczęki oraz błonę śluzową zatoki szczękowej. 7.Tętnica podoczodołowa (a. infraorbitalis) przez szczelinę oczodołową dolną przenika do oczodołu, a następnie przez bruzdę i otwór podoczodołowy wychodzi na przednią powierzchnię twarzy. Unaczynia ona przednie zęby szczęki i okolicę podoczodołową. Mniejsze odgałęzienia wysyła do przewodu słuchowego zewnętrznego, jamy bębenkowej, mięśnia żuchwowo-gnykowego, mięśnia żwacza, mięśni skrzydłowych, mięśnia skroniowego, tylnej części ściany jamy nosowej i dolnych mięśni poruszających gałkę oczną.
Tętnica szyjna wewnętrzna Tętnica szyjna wewnętrzna (a. carotis interna) od miejsca podziału tętnicy szyjnej wspólnej kieruje się ku podstawie czaszki. Przez kanał tętnicy szyjnej (canaiis caroticus) dostaje się ona do wnętrza czaszki, gdzie biegnie ku przodowi obok siodła tureckiego, przez zatokę jamistą. Nieco ku tyłowi od wyrostka pochyłego przedniego zagina się ku górze, przebija oponę twardą i wchodzi na podstawę mózgowia. Odchodzą od niej następujące gałęzie: 1. Tętnica oczna przez kanał wzrokowy dostaje się do oczodołu. Licznymi gałęziami unaczynia gałkę oczną, mięśnie i gruczołłzowy. Jej końcowe gałązki wychodzą przez otwór nadoczodołowy na zewnątrz, unaczyniając okolicę nadoczodołową. 2. Tętnica przednia mózgu biegnie po przyśrodkowej powierzchni półkul mózgowych równolegle do ciała modzelowatego. Tętnice prawa i lewa są zespolone tętnicą łączącą przednią. 3. Tętnica środkowa mózgu wchodzi do dołu bocznego mózgu, oddając gałęzie do płatów skroniowego i ciemieniowego. Tętnica łącząca tylna zespala tętnicę środkową mózgu z tętnicą tylną mózgu. Tętnice mózgu przednie i tylne (p. niżej) oraz tętnice łączące tworzą na podstawie mózgu koło tętnicze mózgu. Poniżej zestawiono gałęzie tętnicy szyjnej wspólnej. • T. szyjna wspólna T. szyjna zewnętrzna T. T. T. T. T. T. T.
tarczowa górna językowa twarzowa gardłowa wstępująca potylięzna uszna tylna szczękowa
T. szyjna wewnętrzna T. oczna T. p r a d n i a mózgu T. środkowa mózgu T. łącząca tylna
Tt. zębodołowe 22 - Anatomia i fizjologia
337
Tętnica podobojczykowa Tętnica podobojczykowa (a. subclavia) prawa uchodzi, od pnia ramienno-głowowego, lewa zaś bezpośrednio od łuku aorty. Wskutek tego tętnica podobojczykowa prawa jest nieco krótsza od lewej. Wychodzi ona z otworu górnego klatki piersiowej, kieruje się nieco ku tyłowi od szczytu płuca, leżąc na osklepku opłucnej. Razem ze splotem ramiennym przechodzi między mięśniami pochyłymi, przylegając do I żebra. W dalszym odcinku tętnica podobojczykowa przechodzi bez wyraźnej granicy w tętnicę pachową. Od tętnicy podobojczykowej odchodzą następujące gałęzie: 1. Tętnica kręgowa (a. vertebralis) podąża ku t y ł o w i , po czym przez o t w o r y w wyrostkach poprzecznych kręgów szyjnych biegnie ku górze, zagina się dookoła tylnego łuku kręgu szczytow e g o i przez o t w ó r wielki kości potylicznej dostaje się do wnętrza czaszki. Tu na przedniej powierzchni rdzenia przedłużonego łączy się z tętnicą kręgową strony przeciwnej, tworząc tętnicę podstawną mózgu {a. basilariś). Tętnica kręgowa oddaje odgałęzienia do mięśni szyi, do rdzenia kręgowego i jego opon, rdzenia przedłużonego oraz tętnicę dolną tylną móżdżku.
2. Tętnica piersiowa wewnętrzna (a. thoracica interna) odchodzi od tętnicy podobojczykowej w pobliżu tętnicy kręgowej i podąża ku dołowi wewnątrz klatki piersiowej poza chrząstkami żebrowymi, równolegle do krawędzi mostka. Na wysokości VI międzyżebrza dzieli się na dwie gałęzie końcowe: tętnice mięśniowo-przeponową i nabrzuszną górną. Pierwsza z nich oddaje odgałęzienia do dolnych przednich międzyżebrzy, druga unaczynia górną część
- T. podpbojczykowa Pień ramienno-głowowy
Ryc. 162. Tętnica podobojczykowa, pachowa i ramienna.
'338
mięśnia prostego brzucha, łącząc się z tętnicą nabrzuszną dolną. Od pnia tętnicy piersiowej wewnętrznej odchodzą tętnice międzyżebrowe przednie do sześciu górnych międzyżebrzy. Tętnice te zespalają się z tętnicami międzyżebrowymi tylnymi. 3. Pień tarczowo-szyjny (truncus thyreocervicalis), długości ok. 1 cm, dzieli się na następujące gałęzie: a) tętnica tarczowa dolna unaczynia krtań i częściowo tarczycę; b) tętnica szyjna wstępująca, odchodząca często od poprzedniej, unaczynia mięśnie pochyłe i oddaje odgałęzienia do rdzenia; c) tętnica szyjna powierzchowna oddaje odgałęzienia do mięśnia czworobocznego i unaczynia skórę bocznej powierzchni szyi; d) tętnica nadłopatkowa biegnie poza obojczykiem. Unaczynia skórę tej okolicy, mięśnie nadgrzebieniowy i podgrzebieniowy oraz oddaje odgałęzienia do sieci barkowej. 4. Pień żebrowo-szyjny (truncus costocervicalis) oddaje dwie gałęzie: a) tętnicę karkową głęboką, unaczyniającą głębokie mięśnie szyi; oddaje ona odgałęzienia rdzeniowe do dwóch otworów międzykręgowych odcinka szyjnego kręgosłupa; b) tętnicę międzyżebrową najwyższą, która biegnie ku dołowi i tyłowi, oddając tętnice międzyżebrowe tylne do dwóch górnych międzyżebrzy. 5. Tętnica poprzeczna szyi przechodzi między pęczkami splotu ramiennego w kierunku przyśrodkowego kąta łopatki. Unaczynia mięśnie grzbietu i karku oraz skórę okolicy karkowej (ryc. 161). Tętnica pachowa Tętnica pachowa (a. axillaris) jest przedłużeniem tętnicy podobojczykowej. Biegnie ona od wysokości górnego brzegu pierwszego żebra do miejsca skrzyżowania z dolnym brzegiem ścięgna mięśnia piersiowego większego, gdzie przechodzi w tętnicę ramienną (a. brachialis). Tętnica pachowa leży w dole pachowym, poza mięśniami piersiowymi większym i mniejszym. Przyśrodkowo i ku przodowi od niej znajduje się żyła pachowa. Nerw pośrodkowy obejmuje tętnicę dwoma pęczkami. Od tętnicy pachowej odchodzą: 1. Tętnica piersiowa najwyższa, rozgałęziająca się w dwóch górnych zębach mięśnia zębatego przedniego i mięśniach międzyżebrowych. 2. Tętnica piersiowo-barkowa (a. thoracoacromialis) oddaie odgałęzienia do mięśni piersiowych, mięśnia ramiennego i do sieci barkowej. 3. Tętnica piersiowa boczna biegnie ku dołowi, unaczyniając mięsień zębaty przedni. 4. Tętnica podłopatkowa kieruję się ku tyłowi i dołowi i dzieli się na gałąź piersiowo-grzbietową, która unaczynia mięsień najszerszy i mięsień zębaty przedni, oraz na tętnicęokalającąłopatkę, która przechodzi przez otwór trójboczny i oddaje odgałęzienia do mięśnia podgrzebieniowego i podłopatkowego. 5. Tętnice okalające ramię przednia i tylna. Pierwsza z nich otacza szyjkę kości ramiennej od przodu i oddaje odgałęzienia do mięśnia ramiennego i torebki stawu ramienneao. Tętnica okalająca ramię tylna otacza od tyłu szyjkę chirurgiczną kości ramiennej, przechodzi przez otwór czworoboczny i rozgałęzia, się w mięśniu naramiennym.
'339
Schemat rozgałęzień tętnicy podobojczykowej i tętnicy pachowej T. pachowa
T. podobojczykowa T. kręgowa T. piersiowa wewnętrzna T. mięśniowo-przeponowa T. nadbrzuszna górna Pień tarczowo-szyjny T. tarczowa dolna T. szyjna wstępująca T. podłopatkowa Pień żebrowo-szyjny T. karkowa głęboka T. międzyżebrowa najwyższa T. poprzeczna szyi
T. T. T. T.
piersiowa najwyższa piersiowo-barkowa piersiowa boczna podłopatkowa T. piersiowo-grzbietowa T. okalająca łopatkę T. okalająca ramię przednia T. okalająca ramię tylna
Tętnica ramienna Tętnica ramienna (a. brachialis) jest przedłużeniem tętnicy pachowej. Leżąc w bruździe przyśrodkowej ramienia, podąża do zgięcia łokciowego, gdzie dzieli się na tętnicę łokciową i promieniową. Tętnicy ramiennej towarzyszą dwie żyły ramienne i nerw pośrodkowy, który początkowo leży bocznie od tętnicy ramiennej, ku dołowi zaś przechodzi na jej powierzchnię przyśrodkową. Od tętnicy ramiennej odchodzą następujące gałęzie: 1. Tętnica głęboka ramienia odchodzi od tętnicy ramiennej na wysokości dolnej krawędzi mięśnia obłego większego. Razem z nerwem promieniowym przebiega w bruździe nerwu promieniowego między głową boczną a przyśrodkową mięśnia trójgłowego. Tętnica głęboka ramienia oddaje odgałęzienia do mięśnia trójgłowego oraz tętnice wchodzące w skład sieci stawowej łokcia. 2. Tętnica promieniowa (a. radialis) jest niejako przedłużeniem tętnicy ramiennej. Odchodzi od niej pod rozcięgnem mięśnia dwugłowego ramienia, przedostaje się na przednio-boczną powierzchnię przedramienia i kieruje się ku dołowi, biegnąc równolegle do mięśnia ramienno-promieniowego. W dolnym odcinku przedramienia leży na kości promieniowej, pokryta jedynie skórą, co umożliwia wyczucie tętna w t y m miejscu. W dalszym ciągu tętnica promieniowa pod ścięgnami odwodziciela długiego kciuka, prostownika krótkiego kciuka i prostownika długiego kciuka przez tzw. tabakierkę anatomiczną przedostaje się na grzbietową powierzchnię ręki, po czym między I a II kością śródręcza dostaje się na dłoń, gdzie przechodzi w łuk dłoniowy głęboki (arcus palmaris profundus). Na swym przebiegu tętnica promieniowa oddaje gałęzie mięśniowe do okolicznych mięśni, tętnicę do sieci stawowej łokcia i gałęzie do sieci nadgarstka. Do ważniejszych rozgałęzień tętnicy promieniowej należą: a) gałąź dłoniowa powierzchowna, która odchodzi od tętnicy promieniowej przed jej przejściem na grzbietową powierzchnię ręki i przyczynia się do utworzenia powierzchownego łuku dłoniowego;
'340
b) tętnica główna kciuka, która odchodzi od końcowego odcinka tętnicy promieniowej i dzieli się na trzy gałęzie, zwane tętnicami palcowymi własnymi; unaczyniają one obie krawędzie powierzchni dłoniowej kciuka i krawędź boczną palca wskazującego; c) tętnice grzbietowe śródręcza, które łączą się zespoleniami z łukiem dłoniowym głębokim i unaczyniają powierzchnie grzbietowe palców.
Ryc. 163. Tętnice przedramienia.
3. Tętnica łokciowa (a. ulnaris) jest drugą dużą końcową gałęzią tętnicy ramiennej. Przechodzi ona przez dół łokciowy i pod mięśniami przyczepiającymi się do kłykcia przyśrodkowego kości ramiennej kierując się ku dołowi po przednio-przyśrodkowej powierzchni przedramienia, równolegle do mięśnia zginacza łokciowego nadgarstka. Po stronie bocznej kości grochowatej oddaje gałąź dłoniową głęboką, która łączy się z końcem tętnicy promieniowej. '341
tworząc łuk dłoniowy głęboki. Tętnica łokciowa oddaje drobne gałęzie do okolicznych mięśni, do sieci łokciowej, sieci nadgarstka. Do ważniejszych odgałęzień naczyń należę: a) tętnica międzykostna wspólna, która odchodzi od tętnicy łokciowej w górnym jej odcinku; krótki pień tętnicy międzykostnej wspólnej dzieli .się na tętnice międzykostne przednią i tylną, które biegną odpowiednio na przedniej i tylnej powierzchni błony międzykostnej i oddają odgałęzienia do okolicznych mięśni oraz do sieci dłoniowej i grzbietowej nadgarstka; b) gałąź dłoniowa głęboka wchodzi w skład łuku dłoniowego głębokiego; c) gałąź dłoniowa powierzchowna bierze udział w budowie łuku dłoniowego powierzchownego.
Końcowe gałęzie tętnicy promieniowej i tętnicy łokciowej łącząc się ze sobą tworzą tętnicze łuki dłoniowe powierzchowny i głęboki. Łuk dłoniowy powierzchowny (arcus palmaris superficialis) utworzony jest przez gałąź dłoniową powierzchowną, odchodzącą od tętnicy prorhieniowej, która łączy się z pełnym światłem gałęzi dłoniowej powierzchownej, odchodzącej od tętnicy łokciowej. Od łuku odchodzą tętnice palcowe wspólne, które w okolicy głów kości śródręcza dzielą się na tętnice palcowe własne. Biegną one do bocznych krawędzi palców. Jak wspomniano, kciuk i boczna strona palca wskazującego otrzymują gałęzie od tętnicy głównej kciuka. Łuk dłoniowy głęboki (arcus palmaris profundus) utworzony jest z gałęzi dłoniowej głębokiej tętnicy łokciowej i tętnicy dłoniowej głębokiej, odchodzącej od tętnicy promieniowej. Łuk dłoniowy głęboki leży na wysokości podstaw kości śródręcza. Oddaje cztery tętnice śródręcza dłoniowe. Łączą się one zespoleniami z grzbietowymi tętnicami śródręcza, po czym uchodzą do tętnic palcowych wspólnych. Istnienie łuków dłoniowych umożliwia sprawne krążenie krwi w obrębie ręki pomimo czasowego zahamowania dopływu krwi do poszczególnych jej części. Schemat rozgałęzień tętnicy ramiennej i
T. ramienna
T. promieniowa ^ Gałąź dłoniowa powierzchowna T. główna kciuka Tt. grzbietowe śródręcza
T. głęboka ramienia
i
+
i
|
T. łokciowa T. międzykostna wspólna Gałąź dłoniowa głęboka Gałąź dłoniowa powierzchowna
Gałęzie aorty zstępującej Odgałęzienia aorty piersiowej Od aorty piersiowej odchodzą gałęzie trzewne i ścienne. Cienkie gałęzie trzewne biegną do oskrzeli, przełyku i do osierdzia. Do gałęzi ściennych należą: '342
a) gałęzie śródpiersiowe i tętnice przeponowe górne, unaczyniające lędźwiową część przepony, b) tętnice międzyżebrowe tylne, występujące w liczbie 10 par, ponieważ górne dwa międzyżebrza zaopatrywane są przez gałęzie tętnicy międzyżebrowej najwyższej. Każda z tętnic międzyżebrowych dzieli się nieco dalej na gałąź grzbietową i gałąź poboczną. Gałąź poboczna biegnie w odpowiednim międzyżebrzu i w okolicy kąta żebrowego dzieli się na dwie gałęzie, z których jedna przebiega w rowku żebra leżącego wyżej, druga - ponad krawędzią żebra leżącego niżej. Tętnice te zespalają się z tętnicami międzyżebrowymi przednimi, odchodzącymi od tętnicy piersiowej wewnętrznej. Gałąź grzbietowa tętnicy międzyżebrowej tylnej unaczynia mięśnie i skórę grzbietu, a przez otwory międzykręgowe oddaje odgałęzienia do rdzenia kręgowego, opon rdzenia i kręgów. Tętnice międzyżebrowe unaczyniają mięśnie międzyżebrowe, oddają odgałęzienia do sąsiednich mięśni klatki piersiowej i jamy brzusznej oraz do gruczołów sutkowych i do skóry odpowiedniej okolicy. Gałęzie aorty brzusznej Naczynia odchodzące od aorty brzusznej dzielimy na gałęzie ścienne i gałęzie trzewne. Gałęzie ścienne aorty brzusznej 1. Tętnica przeponowa rozgałęzia się na dolnej powierzchni przepony i oddaje tętnicę nadnerczową górną. 2. Tętnice lędźwiowe, w liczbie czterech, biegną poprzecznie między mięśniami brzucha. Unaczyniają one mięśnie ściany jamy brzusznej oraz oddają odgałęzienia do rdzenia i mięśni grzbietu. 3. Tętnica krzyżowa pośrodkowa jest cienką gałązką biegnącą w przedłużeniu aorty brzusznej. Gałęzie trzewne nieparzyste aorty brzusznej I. Pień trzewny (truncus celiacus) odchodzi od przedniej ściany aorty na wysokości XII kręgu piersiowego. Krótki, długości 1 cm, pień dzieli się na trzy gałęzie: 1) tętnicę wątrobową wspólną, 2) tętnicę żołądkową lewą, 3) tętnicę śledzionową. 1. Tętnica w ą t r o b o w a wspólna (a. hepatica communis) po oddzieleniu się od pnia trzewnego biegnie ku stronie prawej i dalej jako tętnica wątrobowa właściwa wchodzi do więzadła wątrobowo-dwunastniczego; następnie podążając do wrót wątroby biegnie wraz z przewodem żółciowym współnym i żyłą wrotną. Tętnica wątrobowa wspólna ma następujące odgałęzienia: a) tętnica żołądkowo-dwunastnicza podąża ku dołowi i dzieli się na tętnicę żołądkowo-sieciową prawą i tętnicę trzustkowo-dwunastniczą górną: '343
A) tętnica żołądkowo-sieciowa prawa biegnie ku stronie lewej po krzywiźnie większej żołądka, unaczyniając tę okolicę oraz oddając odgałęzienia do sieci większej; końcem swym łączy się z tętnicą żołądkowo-sieciową lewą; B) tętnica trzustkowo-dwunastnicza górna zaopatruje w krew górną część dwunastnicy oraz głowę i trzon trzustki, zespalając się z tętnicą trzustkowo-dwunastniczą dolną; b) tętnica żołądkowa prawa wchodzi na krzywiznę mniejszą żołądka od strony odźwiernika, biegnie ku stronie lewej i zespala się z tętnicą żołądkową lewą, oddaje odgałęzienia do sąsiednich okolic żołądka; c) tętnica wątrobowa właściwa podąża do wnęki wątroby, gdzie dzieli się na gałąź prawą i lewą, rozgałęziające się w odpowiednich płatach wątroby. Od gałęzi prawej odchodzi również tętniczka do pęcherzyka żółciowego. 2. Tętnica żołądkowa lewa (a. gastrica sinistra) po oddzieleniu się od pnia trzewnego wchodzi od strony lewej na krzywiznę mniejszą żołądka. Łączy się ona z tętnicą żołądkową prawą, tworząc łuk biegnący wzdłuż krzywizny mniejszej żołądka. 3. Tętnica śledzionowa (a. lienalis) po odejściu od pnia trzewnego biegnie poza żołądkiem ku stronie lewej,tworzącliczne wygięcia.Tętnica śledzionowa oddaje po drodze odgałęzienia do żołądka i do trzustki i wpada do wnęki śledziony, dzieląc się na kilka gałęzi. Od tętnicy śledzionowej odchodzi tętnica żołądkowo-sieciowa lewa, która przebiega wzdłuż lewej części krzywizny większej żołądka i łączy się z tętnicą żołądkowo-sieciową prawą, tworząc łuk tętniczy wzdłuż krzywizny większej żołądka. II. Tętnica krezkowa górna (a. mesenteńca superior) odchodzi od przedniej powierzchni aorty brzusznej, 1 - 2 cm poniżej pnia trzewnego. Dzieli się ona wielokrotnie, zaopatrując w krew cienkie jelita i znaczną część jelita grubego. Gałęzie tętnicy zespalają się ze sobą łukowatymi połączeniami, tworzącymi tzw. arkady. Wielka liczba arkad ułatwia krążenie oboczne w przypadku zahamowania lub utrudnienia dopływu krwi do pewnego odcinka jelita. Tętnica krezkowa górna wychodzi spod dolnego brzegu trzustki i wnika do korzenia krezki, w której podąża do miejsca połączenia się jelita cienkiego z jelitem grubym. Gałęzie tętnicy krezkowej biorą nazwy od zaopatrywanych przez siebie odcinków przewodu pokarmowego. Są one następujące: 1. Tętnica trzustkowo-dwunastnicza dolna biegnie do głowy trzustki i dolnego odcinka dwunastnicy, łączy się zespoleniem z tętnicą trzustkowo-dwunastniczą górną. 2. Tętnice jelita czczego i krętego, w liczbie 12 - 15, unaczyniają jelita cienkie. 3. Tętnica krętniczo-okrężnicza unaczynia jelito ślepe i wyrostek robaczkowy. 4. Tętnica okrężnicza prawa rozgałęzia się w części wstępującej okrężnicy. 5. Tętnica okrężnicza środkowa unaczynia okrężnicę poprzeczną oraz łączy się zespoleniami z tętnicą okrężniczą lewą, pochodzącą od tętnicy krezkowej dolnej (ryc. 160). '344
III. Tętnica krezkowa dolna (a. mesenterica interior) odchodzi od aorty na poziomie III kręgu lędźwiowego i, podobnie jaktętnica krezkowa górna, tworzy liczne arkady. Zaopatruje ona okrężnicę zstępującą, esowatą i częściowo odbytnicę. Tętnica krezkowa dolna ma następujące gałęzie: P1. Tętnica okrężnicza lewa unaczynia okrężnicę zstępującą. 2. Tętnice esicze, w liczbie dwóch do pięciu, unaczyniają okrężnicę esowatą oraz tworzą zespolenia z tętnicą okrężniczą lewą i odbytniczą górną. 3. Tętnica odbytnicza górna unaczynia górny odcinek odbytnicy. Zespala się gałęziami z tętnicą odbytniczą środkową, odchodzącą od tętnicy biodrowej wewnętrznej. Gałęzie trzewne parzyste aorty brzusznej 1. Tętnice nadnerczowe środkowe odchodzą od aorty brzusznej nieco powyżej poziomu tętnicy krezkowej górnej i biegną poprzecznie do nadnerczy. 2. Tętnice nerkowe (aa. renales) są najgrubsze z parzystych naczyń trzewnych. Odchodzą od aorty brzusznej pod kątem prostym na wysokości II kręgu lędźwiowego i biegną ku bokowi i nieco ku dołowi. Tętnica nerkowa prawa jest dłuższa i przechodzi poza żyłą główną dolną. Oddaje ona drobne odgałęzienia do nadnercza i tkanki okołonerkowej, dzieli się na kilka gałęzi, które wnikają przez wnękę do miąższu nerki. 3. Tętnice jądrowe lub jajnikowe, zaopatrujące gruczoły płciowe, odchodząc nieco poniżej tętnic nerkowych, biegną ku dołowi i ku bokowi, krzyżują się z moczowodem i osiągają wysokość kresy łukowatej. W dalszym ciągu u mężczyzn tętnice jądrowe podążają przez kanał pachwinowy do jąder, a u kobiet tętnice jajnikowe kierują się przyśrodkowo, wchodzą do więzadła szerokiego macicy, zaopatrują w krew jajniki i oddają odgałęzienia do macicy. Gałęzie końcowe aorty brzusznej Wspomniano wyżej, że aorta brzuszna na wysokości IV kręgu lędźwiowego dzieli się na dwie tętnice biodrowe wspólne. Kierują się one w bok i ku dołowi. Na wysokości stawu krzyżowo-biodrowego tętnica biodrowa wspólna dzieli się na tętnice: biodrową wewnętrzną i zewnętrzną. Tętnica biodrowa wewnętrzna Tętnica biodrowa wewnętrzna (a. iliaca interna) ma długość kilku centymetrów. Na wysokości górnego brzegu otworu zasłonionego dzieli się na gałąź przednią i tylną. Od gałęzi tylnej biorą początek następujące tętnice: 1. Tętnica biodrowo-lędźwiowa unaczynia mięsień lędźwiowy większy i mniejszy. 2. Tętnica krzyżowa boczna przez otwory krzyżowe miednicze oddaje gałęzie rdzeniowe.
'345
3. Tętnica pośladkowa górna przez otwór kulszowy większy ponad mięśniem gruszkowatym zdąża do mięśni pośladkowych.
Gałąź przednia tętnicy biodrowej wewnętrznej zaopatruje trzewa miednicy małej, a więc pęcherz moczowy, odbytnicę, macicę i narządy płciowe zewnętrzne. Jej gałęzie zaopatrują również mięśnie miednicy i mięśnie przywodzące uda. Od przedniej gałęzi tętnicy biodrowej wewnętrznej odchodzą następujące ważniejsze tętnice: 1. Tętnica pępkowa obok bocznej ściany pęcherza wchodzi na przednią ścianę jamy brzusznej. Jest drożna jedynie w życiu płodowym, następnie zanika. 2. Tętnica maciczna, dość gruba gałąź, osiąga macicę na granicy jej trzonu i szyjki, przebiegając w więzadle szerokim macicy, zagina się w kierunku dna macicy. Oddaje gałęzie przebiegające w licznych skrętach do ściany macicy i drobne odgałęzienia do jajnika, jajowodu i pochwy. 3. Tętnica pośladkowa dolna przez otwór podgruszkowy przedostaje się na zewnątrz miednicy małej. Unaczynia mięśnie znajdujące się w miednicy małej i mięśnie pośladkowe, szczególnie mięsień pośladkowy wielki.
Tętnica biodrowa zewnętrzna Tętnica biodrowa zewnętrzna (a. iiiaca externa) leży w przedłużeniu tętnicy biodrowej wspólnej i kieruje się ku dołowi i ku bokowi, sąsiaduje od strony bocznej z mięśniem lędźwiowym większym. Przechodzi przez rozstęp naczyń na udo, gdzie leży po bocznej stronie żyły udowej. Od skrzyżowania z więzadłem pachwinowym tętnica biodrowa zewnętrzna przybiera nazwę tętnicy udowej. Schemat gałęzi aorty piersiowej
Pień trzewny T. wątrobowa wspólna
T. żołądkowo-dwunastnicza
T. żołądkowo-sieciowa prawa
Tt. nerkowe
T. okrężnicza lewa
T. odbytnicza górna
Na swym przebiegu tętnica biodrowa zewnętrzna oddaje: 1. Tętnicę nadbrzuszną dolną, która po zewnętrznej powierzchni mięśnia prostego brzucha kieruje się ku górze i tworzy zespolenie z tętnicą nadbrzuszną górną (gałęzie tętnicy piersiowej wewnętrznej). 2. Liczne cienkie odgałęzienia, z których jedno, zwane tętnicą okalającą biodra, biegnie równolegle do więzadła pachwinowego.
'346
T. b i o d r o w a — wspólna T. b i o d r o w a — } wewnętrzna t
T. u d o w a -
T. głęboka uda—
T. u d o w a
i \
T. p o d k o l a n o w a
T. piszczelowa przednia T. piszczelowa tylna T. strzałkowa
Ryc. 164. T ę t n i c e k o ń c z y n y d o l n e j .
Tętnica udowa Tętnica udowa (a. femoralis) jest przedłużeniem tętnicy biodrowej zewnętrznej. Początkowy odcinek tętnicy leży w obrębie trójkąta udowego, pokryty blaszką powierzchowną powięzi szerokiej uda, sąsiaduje bocznie z nerwem udowym i przyśrodkowo z żyłą udową. W tym miejscu ze względu na powierzchowne położenie daje się na niej łatwo wyczuć tętno. W dalszym przebiegu tętnica udowa leży na mięśniach przywodzących udo, sąsiadując od strony bocznej z mięśniem obszernym przyśrodkowym, wreszcie przez kanał przywodzicieli przechodzi na tylną stronę uda i dostaje się do dołu podkolanowego. W początkowym odcinku od tętnicy udowej odchodzą: 1. Gałęzie od ściany brzucha i narządów płciowych zewnętrznych. '347
2. Tętnica głęboka uda (a. profunda femoris) odchodzi od tylnej powierzchni tętnicy udowej, podąża ku dołowi i ku tyłowi, leżąc pomiędzy mięśniami obszernymi przyśrodkowym a przywodzicielami. Odchodzą od niej tętnice okalające udo: a) tętnica okalająca udo przyśrodkowa okrąża kość udową od strony przyśrodkowej, poniżej przyczepu mięśnia biodrowo-lędźwiowego i oddaje odgałęzienia do okolicznych mięśni oraz panewki stawu biodrowego; b) tętnica okalająca udo boczna przechodzi pod mięśniem krawieckim w bok i dzieli się na dwie gałęzie: wstępującą i zstępującą. Gałąź wstępująca między mięśniami prostym uda a obszernym pośrednim biegnie ku górze i w kierunku bocznym, gdzie tworzy liczne zespolenia z tętnicą okalającą udo przyśrodkową, oddając gałęzie do mięśnia naprężacza powięzi szerokiej i mięśnia pośladkowego wielkiego; gałąź zstępująca rozgałęzia się w mięśniach prostujących udo. W dalszym ciągu tętnica głęboka uda oddaje trzy tętnice przeszywające, które przechodzą na tylną powierzchnię uda i unaczyniają mięśnie zginające goleń.
Dwie główne tętnice uda, tj. tętnica udowa i tętnica głęboka uda, połączone są ze sobą licznymi zespoleniami, dzięki czemu istnieją tu dobre warunki do powstania krążenia obocznego. W kanale przywodzicieli tętnica udowa oddaje drobne odgałęzienia do okolicznych mięśni i do sieci tętniczej stawu kolanowego. Tętnica podkolanowa Tętnica podkolanowa (a. poplitea) od wyjścia z kanału przywodzicieli leży na dnie dołu podkolanowego; nieco bardziej powierzchownie znajduje się żyła podkolanowa i nerw kulszowy. Pęczek naczyniowo-nerwowy pokrywa warstwa tkanki tłuszczowej i z tego powodu w dole podkolanowym trudno jest wyczuć tętno. Tętnica podkolanowa oddaje odgałęzienia do okolicznych mięśni, torebki stawu kolanowego i jego więzadeł oraz do sieci stawowej kolana. W dolnej części dołu podkolanowego tętnica podkolanowa dzieli się na tętnicę piszczelową przednią i tylną (ryc. 164). Tętnice goleni i stopy 1. Tętnica piszczelowa przednia (a. tibialis anterior) przez otwór w błonie międzykostnej dostaje się na przednią powierzchnię, leżąc głęboko po bocznej stronie mięśnia piszczelowego przedniego. W dalszym ciągu tętnica przechodzi pod ścięgnem prostownika długiego palucha na grzbiet stopy, gdzie przybiera nazwę tętnicy grzbietowej stopy. Wspólnie z tętnicą piszczelową przednią biegną dwie żyły oraz nerw strzałkowy głęboki. Na swym przebiegu tętnica piszczelowa przednia oddaje odgałęzienia do sieci stawowej kolana, mięśni goleni oraz sieci kostki bocznej i przyśrodkowej. Tętnica grzbietowa stopy biegnie powierzchownie między ścięgnami mięśnia prostownika długiego palców. Na grzbiecie stopy między I a II kością śródstopia można wyczuć na niej tętno. Na wysokości podstawy I kości śródstopia tętnica grzbietowa stopy dzieli się na gałąź podeszwową głęboką i tętnicę śródstopia pierwszą. Gałąź podeszwowa głęboka przechodzi
'348
między I a II kością śródstopia na stronę podeszwową stopy, gdzie łączy się z końcowym odcinkiem tętnicy podeszwowej bocznej. Tętnica śródstopia pierwsza oddaje odgałęzienia do palucha i bocznej powierzchni palca drugiego. Od tętnicy grzbietowej stopy odchodzą ponadto cienkie tętnice stępowe przyśrodkowe i grubsza tętnica stępowa boczna, która swoim końcem zespala się z tętnicą łukowatą. Tętnica łukowata odchodzi od tętnicy grzbietowej stopy obok gałęzi podeszwowej głębokiej, biegnie łukiem wypukłym ku przodowi po podstawach kości śródstopia, oddaje tętnice grzbietowe śródstopia, z których każda dzieli się na dwie tętnice grzbietowe palców.
2. Tętnica piszczelowa tylna (a tibialis posterior) biegnie w przedłużeniu tętnicy podkolanowej ku tyłowi od błony międzykostnej, razem z nerwem piszczelowym, po czym, owijając się do tyłu koło kostki przyśrodkowej i pod troczkiem mięśni zginaczy, dostaje się na podeszwę. Tu dzieli się na gałęzie końcowe: tętnicę podeszwową przyśrodkową i tętnicę podeszwową boczną. Poza kostką przyśrodkową tętnica piszczelowa tylna leży powierzchownie, dlatego możliwe jest wyczucie na niej tętna. W górnym odcinku tętnica piszczelowa tylna oddaje cienką tętnicę strzałkową, przebiegającą wzdłuż strzałki ku dołowi i unaczyniającą boczną grupę mięśni goleni i boczną okolicę pięty. Tętnica piszczelowa tylna oddaje również odgałęzienia, unaczyniającetylną grupę mięśni goleni, odpowiednią okolicę skóry, oraz odgałęzienia do sieci kostki bocznej, do kostki przyśrodkowej i do pięty. Tętnica podeszwową przyśrodkową unaczynia mięśnie palucha, jej gałąź głęboka łączy się często z łukiem tętniczym podeszwy. Tętnica podeszwową boczna biegnie pod mięśniem zginaczem krótkim palców w kierunku podstawy V kości śródstopia. Zawracając w kierunku przyśrodkowym, łączy się swoim końcem z gałęzią podeszwową głęboką, odchodzącą od tętnicy grzbietowej stopy, i tworzy łuk podeszwowy. Od łuku podeszwowego odchodzą tętnice podeszwowe śródstopia. Od nich odłączają tętnice podeszwowe wspólne palców, z których każda dzieli się na dwie tętnice własne palców.
Schemat rozgałęzień tętnicy biodrowej T. biodrowa
t. nadbrzuszna dolna
T. udowa
t. głęboka uda t. okrążająca udo t. przeszywające
T. podkolanowa
t. piszczelowa przednia
t. piszczelowa tylna t. strzałkowa
t. grzbietowa stopy t. podeszwową głęboka t. śródstopia I
t. podeszwową przyśrodkową t. podeszwową boczna
'349
Naczynia żylne krążenia wielkiego Żyły krążenia wielkiego dzielimy na dwie grupy. Pierwsza z nich obejmuje żyły głębokie towarzyszące tętnicom, druga - żyły biegnące niezależnie ód tętnic, żyły skórne. W większości przypadków mniejszym tętnicom obwodow y m towarzyszą dwie żyły, grubszym zaś pniom tętniczym, np. tętnicy udowej i tętnicy pachowej - jedna żyła. Żyły skórne, niekiedy bardzo dobrze rozwinięte, mają liczne połączenia z żyłami głębokimi, co bardzo ułatwia krążenie oboczne. Żyły krążenia wielkiego dzielimy na kilka układów, a mianowicie: 1) układ żyły głównej górnej, 2) układ żyły głównej dolnej, 3) układ żył serca, 4) układ żyły wrotnej, który, aczkolwiek włączony jest do układu żyły głównej dolnej, ze względu na szczególny charakter został z niego wyodrębniony.
Żyła główna górna Żyła główna górna (v. cava superior) powstaje na wysokości chrząstki I prawego żebra z połączenia żył ramienno-głowowych prawej i lewej. Po około 6-centymetrowym przebiegu uchodzi ona do przedsionka prawego serca. Żyła główna górna sąsiaduje od przodu z grasicą, od strony prawej z opłucną śródpiersiową. Wpadają do niej żyły śródpiersiowe przednie, żyły osierdziowe oraz dość gruby pień żyły nieparzystej. Żyła główna górna nie ma zastawek. Zbiera ona krew ze ścian klatki piersiowej, głowy i szyi oraz z kończyn górnych. Rozpatrzmy kolejno jej dopływy. Żyła nieparzysta krótka Żyła nieparzysta krótka powstaje z połączenia żyły lędźwiowej wstępującej lewej z żyłą podżebrową lewą. Przechodzi ona do klatki piersiowej pod łukiem lędźwiowo-żebrowym po stronie lewej. Na wysokości VII—IX kręgu piersiowego zbacza ku stronie prawej, przechodzi przed kręgosłupem i wpada do żyły nieparzystej. Przyjmuje ona krew z dolnych żył międzyżebrowych lewych tylnych bezpośrednio oraz z górnych lewych żył międzyżebrowych za pośrednictwem żyły nieparzystej krótkiej dodatkowej, która może bezpośrednio uchodzić do żyły nieparzystej, jak również łączyć się z żyłą ramienno-głowową lewą.
Żyły ramienno-głowowe Żyły ramienno-głowowe prawa i lewa ( w . brachiocephalicae) powstają poza odpowiednim stawem mostkowo-obojczykowym z połączenia żyły szyjnej wewnętrznej i żyły podobojczykowej. Żyła ramienno-głowowa prawa jest krótsza i ma przebieg prawie pionowy, lewa, dłuższa, biegnie skośnie od strony lewej i górnej na prawo i ku dołowi. Żyła ramienno-głowowa leWa od przodu sąsiaduje z głównymi odgałęzieniami aorty. Do żył ramienno-głowo'350
wych dochodzą drobne żyły z narządów śródpiersia tylnego. Do większych dopływów tych żył należą: 1. Żyła kręgowa, która, powstając u podstawy czaszki ze splotu żylnego pod potylicznego, kieruje się ku dołowi przez otwory w wyrostkach poprzecznych kręgów szyjnych i po wyjściu z nich zagina się ku przodowi i wpada do żyły ramienno-głowowej. 2. Żyła karkowa głęboka zbiera krew z mięśni głębokich szyi i mięśni karku. 3. Żyła piersiowa wewnętrzna biegnie równolegle do zewnętrznego brzegu mostka w odległości 1 cm po wewnętrznej powierzchni klatki piersiowej. Otrzymuje ona krew z żył międzyżebrowych przednich i żyły nadbrzusznej górnej. 4. Żyły międzyżebrowe łączą żyły piersiowe wewnętrzne z żyłą nieparzystą po stronie prawej oraz z żyłami parzystymi - krótką i krótką dodatkową - po stronie lewej. Żyła międzyżebrowa najwyższa zbiera krew z dwóch do czterech górnych międzyżebrzy.
Żyły głowy i szyi Żyła szyjna wewnętrzna (v. jugularis interna) powstaje z żył mózgu i opon mózgowych, opuszcza czaszkę przez otwór szyjny i, zdążając ku dołowi, leży na tętnicy szyjnej wewnętrznej, a następnie obok tętnicy szyjnej wspólnej.
Ryc. 165. Żyły głowy i szyi.
'351
Łącznie z wymienionymi naczyniami przebiega również nerw błędny; jak wspomniano, poza stawem mostkowo-obojczykowym żyła szyjna wewnętrzna łączy się z żyłą podobojczykową tworząc żyłę ramienno-głowową. Dopływy żyły szyjnej wewnętrznej dzielimy na grupy w zależności od okolic, z których pochodzą. Wyróżniamy więc żyły powłok czaszki, żyły powierzchowne i głębokie twarzy, żyły ścian kostnych czaszki i żyły mózgu. Ważniejszym dopływem żyły szyjnej wewnętrznej jest żyła szyjna zewnętrzna. Powstaje ona kilka centymetrów poniżej otworu słuchowego zewnętrznego z połączenia żyły potylicznej, żyły usznej tylnej i z naczynia zespalającego z żyłą zażuchwową. Od tego miejsca żyła szyjna zewnętrzna kieruje się ku dołowi biegnąc po przedniej krawędzi mięśnia mostkowo-obojczykowo-sutkowego. Uchodzą od niej żyła szyjna przednia, żyły poprzeczne szyi, a niekiedy również żyła nadłopatkowa. Do żyły szyjnej wewnętrznej odpływa krew z zatok żylnych opony twardej mózgowia. Zatoki żylne są to szerokie, o sztywnych ścianach kanały leżące wzdłuż sierpu mózgu, sierpu móżdżku i na wewnętrznej powierzchni podstawy czaszki (p. Opony mózgowia). Najważniejsze z nich to zatoka strzałkowa górna i zatoka strzałkowa dolna przebiegające wzdłuż krawędzi sierpu mózgu. Od tyłu łączy je zatoka prosta. Obok siodła tureckiego leżą zatoki jamiste. Na wewnętrznej powierzchni łuski kości potylicznej przebiega zatoka poprzeczna. Przechodzi ona ku bokowi w zatokę esowatą, opuszczającą czaszkę przez otwór szyjny.
Żyły kończyny górnej Żyły powierzchowne kończyny górnej 1. Żyła odpromieniowa (v. cephalica) powstaje z drobnych żył powierzchni grzbietowej nadgarstka, kieruje się ku górze, przechodzi na przednio-boczną powierzchnię przedramienia. W dalszym ciągu biegnie w bruździe bocznej mięśnia dwugłowego ramienia i między mięśniem naramiennym a piersiow y m większym, osiąga dół podobojczykowy, gdzie zagina się ku tyłowi i uchodzi do żyły pachowej. Na wysokości stawu łokciowego dochodzi do żyły odpromieniowej żyła pośrodkowa łokciowa (v. mediana cubiti). Tworzy ona połączenie między żyłami odpromieniową i odłokciową. Żyła pośrodkowa łokciowa jest miejscem, w którym najczęściej wykonuje się wstrzyknięcia dożylne. 2. Żyła odłokciową (v. basilica) powstaje na tylno-bocznej powierzchni przedramienia, od strony przyśrodkowej przechodzi na jego przednią powierzchnię i w połowie ramienia wpada do żyły ramiennej. Żyły głębokie kończyny górnei Żyły głębokie kończyny górnej towarzyszą tętnicom. Każdej tętnicy palców towarzyszą dwie żyłki. Łączą się one ze sobą po dwie, tworząc na grzbiecie ręki cztery żyły grzbietowe śródręcza. Żyły śródręcza łączą się licznymi odgałęzie'352
niami i dają początek żyłom odpromieniowej i odłokciowej. Żyły na powierzchni dłoniowej ręki są bardzo słabo rozwinięte. Wpadają one do podwójnych żylnych łuków dłoniowych - powierzchownego i głębokiego. Tętnicy promieniowej i łokciowej towarzyszą po dwie żyły. Żyły te w obrębie stawu łokciowego łączą się w dwie żyły towarzyszące tętnicy ramiennej. Do przyśrodkowej
Ż. odłokciowa
-Ż. pośrodkowa łokciowa
ł
Ż. odpr
Ryc. 166. Żyły przedramienia i ręki.
żyły ramiennej wpada żyła odłokciowa. Żyły towarzyszące tętnicy ramiennej leżą w bruździe przyśrodkowej mięśnia dwugłowego razem z tętnicą ramienną. Na wysokości dolnego brzegu mięśnia piersiowego większego żyły te łączą się w żyłę pachową (v. axillaris), która leży ku przodowi i przyśrodkowo od tętnicy ramiennej. Do żyły pachowej uchodzą żyły ze ściany klatki piersiowej, barku i okolicznych mięśni. Zarówno żyły powierzchowne kończyny górnej, jak głębokie mają zastawki. Więcej zastawek jest w żyłach głębokich. 23 - Anatomia i fizjologia
353
Ryc. 167. Żyły ramienia.
Żyła główna dolna Żyła główna dolna (v. cava interior) jest najgrubszym naczyniem organizmu ludzkiego. Powstaje ona z dwóch żył biodrowych wspólnych, łączących się na wysokości chrząstki między IV i V kręgiem lędźwiowym, nieco poniżej i na prawo od miejsca podziału aorty brzusznej. Żyła główna leży poza otrzewną po prawej stronie tętnicy głównej, przechodzi przez miąższ wątroby w bruździe żyły głównej. Przez otwór w środku ścięgnistym przepony dostaje się do śródpiersia, po czym uchodzi do prawego przedsionka serca. Ku przodowi do żyły głównej dolnej leży końcowy odcinek dwunastnicy i głowa trzustki. Dopływy żyły głównej dolnej dzielimy na ścienne i trzewne. Dopływy ścienne żyły głównej dolnej 1. Żyły lędźwiowe, w liczbie czterech par, biegną w kierunku poprzecznym, zbierając krew ze ścian jamy brzusznej. Lewe są dłuższe niż prawe. Żyły lędźwiowe każdej strony połączone są ze sobą znaną nam już żyłą lędźwiową wstępującą, która, łącząc się z żyłą podżebrową, daje początek żyle nieparzystej po stronie prawej, bądź też nieparzystej krótkiej po stronie lewej. 2. Żyły przeponowe zbierają krew z przepony. '354
Dopływy trzewne żyły głównej dolnej 1. Żyła jądrowa lub jajnikowa prawa. U mężczyzn żyła jądrowa biegnie wspólnie z powrózkiem nasiennym, tworząc w nim splot wiciowaty. Żyły splotu wiciowatego łączą się stopniowo ze sobą i przechodzą przez pierścień pachwinowy głęboki do jamy brzusznej. Tu jako jednolity pień żyła jądrowa biegnie pozaotrzewnowo ku górze i uchodzi pod kątem ostrym do żyły głównej dolnej, nieco poniżej ujścia żyły nerkowej. Żyła jądrowa lewa uchodzi do żyły nerkowej lewej. U kobiet żyła jajnikowa rozpoczyna się kilkoma odgałęzieniami zbierającymi krew z jajnika. Żyły te tworzą splot wiciowaty, który leży w więzadle szerokim macicy. Po opuszczeniu miednicy małej przebieg żył jajnikowych jest taki sam, jak jądrowych u mężczyzn. 2. Żyły nerkowe (w. renales) biorą początek z połączenia kilku naczyń, wychodzących z wnęki nerkowej. Żyła nerkowa prawa jest krótsza, lewa dłuższa, krzyżuje się ona od przodu z aortą brzuszną. Do żyły nerkowej lewej uchodzi żyła jądrowa lub jajnikowa i żyła nadnerczowa lewa. Czasem żyła nadnerczowa lewa, podobnie jak prawa, uchodzi bezpośrednio do żyły głównej dolnej. Żyły biodrowe wspólne Żyły biodrowe wspólne (w. iliacae communes) powstają z żyły biodrowej zewnętrznej i biodrowej wewnętrznej na wysokości stawu krzyżowo-biodrowego odpowiedniej strony. Żyła biodrowa wspólna prawa jest krótsza od lewej. Leży ona ku tyłowi, a następnie z boku od tętnicy biodrowej wspólnej prawej. Żyła biodrowa wspólna lewa leży przyśrodkowo od tętnicy o tej samej nazwie i przechodzi poza tętnicę biodrową wspólną prawą. Miejsce połączenia żyły biodrowej wspólnej prawej i lewej znajduje się na prawo i ku dołowi od rozdwojenia tętnicy głównej. Do żyły biodrowej wspólnej lewej wpada żyła krzyżowa pośrodkowa, która zbiera krew ze splotu krzyżowego. 1. Żyła biodrowa wewnętrzna (v. iliaca interna) ma postać krótkiego, grubego pnia, który biegnie po bocznej ścianie miednicy małej, ku tyłowi od tętnicy biodrowej wewnętrznej. Dopływy żyły biodrowej wewnętrznej dzielą się na dopływy trzewne i dopływy ścienne. Występują one najczęściej w postaci naczyń podwójnych i zbierają krew z okolic zaopatrywanych przez tętnice biodrowe. Dopływy trzewne żyły biodrowej wewnętrznej biorą początek ze splotów żylnych narządów miednicy małej; są to: żyły odbytnicze środkowe i żyły pęcherzowe. Ponadto u mężczyzn do żyły biodrowej wpadają żyły ze splotu żylnego sterczowego, a u kobiet żyły ze splotu macicznego. Dopływy ścienne żyły biodrowej wewnętrznej zbierają krew ze ścian miednicy małej i pośladka. Zalicza się do nich żyły pośladkowe górne i dolne, żyły biodrowo-lędźwiowe i żyły krzyżowe boczne. 2. Żyła biodrowa zewnętrzna (v. iliaca externa) jest przedłużeniem żyły udowej. Przechodzi ona przez rozstęp mięśni pod więzadłem pachwinowym '355
i wpada do żyły biodrowej wspólnej. Żyła biodrowa zewnętrzna niekiedy ma zastawki. Wpada do niej żyła nabrzuszna dolna. Żyła nabrzuszna dolna biegnie równolegle do tętnicy nabrzusznej dolnej na tylnej powierzchni przedniej ściany brzucha. Żyła nabrzuszna dolna tworzy liczne zespolenia z żyłą nabrzuszną górną i żyłami przypępkowymi. Żyły kończyny dolnej Z kończyny dolnej krew odpływa żyłami powierzchownymi i głębokimi. Żyły te mają liczne zastawki. Największą zżył powierzchownych jest żyła odpiszćzelowa (v. saphena magna). Bierze ona początek z drobnych żył na powierzchni
Ryc. 168. Żyły kończyny dolnej.
'356
grzbietowej stopy, po czym po przyśrodkowej powierzchni goleni podąża ku górze. Okrążając od tyłu kłykieć przyśrodkowy kości udowej, biegnie na przyśrodkowej, a następnie przedniej powierzchni uda, wreszcie w obrębie otworu owalnego wpada do żyły udowej. Na swoim przebiegu żyła odpiszczelowa przyjmuje wiele dopływów z przyśrodkowej i tylnej powierzchni goleni oraz z przyśrodkowej, przedniej i tylnej powierzchni uda. Żyła odpiszczelowa ma szereg połączeń z żyłami głębokimi kończyny dolnej. Żyła odstrzałkowa (v. saphena parva) bierze początek z drobnych żył grzbietowej powierzchni stopy, poza kostką boczną dostaje się na tylną powierzchnię goleni. W dole podkolanowym przebija powięź i wpada do żyły podkolanowej. Żyła odstrzałkowa ma liczne zespolenia z żyłami głębokimi. Żyły głębokie kończyny dolnej rozpoczynają się na grzbietowej powierzchni stopy i na podeszwie, przy czym każdej tętnicy towarzyszą dwie żyły. Z podwójnych żył piszczelowych przednich i tylnych w dolnym kącie dołu podkolanowego powstaje żyła podkolanowa. Uchodzą do niej żyły okolicy stawu kolanowego oraz żyła odstrzałkowa. W dole podkolanowym żyła podkolanowa sąsiaduje od tyłu z nerwem piszczelowym, od przodu z tętnicą podkolanową. Z dołu podkolanowego żyła przechodzi do kanału przywodzicieli, przybierając tu nazwę żyły udowej. Leżąc przyśrodkowo od tętnicy udowej, przechodzi pod więzadłem pachwinowym przez rozstęp naczyń do jamy brzusznej, gdzie przybiera nazwę żyły biodrowej zewnętrznej. Do żyły udowej wpadają żyły głębokie uda, które przyjmują krew z żył przebijających i żył okalających udo przyśrodkowych i bocznych, z mniejszych żył dolnego odcinka jamy brzusznej, z narządów płciowych zewnętrznych, oraz wspomniana żyła odpiszczelowa (ryc. 168). Układ żyły wrotnej Żyła wrotna (v. portae) prowadzi krew z brzusznej części przewodu pokarmowego, śledziony i trzustki do wątroby. Żyła wrotna powstaje w okolicy głowy trzustki przez połączenie żyły śledzionowej z żyłą krezkową górną. Jej krótki (ok. 5 cm) pień przebiega wewnątrz więzadła wątrobowo-dwunastniczego ku tyłowi od tętnicy wątrobowej i od przewodu żółciowego wspólnego. We wnęce wątroby żyła wrotna dzieli się na gałąź prawą i lewą, które unaczyniają odpowiednie płaty wątroby. Krew żyły wrotnej po przejściu przez układ żył międzyzrazikowych i śródzrazikowych (p. Budowa wątroby) dostaje się do żył wątrobowych, a stąd do żyły głównej dolnej. Układ żyły wrotnej nie ma zastawek. Żyła wrotna ma następujące dopływy: 1. Żyła krezkowa górna zbiera krew z odcinków przewodu pokarmowego, zaopatrywanych przez tętnicę krezkową górną, a zatem z jelita ślepego, wyrostka robaczkowego, z okrężnicy wstępującej i poprzecznej, z jelita cienkiego. Poza t y m do żyły krezkowej górnej uchodzą: żyła żołądkowo-sieciowa prawa i żyły trzustkowo-dwunastnicze. '357
2. Żyła śledzionowa powstaje z gałęzi wychodzących z wnęki śledziony, po czym kieruje się ku stronie prawej, leżąc nieco poniżej tętnicy śledzionowej. Do żyły śledzionowej wpada żyła żołądkowo-sieciowa lewa, zbierając krew z krzywizny większej żołądka i sieci większej, drobne żyły z trzustki i żołądka oraz żyła krezkowa dolna, zbierająca krew z okrężnicy zstępującej i esowatej oraz z górnej części odbytnicy. Żyła krezkowa dolna czasami uchodzi bezpośrednio do żyły wrotnej, w miejscu połączenia żyły krezkowej górnej z żyłą śledzionową. 3. Bezpośrednio do żyły wrotnej uchodzą: żyły żołądkowe prawa i lewa biegnące po jego krzywiźnie mniejszej oraz odgałęzienia od pęcherzyka żółciowego i z więzadła obłego wątroby. Połączenia żyły wrotnej z układami żyły głównej górnej i dolnej W chorobach, w których utrudniony jest odpływ przez żyłę wrotną, krew płynie bezpośrednio do żyły głównej górnej lub dolnej przez drobne połączenia. Istnieją trzy główne połączenia żyły wrotnej z żyłami głównymi: 1. Żyły krzywizny mniejszej żołądka łączą się z żyłami przełyku, które uchodzą do żył nieparzystej i nieparzystej krótkiej, wpadających do żyły głównej górnej. 2. Żyła krezkowa dolna przez żyłę odbytniczą górną ma połączenie ze splotem odbytniczym, który przez żyły dobytnicze środkową i dolną łączy się również z układem żyły biodrowej wewnętrznej. Żyła biodrowa wewnętrzna przez żyłę biodrową wspólną łączy się z żyłą główną dolną. 3. Żyławrotnaprzezdrobneżyłyprzypępkowełączysięzżyłaminadbrzusza górną i dolną. Żyła nadbrzusza dolna należy do układu żyły głównej dolnej.
Naczynia krążenia małego Z prawej komory serca wychodzi pień płucny (fruncus pulmonalis). Kieruje się on ku górze i ku stronie lewej. Po około 6-centymetrowym przebiegu pień płucny dzieli się na dwie tętnice p ł u c n e - p r a w ą i lewą (a. pulmonalis dextra et sinistra). Tętnica płucna prawa, nieco dłuższa, biegnie poza wstępującą częścią aorty i poza żyłą główną górną, po czym uchodzi do wnęki płuca prawego poniżej oskrzela, biegnącego do płata górnego. Tętnica płucna lewa leży ku przodowi od części zstępującej aorty i uchodzi do płuca w obrębie wnęki powyżej oskrzela. W miejscu podziału pnia płucnego między nim a aortą ciągnie się więzadło tętnicze. Istnieje ono jako pozostałość po przewodzie tętniczym (ductus arteriosus), którym w okresie płodowym płynęła krew z pnia płucnego do tętnicy głównej. Tętnice płucne prawa i lewa dzielą się na gałęzie odpowiadające liczbie płatów płuca, a następnie przechodzą przez coraz drobniejsze odgałęzienia aż do naczyń włosowatych, oplatających pęcherzyki płucne. '358
Z wnęki każdego płuca wychodzą dwie żyły płucne (w. pulmonales), które po krótkim przebiegu uchodzą do lewego przedsionka serca. Tętnicami płucnymi płynie krew odtleniona (żylna), żyłami krew natleniona (tętnicza).
Krążenie płodowe U płodu krążenie krwi przebiega w sposób odmienny niż u człowieka dorosłego. Wynika to z odmiennych warunków życia płodu, który tlen i składniki odżywcze pobiera z krwi matki. Proces wymiany produktów przemiany materii między krwią matki a krwią płodu następuje w łożysku. Natleniona i zaopatrzona w produkty odżywcze krew z łożyska odpływa do żyły pępkowej. Żyła pępkowa biegnie do pierścienia pępkowego płodu, wchodząc w skład pępowiny. W dalszym ciągu żyła pępkowa podąża do wątroby leżąc w w o l n y m brzegu więzadła sierpowatego i przebiega w bruździe żyły pępkowej, gdzie dzieli się na dwie gałęzie. Jedna z gałęzi, tzw. przewód żylny (ductus venosus), łączy się z żyłą główną dolną, druga łączy się z żyłą wrotną. Krew z żyły wrotnej przez żyły wątrobowe dostaje się do żyły głównej. W ten sposób krew tętnicza, która płynie z łożyska, miesza się z krwią żylną płodu. Z żyły głównej dolnej krew dostaje się do przedsionka prawego serca i dzięki odpowiednio ukształtowanej zastawce żyły głównej dolnej zostaje w większości skierowana przez otwór owalny przegrody międzyprzedsionkowej do przedsionka lewego. Pozostała część krwi miesza się z krwią płynącą z żyły głównej górnej i odpływa do komory prawej, a stąd do pnia płucnego. Płuca płodu są nieczynne i zapotrzebowanie ich na krew jest niewielkie. Dlatego też z pnia płucnego większość krwi płynie do tętnicy głównej przez przewód tętniczy (ductus arteriosus), który łączy pień płucny z aortą. Odgałęzieniem tętnicy głównej rozchodzi się krew po całym ciele. Od tętnic biodrowych wewnętrznych odchodzą dwie tętnice pępkowe (aa. umbilicales). Biegną one po tylnej powierzchni przedniej ściany jamy brzusznej. Przez pierścień pępkowy i pępowinę dostają się do łożyska, gdzie dzielą się na naczynia włosowate. Z powyższego wynika, że tętnicami pępkowymi płynie krew mieszana, żyłą pępkową zaś - krew tętnicza. Po urodzeniu, wraz z pierwszym wdechem noworodka, krew zaczyna krążyć również po obiegu małym. Przewód żylny zamyka się, z czasem zarasta i przekształca się w więzadło żylne. Po zarośniętym otworze owalnym w przegrodzie międzyprzedsionkowej pozostaje ślad w postaci dołu owalnego. Tętnice pępkowe również zarastają, tracąc swe znaczenie. UKŁAD CHŁONNY Naczynia krwionośne włosowate nie stykają się bezpośrednio ze wszystkimi komórkami narządów. Płyny ustrojowe, znajdujące się w przestrzeniach międzykomórkowych, zapewniają ciągłość środowiska i umożliwiają przeno'359
szenie produktów odżywczych do komórek i produktów przemiany materii z komórek. Z przestrzeni międzykomórkowych płyny tkankowe dostaję się do naczyń chłonnych, które odprowadzają je do dwóch dużych przewodów: przewodu piersiowego (ductus thoracicus) oraz do przewodu chłonnego prawego (ductus lymphaticus dexter). Układ chłonny, w przeciwieństwie do układu krwionośnego, jest układem otwartym, tzn. drobne naczynia chłonne otwierają się bezpośrednio do przestrzeni międzykomórkowych. Naczynie
chłonne odprowadzające
1
/
Naczynia chłonne doprowadzające Ryc. 169. Budowa węzła i naczynia chłonnego.
Ściany naczyń chłonnych budową przypominają żyły. Naczynia drobne mają ściany zbudowane z jednej warstwy śródbłonka. Ściana naczyń większych zbudowana jest z błony wewnętrznej, środkowej i zewnętrznej. Zastawki naczyń chłonnych są rozmieszczone o wiele gęściej niż w naczyniach żylnych. Wygląd zewnętrzny drobnych naczyń chłonnych różni się od wyglądu żył następującymi po sobie rozszerzeniami i zwężeniami, co przypomina wyglądem paciorki. W miejscach zwężeń znajdują się zastawki. W skład układu chłonnego poza naczyniami wchodzą również węzły chłonne. Węzłami chłonnymi (nodi lymphatici) nazywamy twory o kształcie zbliżonym do ziarenka fasoli, o średnicy od 2 do 30 mm. Węzeł chłonny pokrywa torebka, z której w głąb narządu wnikają beleczki łącznotkankowe. Zrąb węzła chłonnego tworzą liczne komórki tkanki siateczkowatej. W okach siateczki tkwią skupienia limfocytów, tzw. grudki chłonne. Grupują się one w obwodowej części węzła chłonnego, wchodząc w skład jego kory. Wewnątrz poszczególnych grudek chłonnych znajdują się młodsze pokolenia limfocytów, tzw. limfoblasty. '360
Część wewnętrzną węzła chłonnego zajmuje rdzeń, w którym widoczne są pasma rdzenne, odpowiadające grudkom chłonnym na obwodzie. Pomiędzy miąższem a torebką oraz pomiędzy miąższem a beleczkami łącznotkankowymi znajdują się wolne przestrzenie chłonne, którymi przepływa chłonka. Węzły chłonne pełnią bardzo ważną rolę. Są one miejscem, w którym powstają limfocyty, tu również zostają zatrzymywane i niszczone bakterie płynące z prądem chłonki. Również w węzłach chłonnych zatrzymywane zostają ciała stałe, które dostały się do chłonki. Węzły chłonne układają się przeważnie w grupy od kilku do kilkunastu sztuk. Grudki chłonne w błonie śluzowej jelita noszą nazwę mieszków i budową przypominają grudki chłonne w więzadłach. W jelicie czczym i krętym spotykamy mieszki chłonne samotne. W odcinku końcowym jelita krętego mieszki chłonne samotne łączą się w większe zespoły, tworząc mieszki chłonne skupione. Układ chłonny jest prawdopodobnie ściśle związany z jamami surowiczymi. Łączność ta jest możliwa dzięki drobnym otworkom leżącym pomiędzy komórkami śródbłonka. Chłonka (lympha) jest płynem lekko żółtawej barwy, o składzie chemicznym i biologicznym podobnym do składu osocza krwi. Od krwi różni się brakiem krwinek czerwonych i znaczną zawartością limfocytów, których ilość w 1 mm 3 wynosi od 2000 do 20 000.
Naczynia i więzy chłonne kończyny dolnej Naczynia chłonne kończyny dolnej dzielimy na powierzchowne i głębokie. Naczynia powierzchowne leżą w tkance podskórnej całej kończyny. Największe z nich towarzyszą żyle odpiszczelowej i odstrzałkowej. Naczynia biegnące obok żyły odstrzałkowej uchodzą do węzłów chłonnych podkolanowych. Naczynia towarzyszące żyle odpiszczelowej wpadają do węzłów pachwinowych. Węzły pachwinowe, w liczbie 20-30, leżą w trójkącie udowym, poniżej więzadła pachwinowego. Głębokie naczynia chłonne zbierają limfę z mięśni, powięzi i okostnej. Mają one liczne połączenia z naczyniami powierzchownymi i wpadają do węzłów podkolanowych i pachwinowych. Do tych ostatnich poza naczyniami chłonnymi głębokimi i powierzchownymi kończyny dolnej uchodzą również naczynia z okolicy pośladka, podbrzusza i zewnętrznych narządów płciowych. Zwęzłów pachwinowych odchodzą naczynia zdążające pod więzadłem pachwinowym do jamy brzusznej, gdzie uchodzą do węzłów chłonnych biodrowych wewnętrznych.
'361
! Naczynia i węzły chłonne jamy brzusznej i miednicy małej Naczynia chłonne jamy brzusznej uchodzą do w ę z ł ó w biodrowych wewnętrznych, które leżą w sąsiedztwie tętnicy biodrowej, do węzłów lędźwiowych, leżących po obydwu stronach aorty brzusznej, i do zespołu węzłów trzewnych, leżących w okolicy pnia trzewnego. Naczynia chłonne głębokie narządów płciowych zewnętrznych oraz naczynia narządów płciowych wewnętrznych u mężczyzn i macicy u kobiet oraz naczynia odbytnicze uchodzą do węzłów chłonnych lędźwiowych. Również do węzłów chłonnych lędźwiowych uchodzi duża część naczyń chłonnych ściany jamy brzusznej. Naczynia chłonne nerek i trzustki uchodzą do węzłów trzewnych leżących w sąsiedztwie pnia trzewnego. Naczynia chłonne wątroby dzielą się na powierzchowne i głębokie. Pierwsze
Ryc. 170. S c h e m a t g ł ó w n y c n n a c z y ń c h ł o n n y c h .
'362
i
z nich w większości wpadają do Węzłów leżących we wnęce wątroby, pozostałe przez przeponę dostają się do węzłów śródpiersiowych przednich. Naczynia chłonne śledziony wpadają do węzłów trzustkowo-śledzionowych, połączonych z węzłami trzewnymi. Naczynia chłonne jelit mają szczególny charakter. Zawartość tych naczyń stanowi tzw. mlecz (chylus), w skład którego wchodzi duża ilość emulgowanych tłuszczów. Naczynia te rozpoczynają się ślepo w kosmkach jelitowych, po czym biegną w krezce, wpadają do węzłów krezkowych i do węzłów trzewnych. Naczynia odprowadzające z węzłów krezkowych uchodzą również do węzłów trzewnych. Naczynia biegnące od węzłów trzewnych tworzą pień jelitowy (truncus intestinalis). Łączy się on z dwoma pniami lędźwiowymi (trunci lumbales), powstałymi z naczyń odprowadzających węzłów lędźwiowych. Naczynia te wchodzą do rozszerzonego zbiornika mleczu (cisterna chyli). Od tego zbiornika bierze początek przewód piersiowy. Przewód piersiowy (ductus thoracicus) jest głównym naczyniem chłonnym ustroju. Zbiera on chłonkę z górnej lewej połowy ciała oraz całej części dolnej. Jak już wspomniano, rozpoczyna się on od zbiornika mleczu, po czym biegnie ku górze, leżąc po prawej stronie aorty, i wraz z nią przechodzi do klatki piersiowej. Tu przebiega na przedniej powierzchni kręgosłupa, nieco na prawo od linii środkowej ciała. Na wysokości IV-V kręgu piersiowego odgina się ku przodowi i ku stronie lewej i uchodzi do lewego kąta żylnego, to jest w miejscu połączenia żyły podobojczykowej lewej z żyłą szyjną wewnętrzną. Do przewodu piersiowego uchodzą naczynia chłonne ze śródpiersia, pnie z lewej połowy głowy, szyi i lewej kończyny górnej. Drugim głównym naczyniem chłonnym, do którego odpływa chłonka z prawej górnej połowy ciała, jest krótki (1,5 cm) przewód chłonny prawy (ductus lymphaticus dexter). Uchodzi on do prawego kąta żylnego. Wpadają do niego naczynia chłonne z prawej połowy klatki piersiowej oraz z pni chłonnych szyjnego i podobojczykowego prawego.
Naczynia i węzły chłonne klatki piersiowej Naczynia chłonne płuc dzieli się na grupy: powierzchowną i głęboką. Naczynia grupy powierzchownej leżą tuż pod opłucną. Naczynia głębokie towarzyszą rozgałęzieniom naczyń płucnych oraz rozgałęzieniom oskrzeli. Z naczyń powierzchownych i głębokich chłonka odpływa do węzłów chłonnych położonych we wnękach płuc, a stąd do w ę z ł ó w śródpiersiowych przednich i tylnych, które leżą w okolicy rozwidlenia tchawicy. Do tych ostatnich uchodzi również limfa z przepony, osierdzia, serca, przełyku i częściowo ze ścian klatki piersiowej i sutka. Z naczyń odchodzących od węzłów śródpiersiowych powstaje pień oskrzelowo-śródpiersiowy, który po stronie prawej wpada do przewodu limfatycznego prawego, a po lewej - do przewodu piersiowego. '363
Naczynia i węzły chłonne głowy i szyi Rozróżnia się tu również naczynia powierzchowne i głębokie. Naczynia chłonne okolicy potylicznej uchodzą do w ę z ł ó w potylicznych, okolicy skroniowej i ciemieniowej - do w ę z ł ó w zausznych. Naczynia chłonne twarzy uchodzą do węzłów chłonnych podżuchwowych. Powierzchowne naczynia chłonne kończą się przeważnie w węzłach chłonnych szyjnych, leżących w okolicy żyły szyjnej zewnętrznej i wzdłuż tylnej krawędzi mięśnia mostkowo-sutkowo-obojczykowego. Rozróżnia się wśród nich węzły szyjne powierzchowne i głębokie. Inne naczynia wchodzą do w ę z ł ó w nadobojczykowych. Z głębszych warstw twarzy i z narządów szyi chłonka odpływa do węzłów leżących po obydwu stronach przełyku i krtani. Mózgowie nie ma haczyń chłonnych, istnieją tu tylko połączenia przestrzeni podpajęczynówkowej z ogólnym korytem chłonki przez osłonki nerwów mózgowych. Z naczyń chłonnych głowy i szyi tworzy się pień szyjny (truncus jugularis), który po stronie lewej uchodzi do przewodu piersiowego, a po prawej do przewodu chłonnego prawego.
Naczynia i węzły chłonne kończyny górnej Naczynia chłonne kończyny górnej dzielimy na powierzchowne i głębokie. Pierwsze z nich leżą w tkance podskórnej i większe ich pnie przebiegają równolegle do żył. Naczynia głębokie zbierają chłonkę z mięśni, kości i powięzi. Naczynia chłonne kończyny górnej uchodzą do węzłów chłonnych łokciowych i do kilkunastu węzłów pachowych. Te ostatnie łączą się również z naczyniami chłonnymi klatki piersiowej i sutka. Węzły pachowe mają połączenia z węzłami chłonnymi nadobojczykowymi. Chłonka z opisywanych okolic uchodzi za pośrednictwem pnia podobojczykowego (truncus subclavius) do głównych naczyń chłonnych: po stronie prawej do przewodu chłonnego prawego, po stronie lewej do przewodu piersiowego.
Śledziona Śledziona (lien) jest narządem barwy ciemnoczerwonej, spoistości miękkiej, o wymiarach średnio 4 x 7 x 12 cm i ciężarze ok. 150 g. Należy jednak zaznaczyć, że wymiary i ciężar śledziony są bardzo zmienne, zależnie od stanu i czynności organizmu. Na kształt śledziony wpływają sąsiadujące z nią narządy. Tak więc wyróżniamy na niej gładką i wypukłą powierzchnię przeponową oraz położone od '364
Ryc. 171. Śledziona od strony wnęki.
strony jamy brzusznej powierzchnie: żołądkową, nerkową i okrężniczą. Między powierzchniami nerkową i żołądkową znajduje się wnęka śledziony. Przez wnękę wnika do narządu tętnica śledzionowa (arteria lienalis) oraz wychodzi kilka gałęzi żylnych, łączących się w żyłę śledzionową (vena lienalis). Powierzchnię przeponową od pozostałych oddzielają brzegi: górny, na którym można dostrzec kilka wcięć, i dolny. Brzeg górny przechodzi w doi ny na końcach śledziony przednim i tylnym. Śledziona leży w podżebrzu lewym, schowana pod łukiem żebrowym, między IX a XI żebrem. Oś długa śledziony,łącząca oba końce, jest równoległa do X żebra. Śledziona przylega do przepony, żołądka, nerki lewej, okrężnicy i ogona trzustki.
Budowa śledziony Narząd otoczony jest torebką łącznotkankową. Wysyła ona przegrody do miąższu śledziony, dzieląc ją na nieregularne zraziki. Zarówno w torebce, jak i w przegrodach łącznotkankowych spotykamy włókna mięśniowe gładkie, umożliwiające kurczenie się śledziony. W miąższu śledziony wyróżniamy grudki śledzionowe, które są ośrodkami rozmnażania limfocytów. Przez każdą grudkę przechodzi tętniczka, która następnie rozpada się na drobne naczynia, zwane pędzelkami. Otwierają się one do rozszerzonych naczyń włosowatych żylnych, zwanych zatokami. Ściany zatok zbudowane są z komórek, o wydłużonych jądrach, w kształcie pręcików. Zatoki śledzionowe łączą się w żyły, które zbierają się w żyłę śledzionową. '365
Czynność śledziony Wspomniano wyżej, że w obrębie grudek śledzionowych odbywa się rozmnażanie limfocytów. Młodsze ich postacie leżą bliżej środka grudki. W miazdze czerwonej znajdują się również krwinki czerwone w różnych stadiach rozpadu. Tak więc w śledzionie z jednej strony przebiega rozpadanie się starych krwinek, a z drugiej powstawanie nowych elementów krwi. Przy małym zapotrzebowaniu część krwi zostaje wyłączona z krwiobiegu i magazynowana w różnych narządach. Śledziona magazynuje krew, a w chwilach dużego zapotrzebowania kurczy się, wyrzucając ją do naczyń. Daje się to czasami odczuć jako kłujący ból w podżebrzu lewym. Śledziona powiększa się nieco podczas trawienia. W chorobach zakaźnych, np. w zimnicy czy durze brzusznym, śledziona powiększa się znacznie i wystaje wtedy spod łuku żebrowego. Mimo spełniania tak ważnej roli w ustroju człowiek może żyć po operacyjnym usunięciu śledziony, gdyż czynności jej mogą być zastąpione przez inne narządy.
FIZJOLOGIA UKŁADU KRĄŻENIA Istota obiegu krwi zajmowała umysły ludzi już w czasach starożytnych, lecz dopiero w r. 1628 Wiliam Harvey ogłosił słynne dzieło pt. „Badania anatomiczne o ruchu serca i krwi u zwierząt". Dowiódł on, że serce jest rodzajem pompy, która tłoczy krew do tętnic. Po przejściu przez tkanki krew dostaje się do naczyń żylnych i powraca nimi do serca. Harvey odkrył istnienie krążenia wielkiego i małego, a swoje tezy poparł licznymi badaniami na zwierzętach. Uzupełnieniem tego wielkiego odkrycia była praca Malpighiego, który wykazał istnienie naczyń włosowatych, przez co zamknął układ krążenia w jedną całość.
Wiadomości ogólne Układ krążenia składa się z dwóch pomp i układu rur - naczyń. Pompami są prawa i lewa część serca, rurami - naczynia płucne i naczynia krążenia wielkiego. Krążenie wielkie rozpoczyna się w komorze lewej i prowadząc przez tętnice, naczynia włosowate i żyły ustroju, kończy się w przedsionku prawym. W krążeniu małym (płucnym) krew z komory prawej zdąża do płuc i po przejściu przez system naczyń włosowatych powraca do przedsionka lewego. Krew płynąc w naczyniach krążenia wielkiego zaopatruje tkanki w tlen i substancje odżywcze, pobiera zaś z nich produkty przemiany materii, przede wszystkim dwutlenek węgla. W obrębie krążenia małego następuje wydalanie C0 2 do pęcherzyków płucnych i pobieranie tlenu (ryc. 172).
'366
Zespół naczyń płucnych
mSSM
Prawa połowa serca Zespół naczyń krążenia wielkiego
Ryc. 172. Schemat krążenia krwi. Składa się nań układ dwóch pomp i dwóch układów naczyń.
Czynność serca Ruch krwi w naczyniach odbywa się jednokierunkowo, tzn. w tętnicach ku obwodowi, w żyłach zaś w kierunku serca. Jednokierunkowość biegu krwi jest uwarunkowana obecnością zastawek, które mogą się otwierać tylko w jedną stronę. Na czynność serca składają się trzy fazy: 1) skurcz (systole), 2) rozkurcz (diastole), 3) pauza, następująca po rozkurczu. W czasie pauzy krew z żył wpływa do przedsionków, a przez otwarte ujścia przedsionkowo-komorowe również do komór. W fazie skurczu pierwsze kurczą się przedsionki. Wyrzucają one do komór krew, która unosi zastawki i zbliża do siebie ich brzegi. Jednocześnie skurcz mięśni leżących dookoła żył wpadających do prawego przedsionka utrudnia cofanie się krwi. Bezpośrednio po skurczu przedsionków rozpoczyna się skurcz komór, składający się z dwóch faz: a) fazy napinania się mięśnia sercowego i b) fazy wyrzucania krwi do tętnic. W fazie napinania się mięsień sercowy naciska z dużą siłą krew znajdującą się w komorach, zastawki - dwudzielna i trójdzielna - zostają szczelnie domknięte, a skurczone mięśnie brodawkowe przytrzymują strunami ścięgnistymi płatki zastawek, nie pozwalając na ich wynicowanie. Napinanie mięśnia sercowego trwa do chwili, aż ciśnienie krwi w komorach przewyższy '367
ciśnienie krwi w tętnicach głównej i płucnej. Wtedy zastawki aorty i pnia płucnego otwierają się i krew zostaje wypchnięta do tętnic. Wyrzucanie krwi trwa tak długo, aż ciśnienie w komorach spadnie poniżej ciśnienia w tętnicach, wtedy zamykają się zastawki aorty i pnia płucnego. W dalszym ciągu rozpoczyna się faza rozkurczu komór, po czym mięsień sercowy przez pewien okres znajduje się w stanie spoczynku. Należy nadmienić, że rozkurcz przedsionków powstaje w fazie napinania się komór. Czasy) poszczególnych faz czynnościowych serca przedstawiają się w przybliżeniu następująco: Jedna ewolucja serca - 0,8 s
skurcz przedsionków - 0,12 s skurcz komór - 0,34 s pauza - 0,34 s
napinanie m. sercowego - 0,06 s — wyrzut krwi - 0,28 s
Uderzenie koniuszkowe i tony serca Uderzenie koniuszkowe jest to zgodne z akcją serca uwypuklanie się ściany klatki piersiowej w V lewym międzyżebrzu ok. 1,5 cm w prawo od linii środkowo-obojczykowej lewej. Powstaje ono wskutek skurczu mięśnia sercowego i obrotu serca wokół osi pionowej od strony lewej ku prawej. Dużą rolę odgrywa tu fakt, że serce w chwili tłoczenia krwi do tętnic, zgodnie z prawem reakcji, zostaje zepchnięte ku dołowi. Tony serca są to dźwięki powstające przy jego prawidłowej czynności. Można je wysłuchać słuchawką (stetoskopem) albo za pomocą przyrządu zaopatrzonego w membranę (fonendoskopem). Wyróżniamy dwa tony serca: skurczowy (systoliczny) i rozkurczowy (diastoliczny). Ton skurczowy, zwany tonem pierwszym, jest dłuższy od rozkurczowego, bardziej głuchy i niski. Składają się na niego trzy czynniki: 1) drgania zastawek w x chwili ich zamknięcia i drgania strun ścięgnistych, 2) drgania całego mięśnia sercowego, 3) drgania aorty i pnia płucnego spowodowane wyrzuconą przez komory krwią. Z wymienionych powyżej czynników drgania zastawek są najsilniejszym źródłem tonu skurczowego. Zaburzenia w ich budowie, a co za tym idzie i czynności, prowadzą do powstawania nieprawidłowych dźwięków, zwanych szmerami. Ton rozkurczowy jest krótszy i wyższy od skurczowego i powstaje wskutek drgań płatków półksiężycowatych oraz ścian tętnicy głównej i pnia płucnego. Zarówno ton skurczowy, jak i rozkurczowy powstają w różnych miejscach serca i można je na tej podstawie różnicować. Wraz z prądem krwi tony serca są przewodzone do określonych miejsc klatki piersiowej. Tak więc zastawkę dwudzielną osłuchujemy w miejscu uderzenia koniuszkowego. Zastawkę trójdzielną osłuchujemy w miejscu przyczepu V chrząstki żebrowej prawej do mostka. Zastawkę aorty osłuchujemy przy mostku w II międzyżebrzu prawym, zastawkę pnia płucnego - w II międzyżebrzu lewym. '368
o -204
-70
i Skurcz komo:
Rozkurcz komory Skurcz przedsionka
Skurcz izometryczny
Ryc. 173. Z m i a n y p o j e m n o ś c i w s k u r c z u i r o z k u r c z u .
W niektórych chorobach poza tonami wysłuchiwać można nieprawidłowe dźwięki, zwane szmerami. Szmery powstają wskutek wadliwej budowy połączeń między przedsionkami a komorami oraz między komorami a tętnicami, jak również wskutek nieprawidłowego połączenia między jamami serca. Nieprawidłowości budowy połączeń mcgą polegać na nieaomykainosci zastawek lub n3 zwężeniu otworów serca. Zarówno tony, jak i szmery powstające w sercu możemy zapisać w postaci wykresu za pomocą przyrządu zwanego fonokardi jgrafem.
Praca serca Pracujące serce przetłacza pewną objętość znajdującej się w ustroju krwi. Dzieląc objętość przez czas, w którym jest ona przepompowywana, otrzymujemy objętość minutową krwi (zwaną inaczej rzutem m i n u t o w y m serca} płynącej w aorcie i naczyniach płucnych. Tłoczona krew jest rozdzielana do odpowiednich narządów w zależności od ich aktualnego zapotrzebowania. Praca serca jest proporcjonalna do objętości przepompowanej krwi. Objętość krwi wypchniętej do tętnic w czasie jednego skurczu nosi nazwę objętości wyrzutowej i równa się w przybliżeniu 70 ml. Aby obliczyć rzut minutowy serca, mnożymy objętość wyrzutową przez liczbę skurczów serca w ciągu jednej minuty. Rzut minutowy = objętość wyrzutowa x częstość skurczów na minutę W spoczynku objętość minutowa wynosi około 5 I, a przy ciężkiej pracy fizycznej może dojść nawet do 25 I (!). Objętość minutowa uwarunkowana jest wielkością podstawowej przemiany materii. Przyjmuje się, że w normalnych warunkach życia potrzeba 62 ml krwi na kilogram ciężaru w ciągu 1 min. Znając więc ciężar ciała, można w przybliżeniu obliczyć rzut minutowy serca. 24 - Anatomia i fizjologia
369
Jak już wspomniano, objętość minutowa zmienia się w zależności od aktualnych potrzeb ustroju. Praca fizyczna może wpływać na podniesienie rzutu serca do 25 I, przy czym objętość skurczowa może osiągnąć wartość 120-140 ml, a nawet więcej. Podniesienie temperatury otoczenia powyżej 30° znacznie zwiększa objętość minutową, także trawienie może ją zwiększyć nawet do 40%.
B
c
Ryc. 174. Schematyczny przebieg krzywych: A - krzywa fonograficzna, B-krzywa ciśnienia krwi w komorach, C - krzywa elektrokardiograficzna. Faza skurczu komory oznaczona kolorem różowym.
W czasie jednej ewolucji serce wykonuje pracę równą 0,15 kGm, w ciągu doby ok. 15 500kGm (!). Krew zachowuje się tak, jak każda inna ciecz, która płynie od miejsca o wyższym ciśnieniu do miejsca o ciśnieniu niższym. Ciśnienie w lewej komorze waha się od 80 mm Hg (ciśnienie rozkurczowe) do 120 mm Hg (ciśnienie skurczowe). Średnie ciśnienie w komorze lewej wynosi ok. 100 mm Hg. Najniższe ciśnienie panuje w dużych żyłach i prawym przedsionku i jest równe w przybliżeniu ciśnieniu atmosferycznemu. Różnica ciśnień między komorą a przedsionkiem jest czynnikiem, dzięki któremu krew przepływa przez naczynia krwiobiegu wielkiego. Naczynia krwionośne stawiają opór przepływowi krwi. Nosi on nazwę oporu o b w o d o w e g o i jest odwrotnie proporcjonalny do czwartej potęgi średnicy naczynia. W krążeniu wielkim największe opory istnieją na poziomie tętniczek (arterioli). Naczynia włosowate stawiają mniejszy opór, ponieważ każdej '370
tętniczce odpowiada większa liczba równolegle biegnących kapilarów, a suma ich przekroju jest większa od przekroju odpowiedniej tętniczki. Z tego co powiedzieliśmy wynika, iż: 1. Przepływ krwi przez naczynia krążenia wielkiego, a zatem i praca serca, są wprost proporcjonalne do różnicy ciśnień i odwrotnie proporcjonalne do oporu obwodowego. 2. Przepływ przez naczynia jest równy przepływowi sercowemu (objętości krwi przepływającej przez serce). Zależności te można ująć następująco: _
,
Przepływ sercowy =
ciśnienie t ę t n i c z e - ciśnienie żylne 0pór obwodowy
a ponieważ ciśnienie żylne wynosi w przybliżeniu 0 to: _
,
ciśnienie tętnicze
Przepływ sercowy = - Q p ó r o b w o d o w y Zatem:
Ciśnienie tętnicze krwi = przepływ sercowy x opór obwodowy
•
|
A
Hr IB
C
v
A
^ i—i
s i
tw
i
i
i
Skurcz
i
J
s
i—i
i
i
Rozkurcz
_i
i
Ryc. 175. Ciśnienie krwi w: aorcie (A), komorach (B), przedsionkach (C). W dolnej części rysunku synchronizowany wykres elektrokardiograficzny.
'371
Objętość minutowa Na wstępie przypomnijmy sobie zależności: Objętość minutowa = objętość wyrzutowa x częstość skurczów na minutę Badania wykazały trzy dalsze zależności, a mianowicie: 1) objętość minutowa jest zależna od oporu obwodowego, 2) objętość minutowa jest równa powrotowi żylnemu (serce przetłacza krew ze strony żylnej na stronę tętniczą), 3) zmiany częstości skurczów nie muszą odbijać się na wielkości pojemności minutowej. Objętość m i n u t o w a a opór o b w o d o w y Z przedstawionego wyżej wzoru wynika, iż serce pracuje jak pompa o stałym przepływie, ale nie o stałym ciśnieniu. W warunkach prawidłowych serce jest w stanie zapewnić stały przepływ również przv zwiększonym oporze obwodowym. Wzrost oporu powoduje oczywiście większą pracę serca. Fakt, że opór obwodowy nie wpływa na objętość minutową, tłumaczy, dlaczego skurcz naczyń powoduje podwyższenie ciśnienia krwi, a rozkurcz - spadek. Objętość minutowa a powrót żylny W warunkach prawidłowych serce jest w stanie przepompować wszystką krew dostarczaną przez żyły do serca. Większy dopływ krwi jest równoważony większym rozszerzaniem się komór w rozkurczu. Podczas następnego skurczu mięsień sercowy kurczy się mocniej. Dzięki temu zwiększa się objętość wyrzutowa (skurczowa). Podczas osłabienia serca krew zalega w żyłach, które rozszerzają się. Stan ten nazywa się niewydolnością serca. Objętość wyrzutową warunkuje ilość krwi znajdującej się w przedsionkach na początku skurczu. Wszystkie żyły zawierają 3 / 5 krwi krążącej. Pojemność układu żylnego jest najważniejszym czynnikiem warunkującym wydolność krążenia. Zmiana napięcia mięśni zaopatrujących żyły powoduje zmiany ich pojemności, a jednocześnie nie wpływa na opory krążenia. Niezbędna ilość krwi dopływająca do przedsionków zależy od wspomnianych dwóch czynników - pojemności żył i ich zdolności do zmiany objętości. Przy objętości 5 I krwi i wypełnieniu dwu przedsionków po 70 ml krwi każdy, objętość reszty krwi wynosi 5 0 0 0 - 1 4 0 = 4860 ml. Nie wyrównany wzrost pojemności naczyń o 1,5% zmniejsza objętość skurczową do połowy. W takim przypadku objętość krwi w naczyniach wyniesie 4930 ml, a do każdego z przedsionków napłynie zaledwie 35 ml krwi. Przy nie wyrównanym wzroście pojemności naczyń o 3% serce nie może pracować z powodu braku krwi. Z powyższego wynika, jak ważne znaczenie dla prawidłowej czynności serca ma napięcie ścian żylnych i ich zdolność do kurczenia się. W czasie skurczu żył więcej krwi dopływa do przedsionków i wzrasta rzut sercowy. '372
Przy zmianie objętości krwi, np. podczas krwotoku, występuje wyrównawczy skurcz żył, a jeżeli to nie wystarcza, zmniejsza się również objętość skurczowa i rzut minutowy serca. Zmniejszenie rzutu odbija się na niedokrwieniu tkanek, co obserwujemy we wstrząsie. Pod wpływem ciążenia krew dąży do gromadzenia się w żyłach poniżej poziomu serca i rozszerza je. Spadek napięcia ścian żył powoduje obniżenie rzutu sercowego. Dzieje się to miedzy innymi podczas działania przyspieszenia o wartości kilku g. Ponieważ krew gromadzi się w częściach ciała leżących w kierunku przeciwnym do kierunku przyspieszenia, kosmonauci w czasie startu rakiety kosmicznej muszą być ubrani w obcisłe kombinezony i zajmować poziomą pozycję ciała prostopadłą do kierunku przyspieszenia. Piloci szybkich samolotów odrzutowych, którzy zajmują pozycję siedzącą, ubrani są w kombinezony uciskające dolną część ciała w celu zapobieżenia gromadzeniu się krwi w kończynach dolnych podczas wirażowania. Jednym z ważnych czynników wpływających na krążenie krwi w żyłach jest tzw. pompa mięśniowa. Skurcz mięśni przetłacza krew z odcinków między zastawkami w kierunku serca. Uszkodzenie lub niedomykalność zastawek (np. w żylakach kończyn) upośledza przepływ krwi żylnej. Krążeniu żylnemu sprzyja również pompa oddechowa. Wiemy, że ciśnienie w dużych żyłach w klatce piersiowej wynosi 0 mm Hg, natomiast ciśnienie w klatce piersiowej jest ujemne i wynosi od -2 mm Hg w fazie wydechu do -8 mm Hg we wdechu. Przedsionki serca znajdują się pod wpływem tego
I IBiilwsills
iJlfcssililt i
l
i
IkhI
A
B
C
Ryc. 176. Mechanizm krążenia krwi w żyłach. A - żyła z zamkniętymi zastawkami, B - u c i ś n i ę c i e żyły powoduje przetłoczenie krwi do odcinka żyły położonego ponad zastawką, C- zwolnienie ucisku powoduje zassanie krwi z obwodowego odcinka naczynia.
'373
ujemnego ciśnienia i krew jest przez nie zasysana. W czasie ciężkiej pracy różnica ciśnień może się wahać od -2 do - 3 0 mm Hg, co znakomicie polepsza warunki powrotu krwi do serca. Objętość minutowa a częstość skurczów Serce pobudzają do skurczów bodźce powstające w samym sercu, a mianowicie w węźle zatokowym. Regulacją częstości skurczów serca zawiaduje ośrodek sercowy, leżący w rdzeniu przedłużonym i składający się z dwu ośrodków: hamującego i pobudzającego serce. Ośrodek hamujący działa poprzez nerw błędny, ośrodek pobudzający działa przez nerwy współczulne. Wwarunkach spokoju istnieje przewaga napięcia nerwu błędnego,a częstość skurczów serca wynosi 70/min. Zwiększone napięcie układu przywspółczulnego (nerw błędny) działa hamująco na serce. Im silniejsza jest czynność nerwu błędnego, t y m wolniej bije serce. Niewielkie zmiany częstotliwości regulują wahania napięcia nerwu błędnego. Całkowite wyłączenie czynności nerwu błędnego prowadzi do zwiększenia częstości skurczów do 150/min. W silnych stanach emocjonalnych i podczas intensywnej pracy fizycznej częstość skurczów może się zwiększyć nawet do 210/min. Tak duże przyspieszenie zachodzi za pośrednictwem zwiększonego napięcia układu wspołczulnego, który powoduje uwolnienie się noradrenaliny i adrenaliny. Ośrodek sercowy, podobnie jak ośrodek naczynioruchowy, znajduje się pod wpływem czynników humoralnych i nerwowych. Na przykład baroreceptory pobudzają część hamującą ośrodka sercowego, stąd też wzmożenie napięcia nerwu błędnego powoduje spadek ciśnienia krwi i częstości skurczów. Wzrost ciśnienia jest powodem zwolnienia czynności serca i zmniejszenia objętości minutowej. Zmniejszenie tej objętości działa w tym samym kierunku co rozszerzenie naczyń zmniejszające ciśnienie krwi. Odwrotnie-spadek ciśnienia krwi zmniejsza napięcie nerwu błędnego i przyspiesza czynność serca. Zwiększa się wówczas objętość minutowa. Na ośrodek sercowy wpływają również wyższe ośrodki nerwowe. Pobudzenie emocjonalne przyspiesza czynność serca dzięki zmniejszeniu napięcia nerwu błędnego, natomiast bardzo silne pobudzenie może doprowadzić do gwałtownego spadku ciśnienia wskutek pobudzenia tego nerwu. U dzieci i młodzieży głębokie oddychanie powoduje wahania napięcia nerwu błędnego i wpływa na przyspieszanie i zwalnianie czynności serca (tzw. arytmia oddechowa). Czynność serca przyspiesza się podczas wdechu i zwalnia podczas wydechu. Podrażnienie nerwów czuciowych w różny sposób wpływa na częstość skurczów. Silne pobudzanie bólowe przyspiesza serce, lecz niektóre urazy, np. uderzenie w brzuch, mogą zatrzymać serce. Niedobór tlenu oraz wzrost dwutlenku węgla wpływa bezpośrednio na ośrodek sercowy podobnie jak podwyższona temperatura ciała. Również hormon tarczycy - tyroksyna, pobudza ośrodek sercowy. '374
Objętość minutowa a objętość wyrzutowa (rzut serca) Z zależności: objętość minutowa = objętość wyrzutowa x częstość skurczów wynika, że serce może pompować tę samą ilość krwi dzięki większej liczbie skurczów lub większej objętości wyrzutowej. Osierdzie ogranicza pojemność serca. W czasie ciężkiej pracy objętość minutowa osiąga 40 I krwi/min przy objętości wyrzutowej 200 ml i częstości skurczów serca do 200/min.
Zjawiska elektryczne w sercu Pracy mięśnia sercowego towarzyszą zjawiska elektryczne, które mogą być badane za pomocą przyrządu, zwanego elektrokardiografem. W spoczynku mięsień sercowy ma ładunek dodatni (+), w okresie pobudzenia - ładunek ujemny (-). Wiemy, że w sercu pobudzenie biegnie od przedsionków do komór, najpierw więc pobudzone są przedsionki, a następnie komory. W ten sposób w różnych częściach serca powstaje różnica potencjałów. Zmiany wielkości napięć elektrycznych można wykryć w obwodowych częściach ciała i odprowadzić do elektrokardiografu, gdzie po odpowiednim wzmocnieniu otrzymuje się ich wykres (elektrokardiogram).
Węzeł zatokowy Węzeł przedsionkowo-komorowy Mięsień komór
R
V
T
Q—S— Ryc. 177. Schemat ilustrujący miejsca powstania potencjałów elektrycznych odzwierciedlonych krzywą ekg (wg Greena).
'375
W celu uzyskania dokładnych danych elektrokardiograficznych stosujemy kilka odprowadzeń prądu z różnych okolic ciała. Najważniejsze są odprowadzenia następujące: 1) odprowadzenie I: ręka prawa - ręka lewa, 2) odprowadzenie II: ręka prawa - noga lewa, 3) odprowadzenie III: ręka lewa - noga prawa. Poza odprowadzeniami I, II i III, w których elektrokardiograf zapisuje różnice potencjałów pomiędzy dwoma odległymi punktami, stosuje się z reguły odprowadzenia przedsercowe, oznaczone symbolem C. Przy takim badaniu jedna z elektrod znajduje się w bezpośrednim sąsiedztwie serca, druga może znajdować się na: 1) kończynie górnej prawej - odprowadzenie CR, 2) kończynie górnej lewej - odprowadzenie CL, 3) w okolicy kręgosłupa - odprowadzenie CB. Często przewody biegnące z kończyn górnych i kończyny dolnej lewej łączymy ze sobą, otrzymując tzw. elektrodę centralną. Odprowadzenie przedsercowe, którego drugim biegunem jest elektroda centralna, oznaczono symbolem CV. Elektrodę przedsercową umieszcza się w jednym z sześciu określonych punktów, położonych w okolicy mostka i lewej połowy klatki piersiowej. Punkty te ponumerowano kolejnymi cyframi od 1 do 6. Tak więc pełny symbol odpowiedniego odprowadzenia przedsercowego brzmi np. CR1, co oznacza, że elektroda przedsercową znajdowała się w punkcie 1, a elektroda druga umieszczona była w prawej kończynie górnej.
Bez względu na rodzaj odprowadzenia wyróżniamy następujące charakterystyczne odcinki krzywej elektrokardiograficznej : 1) załamek P - powstaje przy skurczu przedsionków, 2) krzywa QRS - świadczy o charakterze początku skurczów komór, 3) załamek T - powstaje w czasie rozkurczu komór.
'376
Poszczególne elementy krzywej elektrokardiograficznej przedstawiono na ryc. 178. Oglądając krzywą, należy zwrócić uwagę na rodzaj odprowadzenia, jej kształt, wysokość załamków i czas ich trwania. Wszystkie dane można odczytać z elektrokardiogramu, na którym obok samej krzywej istnieje zespół linii pionowych wyznaczonych co 1/25 s, i poziomych, gdzie podziałka wielkości 1 cm oznacza napięcie 1 mV. Prawidłowe wartości są następujące: Odstępy: P-Q QRS ST TP Załamek: P QRS T
Czas trwania w s: 0,18 0,08 0,24 0,32 Napięcie w mV: 0,25 2,0 0,5
Podane liczby mają charakter jedynie orientacyjny i wahają się w szerokich granicach, w zależności od położenia serca, rodzaju odprowadzenia i wielu innych czynników. Należy zaznaczyć, że elektrokardiograficzna metoda badania czynności serca jest bardzo ścisła i szeroko stosowana w diagnostyce.
Automatyzm serca Doświadczalnie stwierdzono, że serce po wyizolowaniu go z ustroju kurczy się nadal, jeżeli ma zapewnione odpowiednie warunki. Najłatwiej przekonać się o tym na przykładzie serca żaby, które umieszczone w roztworze fizjologicznym soli kurczy się przez wiele minut, a nawet godzin. Serce pracuje dłużej i sprawniej, jeżeli przepływa przez nie płyn Ringera-Locka (u zwierząt stałocieplnych jest to warunek konieczny). Z doświadczeń wynika, że mięsień sercowy jest źródłem własnych skurczów. Bodziec wywołujący skurcze powstaje w komórkach mięśniowych przewodzących serca, tzw. włóknach Purkinjego. Rozmieszczone są one w sposób bardzo charakterystyczny, a ich zespół nosi nazwę układu przewodzącego serca. Głównym miejscem, w którym powstają rytmiczne bodźce, jest węzeł zatokowo-przedsionkowy (Keitha-Flacka), leżący w okolicy ujścia żyły głównej górnej do przedsionka prawego. Pobudzenie rozchodzi się z niego najpierw na przedsionki, a następnie na komory. W przypadku uszkodzenia węzła zatokowo-przedsionkowego rolę czynnika wyzwalającego bodźce przejmuje niższe piętro układu przewodzącego, tj. węzeł przedsionkowo-komorowy (Aschoffa-Tavary), leżący w przegrodzie przedsionkowo-komorowej prawej. Od niego do mięśni beleczkowatych i brodawkowatych biegnie pęczek przedsionkowo-komorowy (Hisa). Przerwanie połączenia między przedsionkami a komorami prowadzi do tzw. bloku przedsionkowo-komorowego, który objawia się tym, że przedsionki kurczą się niezależnie od komór. Jest to jeden z rodzajów tzw. bloku serca. '377
Czynność naczyń krwionośnych Krążenie k r w i w tętnicach Dzięki elastycznym ścianom tętnic krew płynie w nich nie skokami, ale strumieniem ciągłym. Mechanizm tego zjawiska jest następujący: podczas skurczu serce wyrzuca krew do tętnic pod dużym ciśnieniem, rozciągając ich ściany. Gdy serce przestanie tłoczyć krew, rozszerzona tętnica stopniowo się kurczy, przesuwając krew do dalszych odcinków, co z kolei wywołuje rozszerzenie oraz następowy skurcz tętnicy. W ten sposób zredukowane zostają gwałtowne skoki ciśnienia i w małych tętnicach krew płynie już strumieniem ciągłym. Podczas przechodzenia krwi od dużych tętnic przez tętniczki i naczynia włosowate do dużych żył średnie ciśnienie krwi u człowieka spada od ok. 120 do 0 mm Hg w stosunku do ciśnienia atmosferycznego. Jak już wspominaliśmy, miejscowe ciśnienie krwi w naczyniach zależy od oporów. Podczas przejścia krwi przez tętniczki ciśnienie spada do 32 mm Hg, a w naczyniach włosowatych ciśnienie nadai obniża się, osiągając wartość 12 mm Hg. Różnica ciśnień między naczyniami włosowatymi a żyłami jest wystarczająca do jednokierunkowego przepływu krwi do żył. Ciśnienie k r w i Z rozważań nad pracą serca wynika, że ciśnienie krwi w tętnicach jest zmienne i wzrasta w następstwie skurczu serca, maleje zaś przy jego rozkurczu. Ciśnienie krwi w tętnicach, występujące w czasie skurczu serca, nazywamy ciśnieniem skurczowym (systolicznym). Ciśnienie rozkurczowe (diastoliczne) powstaje w następstwie chwilowego braku dopływu krwi z serca (rozkurcz i pauza). Ciśnienie skurczowe mierzymy w milimetrach słupa rtęci (mm Hg).* Bezpośredni pomiar ciśnienia krwi stosuje się do badania ciśnień w przedsionkach i komorach serca podczas jego cewnikowania cewnikiem wprowadzonym najczęściej przez żyłę pośrodkową łokcia. Pomiary takie są niezbędne dla dokładnego rozpoznawania wad serca i nieprawidłowości krążenia. W sposobie bezkrwawym oznaczania ciśnienia krwi opieramy się na metodzie wynalezionej przez Korotkowa, a opracowanej przez Riva-Rocci. Przyrząd służący do bezkrwawego oznaczania ciśnienia krwi nosi nazwę sfigmomanometru. Składa się on ze szczelnego podwójnego mankietu gumowego, z którego światłem łączą się dwie rurki gumowe. Jedna z nich zespolona jest z manometrem rtęciowym lub sprężynowym, druga łączy się z pompą gumową. Mankiet zakładamy na ramię nieco powyżej przegubu łokciowego. W rzucie * Na podstawie zarządzenia Prezesa Polskiego Komitetu Normalizacji i Miar z d n . 5 I 1976 r. obowiązującą jednostką ciśnienia jest 1 paskal (Pa). 1 mm Hg = 0,1333 kPa.
'378
Ryc. 179. Rozkład ciśnień w naczyniach krążenia wielkiego. Kolorem szarym oznaczono zakresy ciśnień we włośniczkach.
'379
tętnicy ramiennej w zgięciu łokciowym przykładamy fonendoskop. Następnie pompujemy powietrze do mankietu, uciskając w t e n sposób tętnicę ramienną. W miejscu zwężenia krew przeciska się z trudem, wywołując rytmiczne szmery słyszalne przez fonendoskop. Przez dalsze pompowanie powietrza do mankietu całkowicie zamykamy światło tętnicy i wówczas szmery znikają. Teraz powoli wypuszczamy powietrze przez specjalny zawór i z chwilą pojawienia się pierwszych szmerów odczytujemy na manometrze ciśnienie skurczowe. Gdy wypuszczamy powietrze w dalszym ciągu, szmery przestają być słyszalne (zanikł ucisk tętnicy), a manometr wykazuje wtedy wielkość ciśnienia rozkurczowego. Na początku pompowania mankietu sfigmomanometru, kiedy dochodzi tylko do zamknięcia żył i krew nie może opuścić obwodowej części kończyny, dochodzi do wzrostu jej objętości, będącej miarą objętości dopływającej krwi. Zjawisko to stanowi podstawę pletyzmografii, metody pozwalającej mierzyć przepływ krwi przez kończynę.
mm Hg zamykając światło żył. Wzrost objętości krwi w kończynie ilustruje krzywa A-B (wg Greena).
'380
W p ł y w siły ciężkości na ciśnienie krwi Omawiane dotychczas ciśnienie krwi w naczyniach dotyczy pomiarów na poziomie serca. Poniżej serca w p ł y w siły ciężkości w naczyniach zwiększa ciśnienie krwi o: l 0 h
'
1
m
m m H g
'
gdzie: h - pionowa odległość serca od miejsca pomiarów, 1,055 - ciężar właściwy krwi, 13,6 — ciężar właściwy rtęci. Ciśnienie krwi powyżej serca zmniejsza się według podanej zasady.
lifilr Ryc. 182. W p ł y w siły ciężkości na żylne i tętnicze ciśnienie krwi. Na poziomie serca ciśnienie skurczowe krwi tętniczej wynosi około 120 mm Hg, krwi żyinej 0 mm Hg.
'381
W pozycji stojącej, przy ciśnieniu na poziomie serca 120 mm Hg,jestono na poziomie stóp większe o ok. 80 mm Hg i wynosi 200 mm Hg. Podobnie o 80 mm Hg wzrasta ciśnienie krwi żylnej. Ciśnienie na poziomie głowy jest niższe o ok. 30 m m Hg. Ciśnienie w żyłach powinno być również teoretycznie niższe i wynosić 30 mm Hg. W praktyce pozostaje ono równe ciśnieniu atmosferycznemu wskutek zapadnięcia ścian żył powierzchownych. Z tego też powodu w zatokach żylnych czaszki, mających sztywne ściany, ciśnienie krwi jest niższe od atmosferycznego. Ujemne ciśnienie w żyłach ujawnia się przy uszkodzeniu ich ściany. Dochodzi wówczas do groźnego dla życia zasysania powietrza (zator powietrzny). Tętno Tętnem nazywamy rytmiczne podnoszenie i zapadanie ścian tętnic. Wiemy, że krew wyrzucona z komory rozciąga ściany tętnic, które bezpośrednio potem kurczą się, co daję wrażenie tętna. Fala tętna rozchodzi się z szybkością ok. 9 m/s, lecz - trzeba to podkreślić - nie pokrywa się z falą prądu krwi, której szybkość wynosi jedynie 0,5 m/s. Tętno możemy wyczuć wszędzie tam, gdzie tętnice przebiegają powierzchownie. Najczęściej wyczuwamy je na tętnicy promieniowej, szyjnej wspólnej, skroniowej powierzchownej i tętnicy grzbietowej stopy. Badanie tętna przynosi informacje o liczbie skurczów serca, objętości skurczowej oraz do pewnego stopnia o ciśnieniu krwi. Określając charakter tętna, opisujemy jego częstość, miarowość, napięcie i szybkość. Tętno może być rejestrowane również graficznie, za pomocą przyrządu zwanego sfigmografem. Ruchy ściany tętnicy promieniowej zostają przeniesione za pomocą układu dźwigni na ramię, które na papierze wykreśla krzywą. Jest to krzywa sfigmograficzna. Krzywa składa sięz regularnych wzniesień. Na każdym z nich można wyróżnić ramię wstępujące (anakrotyczne) i zstępujące (katakrotyczne). Ramię wstępujące przebiega bardziej pionowo i odpowiada części czołowej fali. Na ramieniu zstępującym widoczne jest dodatkowe załamanie krzywej, które nosi nazwę fali dykrotycznej. Powstanie fali dykrotycznej tłumaczymy w następujący sposób: gdy ustanie skurcz komory, krew z tętnic usiłuje się cofnąć, natrafia jednak na zamknięte zastawki półksiężycowate, odbija się od nich, co prowadzi do ponownego rozszerzenia tętnicy i powstania małej fali dykrotycznej. Z kształtu krzywej sfigmograficznej można wyciągnąćwiele wniosków co do prawidłowej lub patologicznej czynności serca i naczyń. Zmiany mogą dotyczyć wysokości fali i kąta nachylenia jej ramion - wstępującego i zstępującego. Niekiedy można również zauważyć dodatkowe fale występujące zarówno przed, jak i po fali dykrotycznej. Szybkość prądu k r w i Szybkość prądu krwi możemy rozpatrywać jako prędkość objętościową albo prędkość liniową. Pierwsza z nich mówi o objętości krwi, która przepływa '382
przez określony przekrój naczynia w jednostce czasu. Jest ona jednakowa we wszystkich częściach układu krwionośnego. Szybkość liniowa mówi o drodze, którą przebywa krew w jednostce czasu. Szybkość ta jest większa w naczyniach dużych i w aorcie, gdzie wynosi 0,5 m/s, w tętnicach obwodowych (promieniowa, piszczelowa) zmniejsza się do ok. 0,2-0,25 m/s, a w naczyniach włosowatych wynosi tylko 0,5 mm/s. Czas, w którym krew przechodzi przez wielki i mały obieg, nazywamy czasem obiegu krwi. U człowieka trwa on około 20 s.
Krążenie k r w i w tkankach Dla utrzymania się przy życiu tkanki ustroju wymagają odpowiedniego przepływu krwi, której objętość zależy w dużej mierze od ciśnienia w naczyniach je zaopatrujących. Do tkanek szczególnie wrażliwych na zmniejszenie przepływu krwi należy tkanka nerwowa, a przede wszystkim mózg, który obumiera już po kilku minutach od ustania krążenia. Mózg człowieka stojącego lub siedzącego znajduje się wyżej niż serce, stąd ciśnienie w tętnicach mózgu człowieka znajdującego się w pozycji pionowej jest o 30-40 mm Hg niższe niż na poziomie serca. Różnica znika w pozycji leżącej. Zjawisko to wykorzystujemy w leczeniu utraty świadomości spowodowanej spadkiem ciśnienia krwi (omdlenie). Prawidłowe ciśnienie krwi jest niezbędne dla sprawnej pracy narządów. Na przykład spadek ciśnienia w nerkach powoduje zmniejszoną filtrację i zahamowuje lub zatrzymuje wydalanie moczu. Przypomnijmy sobie, iż: ciśnienie tętnicze = przepływ sercowy x opór obwodowy i rozpatrzmy rolę oporu obwodowego w przepływie krwi przez tkanki. Wspomnieliśmy już wyżej, iż opór ten jest największy w najmniejszych tętniczkach. Wiemy również, że opór wzrasta w miarę zmniejszania się światła naczyń, np: zmniejszanie światła naczynia z 1,2 do 0,8 mm powoduje czterokrotne zwiększenie oporu. Najważniejszym sposobem regulacji ciśnienia krwi w tkankach jest zatem regulacja szerokości tętniczek. Ściany tętniczek zawierają mięśnie gładkie, które są w stanie częściowego przykurczu pod wpływem działania nerwów współczułnych. Wzrost napięcia układu współczulnego powoduje zwężenie tętniczek i wzrost oporów krążenia. Rozszerzenie tętniczek prowadzi do zmniejszenia oporów i spadku ciśnienia krwi.
Regulacja przepływu tkankowego krwi Baroreceptory Stała wielkość ciśnienia krwi zależy od obecności odpowiednich czujników, tzw. baroreceptorów. Znajdują się one w ściankach niektórych tętnic i informują ośrodek naczynioruchowy o wielkości ciśnienia. Baroreceptory stwier'383
dzono w łuku aorty, tętnicach szyjnych wspólnych, w miejscu rozwidlenia się na tętnicę szyjną zewnętrzną i wewnętrzną (zatoka szyjna). Wzrost ciśnienia przyspiesza częstotliwość wysyłania impulsów z baroreceptorów do ośrodka naczyniowego i odwrotnie-spadek ciśnienia pociąga za sobą obniżenie liczby impulsów. Zwiększona aktywność baroreceptorów hamuje ośrodek naczynioruchowy, który przez włókna odśrodkowe zmniejsza napięcie mięśni gładkich tętnic. Tętnice rozszerzają się - ciśnienie krwi obniża. Odwrotnie - niska częstotliwość impulsów wysyłanych przez baroreceptory pobudza czynność ośrodka naczynioruchowego, który wysyła bodźce do mięśni gładkich tętnic zwiększając ich napięcie. Układ regulacyjny działa na zasadzie sprzężenia zwrotnego według schematu: Ciśnienie krwi •Ą— baroreceptory ośrodek naczynioruchowy
ł
mięśnie tętniczek
1
Wysoka sprawność regulacji ciśnienia krwi przez baroreceptory pow; iż nie zmienia się ono w pozycji leżącej ani stojącej. Niekiedy baroreceptory mogą utracić swą sprawność. Po długotrwałym przebywaniu w łóżku, np. z powodu ciężkiej choroby, zmiana położenia ciała na pionową może doprowadzić do omdlenia. Podobne warunki istnieją u astronautów przebywających przez dłuższy czas w warunkach nieważkości. Wspomnieliśmy już, że czynność ośrodka naczynioruchowego zależy nie tylko od impulsów napływających z baroreceptorów, ale również od szeregu innych czynników, z których najważniejsze to: 1) stężenie dwutlenku węgla we krwi, 2) wysycenie krwi tlenem (stężenie 02), 3) czynność ośrodka oddechowego, 4) działanie nerwów czuciowych, 5) czynność wyższych ośrodków układu nerwowego. Stężenie C02 Dla czynności ośrodka naczyniowego ważne znaczenie ma prawidłowy poziom dwutlenku węgla we krwi. Zwiększenie zawartości C02 pobudza czynność ośrodka i podnosi ciśnienie krwi, jego zmniejszenie hamuje funkcję ośrodka. Wahania ciśnienia krwi w zależności od stężenia C02 pozwalają utrzymać prawidłowy poziom bezwodnika węglowego we krwi. Podczas prowadzenia sztucznego oddychania należy pamiętać o możliwości nadmiernej wentylacji, w czasie której dochodzi do usunięcia dwutlenku węgla z krwi, co powoduje spadek ciśnienia. '384
Stężenie 0 2 Zmniejszenie wysycenia krwi tlenem pobudza czynność ośrodka naczynioruchowego bezpośrednio i za pośrednictwem specjalnych czujnikó3v, zwanych chemoreceptorami. Chemoreceptory, podobnie jak baroreceptory, leżą w ścianie aorty i tętnicach szyjnych. Informują one ośrodek naczynioruchowy 0 spadku stężenia tlenu we krwi. Nerwy czuciowe Podrażnienie nerwów czuciowych może również wpływać na czynność serca. Dotyczy to szczególnie nerwów przewodzących ból. Niewielkie podrażnienie nerwów pobudza ośrodek wazomotoryczny, co prowadzi do wzrostu ciśnienia. Silny ból zmniejsza aktywność ośrodka naczynioruchowego i obniża ciśnienie krwi. Ośrodkowy układ n e r w o w y Mózg oddziałuje na czynność ośrodka naczynioruchowego przede wszystkim w stanach silnego napięcia emocjonalnego, które pobudza ośrodek. Wzrost ciśnienia łączy się ze zwiększeniem rzutu sercowego. Jednocześnie dochodzi do zwiększenia napięcia układu współczulnego i wzmożonej czynności nadnerczy, wyrażającej się nasileniem wydzielania noradrenaliny i innych katecholamin. W niektórych rodzajach wstrząsu emocjonalnego w p ł y w wyższych ośrodków nerwowych może być odwrotny - rozszerzenie tętniczek 1 spadek ciśnienia krwi (omdlenie w przestrachu). Inne czynniki regulujące objętość łożyska naczyniowego Ośrodek naczynioruchowy wpływa, jak wiemy, na objętość krwi krążącej w naczyniach obwodowych poprzez regulację napięcia mięśni zwężających naczynia, przede wszystkim tętniczki. Intensywna czynność narządów wymaga zwiększonego przepływu krwi i ośrodek naczynioruchowy zmniejsza napięcie mięśni zwężających naczynia. Jego czynność wspomagają miejscowe mechanizmy regulacji. W celu utrzymania stałego ciśnienia w naczyniach obwodowych jednocześnie z rozszerzeniem naczyń w narządzie pracującym zwężają się naczynia w narządach pozostających w spoczynku. W ten sposób dochodzi do przemieszczenia krwi z narządów nie pracujących do narządów, w których istnieje większe zapotrzebowanie na krew. Produkty przemiany materii są czynnikami powodującymi rozszerzenie naczyń. Należą do nich przede wszystkim dwutlenek węgla i kwas mlekowy. Dwutlenek węgla wpływa na napięcie błony mięśniowej tętniczek bezpośrednio i za pośrednictwem ośrodka naczynioruchowego. Nagromadzenie produktów przemiany materii wywołane upośledzeniem krążenia wywołuje ból zwany ischemicznym. 25 - Anatomia i fizjologia
385
Wpływ nerwów rozszerzających naczynia Jak już wiemy, nerwy współczulne wpływają na zwężenie naczyń obwodowych, a rozszerzenie naczyń serca i mózgu. Rozszerzenie naczyń jest spowodowane zmniejszeniem napięcia tego układu. Wiemy także, że mięśnie naczyń krwionośnych są unerwione również przez nerwy przywspółczulne, powodujące rozszerzenie naczyń. Obecnie uważa się, że aktywne rozszerzanie naczyń odbywa się za pośrednictwem bradykininy, powstającej w wyniku pobudzenia układu przywspółczulnego. Układ naczyń jest również powiązany z układem nerwowym somatycznym. Na przykład drażnienie tylnych korzeni rdzenia kręgowego, przewodzących czucie od obwodu do ośrodkowego układu nerwowego, powoduje rozszerzenie naczyń skóry w polu zaopatrywanym przez te korzenie. Również bezpośrednie podrażnienie skóry może spowodować rozszerzenie naczyń. Niewielkie uciśnięcie skóry wąskim przedmiotem do granicy bólu powoduje wkrótce po uciśnięciu zblednięcie skóry, ograniczone do miejsca ucisku, które znika po kilku minutach. Jest to wywołane miejscowym skurczem naczyń włosowatych, które kurczą się, jeśli ciśnienie w nich spada do 50 mm Hg. Podniesienie ciśnienia do 90 mm Hg otwiera je ponownie. Przy większej sile ucisku i skaleczeniu skóry dochodzi do następujących zmian: 1. W miejscu ucisku powstaje czerwona iinia wskutek rozszerzenia naczyń włosowatych. 2. W ciągu następnych 30 s pojawia się zaczerwienienie w otoczeniu rany, obejmujące obszar zaopatrywany przez tętniczkę. Jednocześnie podnosi się miejscowo temperatura, co wywołane jest rozszerzeniem naczyń. Objawy mają charakter odruchowy i występują w przypadku nie uszkodzonego układu nerwowego. 3. Wokół miejsca zranienia powstają pęcherzyki wypełnione płynem. Opisane zmiany powstają w wyniku różnych niewielkich uszkodzeń skóry, jak ukłucie, zadrapanie, oparzenie termiczne, elektryczne, chemiczne, ukąszenie owadów i inne. Mechanizm zmian poiega na wpływie na naczynia histaminy uwolnionej podczas uszkodzenia skóry. Wydzielanie dużej ilości histaminy obserwujemy w reakcjach zachodzących między obcym białkiem i ciałami obronnymi ustroju. Wstrzyknięcie po raz pierwszy obcego białka do ustroju nie powoduje groźnych objawów, ale jest przyczyną rozwoju nadwrażliwości. Ujawnia się ona przy następnych wstrzyknięciach takiego samego białka. Rozwijają się wówczas objawy zatrucia histaminą, uwolnioną podczas reakcji łączenia się obcego białka (antygenu) z przeciwciałami, które wytworzyły się po pierwszym wstrzyknięciu obcego białka. Uogólnione rozszerzanie naczyń włosowatych w czasie wstrząsu anafilaktycznego prowadzi do gwałtownego spadku ciśnienia i może być przyczyną śmierci.
Skóra jest jedynym narządem, w którym ukrwienia nie regulują zachodzące w niej procesy przemiany materii, ale ośrodek regulacji temperatury leżący w pniu mózgowym. Podwyższona temperatura zewnętrzna powoduje rozszerzenie naczyń skóry. Zanurzenie nogi w gorącej wodzie o temperaturze 40-45°C powoduje rozszerzenie naczyń nie tylko skóry kończyny zanurzonej, ale również przeciwnej, a niekiedy również skóry rąk i twarzy. Jestto wynikiem podniesienia się temperatury krwi. Efekt działania niskiej temperatury jest złożony. Przy temperaturze 10-20° rtastępuje skurcz naczyń. Skurcz tętniczek powoduje zablokowanie krwinek '386
w naczyniach włosowatych. Utrata przez te krwinki tlenu powoduje miejscową sinicę. Przy jednoczesnym skurczu naczyń włosowatych skóra blednie. Zanurzenie rąk do wody o temperaturze 5-10° powoduje zaczerwienienie skóry, wywołane odruchem obronnym przed zimnem. Czerwone zabarwienie skóry wynika również z wolniejszego w niższych temperaturach odłączania tlenu od hemoglobiny. Ważną rolę w regulacji przepływu odgrywają nerki. Niedostateczny dopływ krwi do nerek prowadzi do uwolnienia przez nie reniny. Renina działa na hipertensynogen - składową frakcji globulinowej plazmy krwi. Wskutek tej reakcji powstaje angiotensyna (hipertensyna, angiotonina), która powoduje uogólniony skurcz naczyń i podwyższenie ciśnienia krwi. Jednocześnie dochodzi do uwolnienia aldosteronu z rdzenia nadnerczy i zahamowania zwrotnego wchłaniania wody w kanalikach nerkowych.
Płyn tkankowy i limfa Jak wiemy, krew krąży w naczyniach krwionośnych i nie styka się bezpośrednio z komórkami (poza komórkami wątroby i śledziony). Komórki natomiast tkwią w płynie tkankowym, który jest pośrednikiem między krwią i komórkami. Powstawanie i resorpcja płynu tkankowego Wspomniano, że ciśnienie w końcowych tętnicach i naczyniach włosowatych wynosi 32 mm Hg. Ciśnienie zmniejsza się stopniowo wzdłuż przebiegu naczyń włosowatych do ok. 12 mm Hg. Jak wiemy, naczynia włosowate przepuszczają składniki krwi z wyjątkiem składników morfotycznych krwi, plazmy i białek. Ciśnienie krwi powoduje powstanie siły przemieszczającej
i Ryc. 183. Schemat krążenia płynu tkankowego w otoczeniu naczynia włosowatego. W początkow y m odcinku naczynia płyn przechodzi do przestrzeni międzykomórkowej, w odcinku końcowym - z przestrzeni międzykomórkowej powraca do włośniczki. A - t ę t n i c z k a , 0- naczynie włosowate, C-żyła.
'387
płyn przez ściany naczyń włosowatych do otaczających tkanek. Obecność białek w naczyniach i brak ich w płynie otaczającym jest powodem powstawania ciśnienia osmotycznego o wartości 25 mm Hg, które powoduje przechodzenie płynu do naczyń. Istnieją zatem dwie przeciwstawne siły - ciśnienie krwi w naczyniach włosowatych i ciśnienie osmotyczne. W początkowych odcinkach naczyń włosowatych przeważa ciśnienie krwi i płyn przechodzi do tkanek, w odcinkach końcowych przewagę ma ciśnienie osmotyczne, które wtłacza płyn ponownie do naczyń. Płyn tkankowy jest zatem wydzielany w początkowych odcinkach naczyń włosowatych, a absorbowany w odcinkach dalszych. Obrzęk Stan nadmiernego wytwarzania lub wadliwego wchłaniania płynu prowadzi do jego nagromadzenia się w tkankach, co nosi nazwę obrzęku (oedema Obrzęk może wystąpić wskutek działania wielu czynników chorobowych wywołujących wzrost ciśnienia w naczyniach włosowatych i zmniejszenie ciśnienia osmotycznego, zależnego od składu białek plazmy krwi.
•iii
mm
Ryc. 184. Krążenie chłonki w naczyniu limfatycznym. Skurcz mięśnia powoduje przetłoczenie chłonki do wyższych odcinków naczyń.
'388
Ciśnienie włośniczkowe wzrasta w przypadku rozszerzenia tętniczek (spadek ciśnienia w tętniczkach powoduje wzrost ciśnienia w naczyniach włosowatych). W przypadku uszkodzenia ścian naczyń włosowatych, np. wskutek ogólnego niedotlenienia, ściany ich przepuszczają białka. Wynikiem jest zmniejszenie ciśnienia osmotycznego i upośledzenie wchłaniania płynu z przestrzeni międzykomórkowej. Taki jest mechanizm obrzęków po ukąszeniu owadów. Uszkodzenie naczyń włosowatych jest częściowo wynikiem uwolnienia histaminy. Obrzęk może powstać z powodu zatrzymania sodu wskutek zaburzeń w wydalaniu chlorku sodowego. Sól przechodząca do płynu tkankowego „przyciąga w o d ę " .
Płyn z przestrzeni międzykomórkowych odpływa do naczyń chłonnych, a wraz z nim usuwane są z przestrzeni międzykomórkowej białka, których nagromadzenie prowadzi do zatrzymywania płynu wskutek zwiększenia ciśnienia osmotycznego w przestrzeni pozanaczyniowej. Wzrost ciśnienia w naczyniach włosowatych, występujący podczas wysiłku fizycznego, jest wynikiem rozszerzenia tętniczek. Powoduje on zwiększone przechodzenie płynu do tkanek. Z drugiej strony rozkład dużych cząsteczek glikogenu i zwiększenie produktów przemiany materii prowadzą do wzrostu ciśnienia osmotycznego w płynie tkankowym. Z tego powodu ciężko pracujące mięśnie zatrzymują wodę, stają się sztywne i bolesne. Delikatne ćwiczenia i masaż mięśni otwierają naczynia chłonne, którymi odpływa nadmiar płynów zatrzymanych przez mięśnie. Zamknięcie lub uszkodzenie naczyń chłonnych powoduje powstanie obrzęku. Płyn tkankowy gra dużą rolę w odtwarzaniu objętości krwi utraconej w następstwie krwotoku. Krwotok, jak wiadomo, wiąże się ze skurczem tętniczek. Jego wynikiem jest spadek ciśnienia we włośniczkach, co z kolei powoduje zmniejszenie objętości płynu przechodzącego do tkanek i zwiększenie wchłaniania zwrotnego.
Krążenie płucne Krew powracającą do prawego przedsionka prawa komora pompuje do płuc, skąd powraca ona do lewej połowy serca. Naczynia płucne stawiają mniejszy opór niż tętniczki i naczynia włosowate krążenia wielkiego. W płucach nie ma zatem naczyń „ o p o r o w y c h " (tętniczek) w postaci występującej w krążeniu wielkim. Opór w krążeniu płucnym stawiają mniejsze gałęzie tętnic płucnych, naczynia włosowate i przynależne do nich żyły. Wynikiem małego oporu jest mniejsze ciśnienie wytwarzane przez prawą komorę, która tłoczy tę samą objętość krwi co komora lewa. Ciśnienie w tętnicach płucnych wynosi zaledwie ok. 25/8 mm Hg, podczas gdy ciśnienie w dużych tętnicach krążenia wielkiego wynosi, jak wiemy, ok. 120/80 mm Hg. Ciśnienie w żyłach płucnych jest takie samo, jak w sąsiadujących z sercem żyłach krążenia wielkiego, tj. ok, 0 m m Hg. Naczynia płucne mają cienkie ściany i są rozciągliwe, stąd ich duża pojemność. W naczyniach płucnych znajduje się ok. 1 l krwi, tj. 1 / 5 objętości krwi krążącej. Objętość krwi w płucach jest zmienna - zwiększa się podczas wdechu, równolegle ze spadkiem ciśnienia wokół naczyń (p. str. 264). Spadek '389
pojemności naczyń płucnych automatycznie zwiększa objętość krwi w naczyniach krążenia wielkiego i zwiększa rzut sercowy poprzez lepsze wypełnienie komór. Naczynia płucne zaopatrują nerwy współczule, których pobudzenie zwęża naczynia. Przy wzroście ciśnienia w naczyniach włosowatych, np. wskutek osłabienia lewej komory, dochodzi do zahamowania odpływu żylnego do lewego przedsionka. Prowadzi to do niebezpiecznego dla życia obrzęku płuc.
KREW
Krew i płyn tkankowy - limfa, tworzą środowisko wewnętrzne ustroju, w którym żyją wszystkie komórki i tkanki ciała. W związku z t y m krew ma do spełnienia szereg bardzo ważnych zadań. Polegają one na: 1) nieustannym dostarczaniu komórkom i tkankom substancji odżywczych i tlenu, 2) zabieraniu z nich produktów rozpadu, które powstają w procesach przemiany i muszą ulec wydaleniu z ustroju, 3) utrzymaniu stałości składu środowiska wewnętrznego ustroju, niezależnie od ciągle zachodzących zmian pod w p ł y w e m czynników zewnętrznych i wewnętrznych, 4) zapewnieniu łączności pomiędzy poszczególnymi częściami i narządami ustroju za pomocą różnych substancji krążących we krwi (hormony), 5) ochronie ustroju przed różnymi czynnikami szkodliwymi przenikającymi do niego z zewnątrz (ciałami obcymi, drobnoustrojami).
\\
y\—T7
//
90
80 -Surowica-
40
-Krwinki _ czerwone
W niedokrwistości (15%)
Ryc. 185. Hematokryt.
'391
Ilościowo krew u człowieka stanowi ok. 6,5% ciężaru ciała u kobiety i ok. 7% u mężczyzny. Krew jest płynem nieprzezroczystym, czerwono zabarwionym, który pozostawiony przez pewien czas w naczyniu rozdziela się na dwie warstwy. W warstwie dolnej, wskutek większego ciężaru właściwego, skupiają się komórki krwi, czyli tzw. elementy upostaciowane (morfotyczne). Należą do nich krwinki czerwone, krwinki białe oraz płytki krwi. Płynna warstwa górna, 0 opalizującym żółtawym zabarwieniu, nazywa się osoczem. OSOCZE KRWI Osocze stanowi objętościowo ok. 50-60% krwi. Stosunek objętościowy osocza do elementów upostaciowanych oznaczamy za pomocą specjalnego przyrządu, zwanego hematokrytem. Jest to wąska rurka szklana zatopiona na jednym końcu, wyskalowana na sto podziałek. Rurkę taką wypełnioną krwią z dodatkiem niewielkiej ilości substancji zapobiegającej krzepnięciu wirujemy z dużą szybkością (3500 obrotów/min) przez pół godziny. Elementy upostaciowane, jako cięższe, pod wpływem siły odśrodkowej opadają na dno, a osocze zbiera się nad zbitą warstwą krwinek. Z podziałki odczytujemy w procentach objętość obu warstw (ryc. 185). Osocze zawiera 90-92% wody i 8-10% substancji stałych. Większość substancji stałych, bo ok. 7% całego osocza, stanowią białka krwi. Białka krwi nie są jednorodne. Różnią się one między sobą ppd wieloma względami składem aminokwasowym, budową chemiczną, strukturą przestrzenną, wielkością cząsteczek, ładunkiem elektrycznym. Istnieją różne sposoby klasyfikacji białek, najczęściej oparte na metodach, jakimi się dają poszczególne grupy od siebie oddzielić. Tradycyjny podział oparty na metodzie wysalania rozróżnia trzy główne frakcje - albuminy (4%), globuliny (2,8%) i fibrynogen (0,4%). Stosując dokładniejsze i bardziej skomplikowane metody rozdziału można wyróżnić we frakcji globulinowej liczne podgrupy. Do ważniejszych należą gamma-globuliny będące nosicielami przeciwciał oraz lipoproteidy odgrywające ważną rolę w transporcie ciał tłuszczowych (lipidów). Względne rozmiary 1 ciężary cząsteczkowe białek osocza ilustruje ryc. 187. Dzięki dużej wielkości cząsteczek białka osocza w zasadzie nie przenikają przez ścianę naczyń włosowatych, podczas gdy woda i rozpuszczone w niej sole mineralne oraz inne substancje drobnocząsteczkowe przechodzą przez nią bez trudu. Białka osocza, głównie albuminy, wywierają ciśnienieonkotyczne rzędu ok. 25 mm Hg. Ciśnienie hydrostatyczne w naczyniach włosowatych wynosi ok. 38 mm Hg, przewyższa zatem ciśnienie onkotyczne o ok. 7 mm Hg. Dzięki tej różnicy odbywa się filtracja płynu przez błonę naczyń włosowatych, powodując stały niewielki prąd płynu z naczyń do przestrzeni międzykomórkowej, a stamtąd do naczyń limfatycznych. Białka osocza, głównie albuminy, przyczyniają się do utrzymywania płynu w łożysku naczyniowym. Spadek stężenia albumin w osoczu*poniżej 1 g% powoduje nadmierną ucieczkę płynu z łożyska naczyniowego i powstawanie obrzęków. 4
'392
We frakcji globulinowej odróżniamy globuliny oznaczone pierwszymi kolejnymi literami alfabetu greckiego: alfa, beta i gamma. Do gamma-globulin należą krążące we krwi przeciwciała, o których będzie jeszcze mowa. Są one produkowane przez komórki plazmatyczne. Natomiast albuminy i białka układu krzepnięcia krwi (fibrynogen, protrombina) są wytwarzane w wątrobie.
Krew
5-6 I
56%
44%
OsOC7t>
Krwinki
3 0 0 - 6 0 0 tys/mm 3
4 , 5 - 6 tys/mm 3
' ' . Cf
,
Krwinki płytkowe (tromtfocyty)
Krwinki białe (leukocyty)
ł
4 , 5 - 5 mln/mm 3
Krwinki czerwone (erytrocyty)
V
jfeJL JKmf Limfocyty 36 %
Kwasochłonne 2-5%
Obojętnochłonne 57%
Zasadochłonne 0,5 %
Ryc. 186. Skład krwi.
'393
Reszta substancji stałych - to związki organiczne i sole mineralne. Do związków organicznych należą związki azotowe: aminokwasy (zawartość azotu od 0,004 do 0,007%), mocznik (0,01-0,02%), kwas moczowy (0,00250,004%), kreatynina (0,0008-0,001%) oraz związki bezazotowe: cukier gronowy (0,08-0,12%), tłuszcze i lipidy (0,4-0,88%), kwas mlekowy (0,01-0,02%). Sole mineralne, czyli elektrolity, stanowią około 0,9-1 % osocza. Występują tu kationy: sód, potas, wapń, magnez, żelazo, oraz aniony: chlor,fosfor, siarka, jod. Do anionów zaliczamy również białka. Zawartość poszczególnych elektrolitów przedstawia tab. 29.
Na+
100 A
Albuminy 69000
y-globuliny 156000
Cl-
Hemoglobina 68000
aHipoproteidy 200000
Glukoza
j3n-globuliny 90000
fi,-lipoproteidy 1300000
Fibrynogen 400000 Ryc. 187. Białka osocza - względne wielkości cząsteczek i ciężary cząsteczkowe.
T a b e l a 29 Zawartość e l e k t r o l i t ó w / Elektrolity Na +
K+ Ca ++ Mg + +
'394
Stężenie 310-340 m g % 135-147 m m o l / l 14-22 m g % 3,5-5,5 m m o l / l 9-11 m g % 4,7-5,3 m E q / l 2,6 m g % 2,2 m m o l / l
Elektrolity
cr
Stężenie
HCO-3
340-370 m g % 97-106 m m o l / l 2,5-4,5 m g % 1,5-2 mmol/l 26-32 m m o l / l
Białko
18 m m o l / l
HP0"4
Większość soli mineralnych osocza, bo około 0,6%, stanowi chlorek sodowy - NaCI. Reszta kationów występuje w postaci węglanów, chlorków oraz nieznacznej ilości siarczanów i fosforanów. Niewielka część soli jest związana z białkami, większość jest rozpuszczalna w wodzie osocza.
ODCZYN KRWI W temperaturze ciała pH krwi wynosi 7,4, a zatem krew oddziałuje słabo zasadowo. Stałość stężenia jonów wodorowych, czyli odczynu krwi, ma wielkie znaczenie biologiczne, gdyż nawet niewielkie przesunięcia w kierunku kwaśnym lub zasadowym prowadzą do zaburzeń wielu procesów fizjologicznych. Większość procesów związanych z przemianą materii ma tendencję do zakwaszania ustroju. Istnieje szereg bardzo czułych mechanizmów stabilizujących stężenie jonów wodorowych. Należą tu przede wszystkim nerki i płuca. Płuca wydalają nadmiar kwasu węglowego w postaci C0 2 , nerki wydalają nadmiar kwasów w postaci soli kwaśnych, a nadmiar zasad w postaci węglanów, głównie sodowego. Stałość odczynu krwi zapewniają jej właściwości buforowe. Właściwości buforowe mają roztwory zawierający słaby, tj. słabo zdysocjowany na jony, kwas i sól tego kwasu z mocną zasadą. Dodanie do takiego roztworu mocnego kwasu lub ługu nie zmienia odczynu roztworu w tym stopniu, co dodanie tej samej ilości kwasu lub zasady do wody. Najważniejszym układem buforowym krwi są: wodorowęglany - kwas węglowy; w mniejszym stopniu: fosforany pierwszo- i drugorzędowe, białka krwi oraz hemoglobina -oksyhemoglobina. Dzięki tym właściwościom buforowym kwas mlekowy, fosforowy, siarkow y , węglowy, powstające w procesach przemiany, nie zmieniają w sposób widoczny pH krwi. Zmiany w odczynie krwi mogą nastąpić dopiero po wyczerpaniu się zdolności buforowych krwi, do czego prowadzą np. niektóre choroby. KRZEPNIĘCIE KRWI Wynaczyniona krew człowieka po 3 - 4 min zaczyna krzepnąć i po 5 - 6 min zamienia się w galaretowaty skrzep, który przybiera kształt naczynia, w którym odbywa się krzepnięcie. Krzepnięcie krwi polega na przemianie rozpuszczonego we krwi białka - fibrynogenu w postać nierozpuszczalną - fibrynę, czyli włóknik. Przy krzepnięciu włóknik wytrąca się w postaci cienkich, splątanych ze sobą nici, które obejmują elementy upostaciowane, wskutek czego skrzep ma barwę czerwoną. Osocze, oddzielone od elementów upostaciowanych, krzepnie tak samo, jak pełna krew. Osocze można uzyskać np. przez odwirowanie krwi w niskiej temperaturze, która zapobiega krzepnięciu. Skrzep otrzymany z odwirowanego osocza ma budowę drobnowłóknistą i białe zabarwienie. '395
Osocze krwi pozbawione włóknika nazywa się surowicą. Skrzep powstający z pełnej krwi po pewnym czasie kurczy się i wyciska część surowicy. Surowica różni się od osocza tylko brakiem fibrynogenu. Wewnątrz naczyń krwionośnych w ustroju krew nie krzepnie, jeżeli ściany naczynia nie są uszkodzone. Biologiczne znaczenie krzepnięcia polega na tym, że powstający przy przerwaniu naczynia skrzep tworzy tampon, który zapobiega dalszemu krwawieniu. W krwawiączce (hemofilii), chorobie polegającej na wrodzonym zaburzeniu krzepliwości krwi, nawet niewielkie zranienie może doprowadzić do skrwawienia śmiertelnego. Mechanizm krzepnięcia krwi jest bardzo złożony i wciąż jeszcze nie całkowicie wyjaśniony. Istota jego polega na szeregu kaskadowych reakcji, w których bierze udział co najmniej 30 różnych substancji. Upraszczając jednak sprawę, można wyróżnić trzy główne etapy krzepnięcia krwi - pierwszy polega na powstaniu substancji zwanej aktywatorem protrombiny w odpowiedzi na przerwanie naczynia lub zmiany w samej krwi. W drugim - aktywator protrombiny katalizuje przemianę protrombiny na trombinę. W trzecim - trombina działa jako enzym zamieniający fibrynogen na fibrynę, czyli włóknik. Ten ostatni ma postać gęstej sieci złożonej z długich włókien (stąd nazwa), przebiegających w różnych kierunkach, w której zostają uwięzione elementy komórkowe (krwinki czerwone i białe) tworząc galaretowaty konglomerat zwany skrzepem. W procesie tym niezbędne są jony wapnia. Wytrącenie jonów wapnia, np. kwasem szczawiowym, zapobiega krzepnięciu. Taka krew szczawianowa krzepnie po dodaniu rozpuszczalnych soli wapnia, np. CaCI2. Jeżeli krzepnięcie krwi w miejscu przerwania ciągłości naczynia jest reakcją korzystną, chroniącą ustrój przed wykrwawieniem, to powstawanie skrzepu wewnątrz naczynia jest zjawiskiem patologicznym, wysoce niebezpiecznym, prowadzącym niekiedy do groźnych dla życia następstw. Zatkanie przeż zakrzep tętnicy prowadzi do martwicy obszaru przez tę tętnicę zaopatrywanego, co nazywamy zawałem. Obok mechanizmów krzepnięcia ustrój rozporządza jeszcze mechanizmami, które stale przeciwstawiają się i zapobiegają powstawaniu skrzepów w naczyniach. Jednym z takich mechanizmów jest układ fibrynolityczny, który ogranicza krzepnięcie w ten sposób, iż prowadzi do rozpuszczania powstającego włóknika.
Substancje przeciwkrzepliwe Substancje zapobiegające lub utrudniające powstawanie skrzepu noszą nazwę środków przeciwkrzepliwych. Krzepnięciu krwi in vitro można, jak wspomniano, zapobiec usuwając z niej jony wapnia. Używa się w tym celu substancji wiążących wapń, takich jak szczawian, cytrynian lub wersenian sodowy. '396
Silnym środkiem przeciwkrzepliwym działającym in vivo i in vitro jest heparyna. Jest to sulfonowany wielocukier wytwarzany w komórkach tucznych, które znajdują się w licznych tkankach. Działanie jej polega na hamowaniu trombiny. Do leków przeciwkrzepliwych stosowanych w klinice należą także pochodne kumaryny, które hamują syntezę protrombiny w wątrobie.
LIMFA Limfą (chłonką) nazywamy płyn tkankowy, który wchodzi z przestrzeni międzykomórkowych do naczyń limfatycznych. Skład limfy jest zbliżony do składu osocza, z tym że zawiera ona znacznie mniej białka. Zastój limfy doprowadza do jej krzepnięcia i powstawania obrzęków. Limfa z licznych naczyń limfatycznych dostaje się ostatecznie prze przewód piersiowy (ductus thoracicus) do krwi żylnej. Na drodze naczyń limfatycznych znajdują się liczne węzły chłonne (limfatyczne) wytwarzające limfocyty. Stąd pochodzi większa część limfocytów znajdujących się we krwi. Do naczyń limfatycznych krezki w jamie brzusznej wchłaniane są tłuszcze w postaci tzw. chylomikronów; nadają one limfie odpływającej z jelit podczas trawienia tłuszczu wygląd mlećzny.
ELEMENTY UPOSTACIOWANE KRWI
Krwinki czerwone (erytrocyty) Krwinki czerwone mają kształt dwuwklęsłych krążków o średnicy ok. 7,5 grubości ok. 2 [xm. Widziane z boku mają kształt biszkopta (ryc. 188). Tego rodzaju kształt zwiększa powierzchnię krwinek. Krwinki czerwone są wytwarzane w szpiku kostnym czerwonym. U człowieka przed wydostaniem się do krwi krwinki czerwone tracą jądro komórkowe. Liczba krwinek czerwonych w 1 mm 3 wynosi ok. 5,4 min u mężczyzny i ok. 4,8 min u kobiety. Krwinki czerwone żyją ok. 120 dni. Pełnią one w tym czasie ważną rolę w przenoszeniu gazów krwi - tlenu i dwutlenku węgla, dzięki zawartemu w nich barwnikowi - hemoglobinie. 2,2 jjm
Ryc. 188. Wielkość i kształt krwinki czerwonej.
'397
Ryc. 189. Krwinki czerwone układające się w rulony.
Komórka wątrobowa, w której następuje sprzęganie bilirubiny z kwasem glukuronowym
Komórka siateczkowo-śródbłonkowa gwiaździsta Bilirubina pośrednia nie przechodzi przez nerkę
Pęcherzyk żółciowy
Jelito
O
Śledziona
Krwinka czerwona
Kał zabarwiony na brązowo Bilirubina pośrednia <£) nierozpuszczalna w wodzie
Kw. glukuronowy ^
Urobilinogen
Bilirubina bezpośrednia 0 związana z kw. glukuronowym rozpuszczalna w wodzie
Ryc. 190. Model przemiany hemoglobiny. W układzie siateczkowo-śródbłonkowym (USŚ), śledzionie, w komórkach siateczkowo-śródbłonkowych gwiaździstych hemoglobina ulega przemianie na bilirubinę, bilirubina ta jest nierozpuszczalna w wodzie; w wątrobie zostaje ona połączona z kwasem glukuronowym i zamienia się w postać rozpuszczalną, która przechodzi do żółci. W jelicie grubym pod działaniem bakterii bilirubina zamienia się w urobilinogen.
'398
Hemoglobina jest białkiem o ciężarze cząsteczkowym ok. 68 000. Hemoglobina wiąże tlen tworząc tzw. oksyhemoglobinę, przy czym cząsteczka 0 2 jest dosyć luźno związana z żelazem dwuwartościowym. Reakcji tej nie należy utożsamiać z utlenieniem. Przy zadziałaniu na krew w organizmie bądź poza nim środków utleniających dwuwartościowy jon Fe ++ zamienia się na jon trójwartościowy Fe + + + i taka hemoglobina, zwana methemoglobiną, nie może już pełnić roli przenośnika tlenu. Związana z tlenem hemoglobina, zwana oksyhemoglobiną, ma kolor żywoczerwony nadający zabarwienie właściwe krwi tętniczej. Przeciętna zawartość hemoglobiny we krwi wynosi ok. 16 g w 100 ml u mężczyzny i ok. 14g/100 ml u kobiety. Zatem mężczyzna ważący 70 kg ma we krwi ok. 900 g hemoglobiny. Ponieważ za okres życia krwinki czerwonej można przyjąć 120 dni, każdego dnia ulega rozpadowi ok. 7,5 g hemoglobiny i tyleż musi na nowo powstać. ^ „Wysłużone" krwinki czerwone są niszczone w tzw. układzie siateczkowo-śródbłonkowym, głównie w śledzionie. Następuje przy tym odłączenie od hemoglobiny części zwanej hemem i rozerwanie pierścienia hemowego, który zamienia się na biliwerdynę. U człowieka większa część biliwerdyny ulega przekształceniu na bilirubinę, która w wątrobie zostaje sprzężona z kwasem glukuronowym i wydalona z żółcią. Wyzwolone przy rozpadzie hemu żelazo zostaje zatrzymane w ustroju i zużyte do ponownej syntezy hemu. Przy dużych stratach krwi dochodzi równocześnie do strat żelaza i tzw. niedokrwistości niedobarwliwej. Brak żelaza uniemożliwia wyprodukowanie dostatecznej ilości barwnika krwi - hemoglobiny. Niewielka część bilirubiny związanej z kwasem glukuronowym przenika do krwi - jest to tzw. bilirubina bezpośrednia, natomiast bilirubina wolna, która nie przeszła przez wątrobę, jako trudniej rozpuszczalna w wodzie stanowi bilirubinę pośrednią. W warunkach prawidłowych poziom bilirubiny całkowitej (bezpośredniej i pośledniej) we krwi nie przekracza 1,5 mg w 100 ml. Przy wyższym jej stężeniu (ponad 2mg%) następuje odkładanie się tego barwnika w tkankach i skórze powodując żółtaczkę. Przyczyną żółtaczki może być nadmierne wytwarzanie bilirubiny lub utrudnione jej wydalanie przez wątrobę lub drogi żółciowe. W żółtaczce spowodowanej hemolizą krwinek czerwonych wzrasta poziom bilirubiny pośredniej.
Krwinki płytkowe (płytki krwi, trombocyty) Płytki krwi są to małe (średnicy 2 - 4 (im) fragmenty cytoplazmy oderwane od olbrzymich komórek szpiku, tzw. megakariocytów. Krwinki płytkowe biorą udział w procesie krzepnięcia krwi; zawierają one dużą ilość substancji obkurczającej naczynia krwionośne-serotoniny. Krwinki płytkowe gromadzą się w miejscach uszkodzenia naczyń, gdzie przylegają do uszkodzonej ściany naczyniowej i uwalniają serotoninę, która powoduje lokalny skurcz ściany naczyniowej zmniejszając krwawienie. 1 mm 3 krwi zawiera ok. 300 000 płytek. '399
SZPIK KOSTNY Szpik kostny wypełniający jamy szpikowe kości jest miejscem powstawania krwinek czerwonych, granulocytów, płytek krwi oraz w małym stopniu - t a k ż e limfocytów i monocytów. Do 20 roku życia szpik jest czynny we wszystkich kościach. Po tym okresie szpik wypełniający jamy szpikowe kości długich staje się nieczynny. Wskutek nagromadzenia się w nim tłuszczu czynny „szpik czerwony", zamienia się na nieczynny „szpik żółty". Szpik zawiera dwa szeregi komórek - układu białokrwinkowego (75%) i czerwonokrwinkowego (25%). Komórki te stanowią różne etapy wykształcania się ostatecznych komórek krwi. Określanie ich wzajemnych proporcji ilościowych ma duże znaczenie w hematologii.
KRWINKI BIAŁE 1 mm 3 krwi w warunkach prawidłowych zawiera 4000-11 000 krwinek białych (leukocytów). Liczba ta stanowi tylko małą część ogólnych zasobów tych krwinek w ustroju. Duże ich zapasy są stale zgromadzone w szpiku kostnym, śledzionie i węzłach chłonnych, skąd pod wpływem odpowiedniego bodźca mogą być natychmiast wyrzucone do krwi, gdzie ich liczba może się zwiększyć nawet dziesięciokrotnie. Pod wpływem odpowiednich bodźców może się także zwiększyć wytwarzanie leukocytów. W zależności od wyglądu i budowy dzielimy leukocyty na kilka rodzajów. Największą grupę stanowią granulocyty, zwane tak dlatego, że zawierają w cytoplazmie liczne ziarnistości. Zależnie od sposobu, w jaki owe ziarnistości się barwią, odróżniamy granulocyty obojętnochłonne, kwasochłonne i zasadochłonne. Najwięcej, bo 50-70% całkowitej liczby krwinek białych, jest granulocytów obojętnochłonnych. Granulocyty kwasochłonne stanowią 14%, zaś zasadochłonne - tylko ok. 0,4% całkowitej liczby krwinek białych. Młode granulocyty mają jedno jądro w kształcie pałeczki lub podkowy; w miarę dojrzewania jądra granulocytów stają się wielopłatowe (segmentowane). Oprócz granulocytów we krwi obwodowej znajdują się jeszcze dwa rodzaje krwinek białych jednojądrzastych. Są to: 1) limfocyty - komórki o dużym okrągłym jądrze i małej ilości cytoplazmy (20-40% wszystkich leukocytów) oraz 2) monocyty - komórki o dużej ilości bezziarnistej cytoplazmy i jądrze okrągłym lub owalnym, podobnym do kłębka jedwabiu. Granulocyty są wytwarzane w szpiku kostnym. Komórki obojętnochłonne wchodzą do układu krążenia i przebywają we krwi przeciętnie ok. 7 h. Ich całkowity okres przeżycia wynosi ok. 30 h. Duża liczba tych krwinek przenika do przewodu pokarmowego i opuszcza tą drogą ustrój. Granulocyty obojętnochłonne mogą się przedostawać przez ścianę naczyń włosowatych do tkanek. Proces ten jest samoistny i nazywa się diapedezą, a następuje zwykle pod '400
w p ł y w e m pewnych związków chemicznych powstających w zakażonych tkankach. Granulocyty te w dużej ilości otaczają bakterie lub inne ciała obce usiłując je zniszczyć. W ognisku zakażenia i wokół niego wytwarza się zwykle płyn złożony z milionów fagocytów - w większości krwinek białych obojętnochłonnych, bakterii i produktów ich rozpadu, zwany ropą. W przebiegu zakażenia ustroju bakteriami zwiększa się liczba granulocytów krążących we krwi, co spowodowane jest mobilizacją ich do krwi przez czynnik zakaźny.
Limfocyty Większa część limfocytów wytwarzany jestwwęzłach chłonnych (limfatycznych) i śledzionie; stosunkowo niewielka c z ę ś ć - w szpiku kostnym. Limfocyty należą do komórek immunokompetentnych - tj. zdolnych do wytwarzania przeciwciał, o czym będzie mowa dalej. Odróżnia się limfocyty małe i duże. Limfocyty małe (średnicy 6-10 jim) wyglądają prawie jak jądra komórkowe otoczone znikomą ilością cytoplazmy. Limfocyty duże mają średnicę ok. 12-16 [xm o jądrze jajowatym i nieco większej ilości przejrzystej cytoplazmy.
Monocyty Monocyty są czynnymi fagocytami, podobnie jak granulocyty obojętnochłonne. UKŁAD SIATECZKOWO-ŚRÓDBŁONKOWY Wspólną nazwą układu siateczkowo-śródbłonkowego obejmuje się komórki obdarzone zdolnością fagocytarną występujące w wątrobie (komórki Kupffera), śledzionie, szpiku kostnym, węzłach chłonnych. Komórki te pochłaniają duże cząsteczki koloidalne, co wykorzystane zostało do badań scyntygraficznych wątroby. Radioaktywne złoto w roztworze koloidalnym, wstrzyknięte dożylnie, wychwytywane jest przez te komórki w wątrobie, co pozwala uwidocznić wielkość, kształt i zlokalizować ewentualne guzy w wątrobie.
MECHANIZMY OBRONNE KRWI Jeżeli do ustroju dostanie się obce białko, a to jest bardzo często równoznaczne z wtargnięciem niebezpiecznych dla życia drobnoustrojów chorobotwórczych, następuje wówczas szereg reakcji obronnych ujmowanych ogólną nazwą reakcji immunologicznej. Istota jej polega na wytworzeniu substancji, które reagują z wtargniętym do ustroju ciałem obcym, tak aby je unieszkodliwić i wydalić. 26 - Anatomia i fizjologia
401
Antygeny Czynniki wywołujące reakcje immunologiczne nazywamy antygenami. Pod względem chemicznym są to najczęściej substancje białkowe połączone z lipidami lub węglowodanami. Niekiedy antygenem może być także substancja niebiałkowa; przypuszcza się jednak, że po to, aby wywołać reakcję immunologiczną, musi ona najpierw połączyć się z jakimś białkiem ustroju. Ustrój rozpoznaje własne białka i nie reaguje na nie, jeżeli jednak takie własne białko połączy się z jakąś inną substancją obcą, np. penicyliną, wówczas staje się dla ustroju „ o b c e " - nabiera cech antygenu. W niektórych sytuacjach, z przyczyn niezupełnie jeszcze poznanych, antygenem może sięstaćwłasne białko. Powstająca wówczas reakcja immunologiczna staje się podłożem tzw. chorób autoimmunologicznych (chorób z autoagresji).
Przeciwciała Wtargnięcie do ustroju antygenu pociąga za sobą wytwarzanie swoistego przeciwciała, którym jest białko o budowie chemicznej dopasowanej do danego antygenu „jak klucz do zamka". Przeciwciała takie mogą krążyć w surowicy krwi we frakcji gamma-globulinowej albo też pozostawać związane z niektórymi tkankami. Mechanizm powstawania przeciwciał Mechanizm powstawania przeciwciał jest dziś przedmiotem intensywnych badań i w ostatnich latach dokonano w tej dziedzinie dużego postępu. Rzecz w tym, iż te, skądinąd bardzo korzystne dla organizmu reakcje obronne, stanowią równocześnie największą przeszkodę dla skutecznego przeszczepiania narządów, powodując odrzucanie przeszczepów. Według dzisiejszych poglądów kluczową rolę w t y c h reakcjach.odgrywają limfocyty. Antygen styka się w ustroju z milionami limfocytów, które mają różnorodnie ukształtowane struktury chemiczne na swej powierzchni. Zetknięcie się limfocytu, którego ukształtowanie powierzchni „pasuje" do struktury chemicznej antygenu, pobudza (warunkuje) limfocyt do szybkiego mnożenia się i produkcji przeciwciał. Limfocyty ulegają przy tym przemianie na komórki plazmatyczne i te dopiero są miejscem wytwarzania przeciwciał wyzwalanych do krwi w postaci swoistej globuliny. Z przyczyn dotychczas nie znanych zdarza się niekiedy tak, że jedna taka komórka plazmatyczna zaczyna się rozmnażać w sposób nie kontrolowany i wypierając niejako wszystkie inne plazmocyty nabiera cech nowotworu zwanego plasmocytoma (szpiczak). Takie komórki produkują w dużej ilości jeden rodzaj białka o charakterze przeciwciała, niepotrzebnego zresztą ustrojowi. Uzyskane właśnie z takich przypadków duże ilości jednorodnego białka umożliwiły badaczom dokładne poznanie budowy chemicznej i struktury przestrzennej przeciwciał. '402
Grasica Grasica opisana w innym miejscu (p. str. 515), we wczesnym okresie życia, jest pierwotnym źródłem limfocytów, które wywędrowują z niej, tworząc w różnych miejscach kolonie - w węzłach chłonnych, śledzionie, szpiku kostnym, migdałkach, wyrostku robaczkowym.
Reakcja antygen - przeciwciało Reakcja przeciwciała z antygenem jest skierowana na zniszczenie lub unieszkodliwienie antygenu. Jeżeli antygen jest związany z przeszczepioną tkanką, następuje obumarcie i odrzucenie przeszczepu. Jeżeli antygen zlokali-
Ryc. 191. Schemat działania przeciwciał - wiązanie antygenów powierzchniowych przez przeciwciała w kompleksy ulegające wytrąceniu.
'403
zowany jest na powierzchni pojedynczej komórki bakteryjnej w jej błonie, to pod wpływem swoistego przeciwciała może nastąpić jej rozpuszczenie, unieruchomienie, obumarcie, aglutynacja lub zostaje umożliwiona fagocytoza pożarcie obcej komórki przez makrofagi. Często reakcja antygen-przeciwciało wywołuje w ustroju duże zaburzenia, dające znane objawy chorób uczuleniowych, czyli alergicznych. Dzieje się to zwłaszcza wtedy, gdy przeciwciała są związane z komórkami w naczyniach krwionośnych lub drogach oddechowych. W wyniku reakcji antygen-przeciwciało następuje uszkodzenie komórek gospodarza. Wydzielają się przy t y m pewne substancje (histamina, serotonina), które powodują np. skurcz oskrzeli (dychawica oskrzelowa), rozszerzenie naczyń błony śluzowej nosa (katar sienny), uszkodzenie naczyń włosowatych skóry (pokrzywka). Pomimo tych niekiedy niekorzystnych zjawisk ubocznych reakcji antygen-przeciwciało odpowiedź immunologiczna ustroju jako podstawowy mechanizm obronny jest niezwykle ważna dla życia. Młode zwierzęta, którym usunięto grasicę, pozostają niezdolne do wytwarzania przeciwciał i szybko giną z powodu inwazji drobnoustrojów. Przed śmiercią może je uchronić tylko hodowla w warunkach całkowicie jałowych.
GRUPY KRWI I PRZETACZANIE KRWI Odczyny immunologiczne, mające na celu obronę organizmu przed inwazją ciał obcych, jak już wspomniano, stoją na przeszkodzie w przeszczepianiu tkanek od jednego osobnika do drugiego. Pierwszą taką tkanką, którą spróbowano przeszczepiać w większej ilości z jednego organizmu do drugiego/była krew. Problemy immunologiczne, jakie przy tym wynikły, okazały się stosunkowo proste. Stwierdzono, że białka osocza u różnych ludzi są pod względem antygenowym tak bardzo do siebie podobne, że po przetoczeniu do innego ustroju nie wywołują powstawania przeciwciał. Inaczej ma się rzecz z krwinkami czerwonymi. Pod względem antygenowym krwinki czerwone różnych ludzi różnią się między sobą. Zależnie od tego, jaki rodzaj antygenu ma na swej powierzchni krwinka czerwona, istnieje szereg różnych gatunków krwinek. Z praktycznego punktu widzenia ważne są jednak tylko trzy antygeny, które oznacza się symbolami A, B i Rh, ponieważ tylko te antygeny wywołują poważniejsze reakcje immunologiczne. Inne antygeny, których liczba sięga już siedemdziesięciu, są tak „słabe", że można ich nie brać pod uwagę, chociaż przed każdym przetoczeniem wykonuje się tzw. próbę krzyżową, o czym wspomnimy dalej. W świetle tego, co powiedzieliśmy wyżej o reakcji antygen-przeciwciało, jest rzeczą oczywistą, że surowica krwi nie może zawierać przeciwciał przeciwko własnym krwinkom. Ponieważ przeciwciała te powodują zlepianie się krwinek "zerwonych (aglutynację), nazywamy je aglutyninami. Z przyczyn niezupełnie jasnych we krwi osób, których krwinki nie posiadają antygenu A lub B, wkrótce po urodzeniu powstają przeciwko nim przeciwciała, mimo że '404
osoby te nie były przeciwko nim uczulone. Reasumując: ze względu na rodzaj antygenu zawartego w krwinkach czerwonych odróżnia się cztery różne grupy krwi: A, B, AB i 0. Osoba z grupą krwi A zawiera w surowicy przeciwciała (aglutyniny) anty B. Osoba z grupą krwi B - zawiera w surowicy aglutyniny anty A. Osoba z grupą krwi AB - nie zawiera w surowicy aglutyniny anty A, ani anty B. Osoba z grupą krwi 0 - zawiera aglutyniny zarówno anty A, jak i anty B. Nieco inaczej przedstawia się sprawa z antygenem Rh. Krwinki każdej z tych grup (A, B, AB i 0) mogą zawierać jeszcze antygen Rh. Osoby bez tego antygenu w krwinkach nie mają przeciwciała anty Rh, dopóki nie przetoczy im się krwinek czerwonych zawierających ten antygen. Każde następne przetoczenie krwinek Rh + wywoła już ich aglutynację.
/
0
• $ • A (anty B)
B (anty A)
A (anty B)
B (anty A)
'
A (anty B)
•
A (anty B)
A
B
•
B (anty A)
w
AB
B (anty A)
Ryc. 192. Oznaczanie grup krwi. 1- Nie ma aglutynacji ani w surowicy grupy B (anty A) - k r w i n k i należą do grupy 0 . 2 - Aglutynacja występuje tylko w surowicy grupy B (anty A) - k r w i n k i należą do grupy krwi A. 3- Aglutynacja w surowicy grupy A (anty B) - krwinki należą do krwi grupy B. 4Aglutynacja w obu surowicach wzorcowych (anty A i anty B) - krwinki należą do krwi grupy AB.
'405
Z w y w o d ó w tych wynikają reguły, jakimi kierujemy się przy przetaczaniu krwi: 1. Można przetaczać krew jednoimienną. 2. Osoba z grupą krwi AB jest uniwersalnym biorcą, ponieważ nie ma w surowicy przeciwciał anty A ani anty B, można jej podać krew wszystkich grup. 3. Osoba z grupą krwi 0 jest uniwersalnym dawcą. Krew tej grupy można przetaczać wszystkim biorcom. 4. Krew grupy A można przetaczać biorcy z grupą A i AB. 5. Krew grupy B można przetaczać biorcy z grupą B i AB. 6. Krew grupy AB można przetaczać tylko biorcy z grupą AB. 7. Osobom z grupą Rh" (Rh minus) nie należy przetaczać krwi grupy Rh + . Z powyższego zestawienia widać, że „najlepsza" dla celów transfuzji jest krew grupy 0 (Rh"). Trzeba jednak pamiętać o tym, że osocze grupy 0 zawiera przeciwciała anty A i antyB i przetoczenie bardzo dużej ilości takiej krwi może spowodować aglutynację krwinek własnych biorcy. W każdym przypadku przed przetoczeniem krwi wykonuje się tzw. próbę krzyżową, polegającą na zmieszaniu krwi dawcy z osoczem lub surowicą biorcy. Oznaczanie grup krwi wykonuje się na szkiełku podstawowym lub na białej płytce porcelanowej w ten sposób, iż miesza się badane krwinki z wzorcowymi surowicami znanej grupy i surowicą anty Rh o wysokim mianie przeciwciał i obserwuje się, z jakim wzorcem następuje aglutynacja. Szczegółowe przepisy znajdują się w podręcznikach technik laboratoryjnych.
UKŁAD N E R W O W Y
Jedną z podstawowych właściwości żywej materii jest pobudliwość, tj. zdolność reagowania na bodźce w sposób zapewniający ciągłość procesów życiowych. Zdolnością tą w najwyższym stopniu obdarzony jest układ nerwow y , dzięki czemu zajmuje on w ustroju szczególne miejsce. Układ nerwowy utrzymuje łączność organizmu ze światem zewnętrznym. Łączność ta jest niezbędna dla życia, gdyż ze środowiska zewnętrznego ustrój czerpie substancje dla budowy własnego ciała i podtrzymania podstawowych procesów życiowych, jak również dlatego by uniknąć czynników tego środowiska zagrażających życiu. Układ nerwowy integruje czynności poszczególnych narządów i układów narządów tworząc z organizmu jedną całość, której składniki harmonijnie ze sobą współpracują. Czynności układu nerwowego umownie dają się podzielić na dwie kategorie - jedna z nich, nastawiona na łączność ze światem zewnętrznym, odbiera zeń różnorodne informacje za pośrednictwem narządów zmysłów oraz zarządza aparatem ruchowym, umożliwiając poruszanie się w przestrzeni i reagowanie w sposób celowy na bodźce zewnętrzne; druga - polega na regulowaniu przemiany materii i czynności narządów wewnętrznych organizmu. Czynnościami pierwszej kategorii zarządza układ nerwowy somatyczny (soma-ciało), drugiej - u k ł a d nerwowy autonomiczny ( a u t o s - s a m + nomos - prawo, tj. samorządzący się), zwany także układem wegetatywnym. Działalność obu tych układów jest koordynowana na wszystkich piętrach układu nerwowego.
NEURON Podstawowym elementem układu nerwowego jest komórka nerwowa z odchodzącymi od niej wypustkami. W roku 1891 niemiecki anatom, W. Waldayer, nazwał taki element neuronem. Rycina 194 przedstawia powiększony schemat neuronu. Widnieje na nim bańkowate ciało komórkowe, od którego na obwodzie odchodzi szereg rozgałęziających się wypustek. Liczne krótkie wypustki tworzące drzewkowate rozgałęzienia nazywają się dendrytami. Jedna wypustka jest długa, pokryta zwykle tłuszczową jasną otoczką, tzw. osłonką mielinową, i nosi nazwę neurytu lub aksonu. Niektóre aksony osiągają długość jednego metra. Pojęcie o wzajemnych proporcjach poszczególnych części neuronu może dać następujące porównanie: gdyby ciało neuronu, którego akson biegnie do mięśni stopy, miało wielkość piłki tenisowej, to dendryty zajęłyby przestrzeń '407
równą pokojowi mieszkalnemu średniej wielkości, a akson miałby długość ok. 1,5 km przy średnicy przekroju 1 cm. W rzeczywistości maksymalna przestrzeń, jaką zajmuje neuron ze wszystkimi wypustkami w ośrodkowym układzie nerwowym, nie przekracza kilku tysięcznych cm 3 . Układ nerwowy zbudowany jest z olbrzymiej ilości neuronów - liczbę ich szacuje się na ok. 30 mld. Pod względem czynnościowym neurony dzielą się na czuciowe, ruchowe i pośrednie. Dla łatwiejszego zrozumienia zasady funkcjonowania układu nerwowego porównuje się go często do komputera elektronowego. Neurony ruchowe można wówczas nazwać urządzeniami wyjściowymi, neurony czuciowe urządzeniami wejściowymi, zaś wszystko, co jest pomiędzy nimi, stanowią neurony pośrednie. Niektóre neurony czuciowe zaopatrzone są na obwodzie w specjalne urządzenia - czujniki, przystosowane do odbierania i przekazywania tych informacji, które potrafią przyjmować nasze zmysły, a więc temperatury, ciśnienia, światła, barwy, składu chemicznego pokarmu (smak), powietrza (zapach) itp. Zadanie neuronów czuciowych polega na odebraniu i ocenie siły tych wszystkich bodźców i przekształceniu ich w rodzaj elektrochemicznego sygnału, który zostaje następnie przesłany dla dalszego przetworzenia w układzie nerwowym. Tak np. pręciki i czopki w siatkówce oka zawierają pewną substancję chemiczną, która pod w p ł y w e m światła ulega rozpadowi, podobnie jak związki srebra w kliszy fotograficznej. Produkt tego rozpadu stanowi bodziec powodujący zmiany elektryczne w neuronie, przekazywane do odpowiednich ośrodków w mózgu. Inne rodzaje wyspecjalizowanych czujników, zwane w fizjologii receptorami, odbierają i mierzą natężenie takich działań, jak ciepło, zimno, ból itd. Wszystkie tak bardzo różne rodzaje informacji zostają przetworzone w jeden rodzaj bodźców elektrycznych - podobnie jak w komputerze elektronowym wszystkie wprowadzone informacje muszą być przełożone na odpowiedni język dający się zamienić na impulsy elektryczne lub w telewizji - obraz i dźwięk - muszą zostać zamienione na fale elektromagnetyczne. Takie same impulsy elektryczne wysyłają neurony ruchowe, czyli efektorowe do mięśni. Impulsy te zostają „odczytane" jako odpowiednie rozkazy dla skurczu poszczególnych mięśni lub grup mięśniowych dających w efekcie zamierzone ruchy. Aby zrozumieć mechanizm działania układu nerwowego, należy dokładnie zapoznać się z czynnością pojedynczego neuronu. Różne jego części mają różne funkcje. Tak więc neuryt, czyli akson, przewodzi impulsy powstające w ciele komórkowym do jakiegoś innego miejsca, najczęściej do innego neuronu lub włókna mięśniowego. Akson można porównać do kabla tworzącego pewien obwód elektryczny. Po dojściu do miejsca przeznaczenia akson dzieli się na liczne gałązki, które nawiązują kontakt z dendrytami lub bezpośrednio z ciałem komórkowym innego neuronu. W miejscu takiego kontaktu znajduje się specjalne urządzenie do przekazywania bodźców z jednego neuronu na drugi, zwane synapsą. '408
Proces przewodzenia bodźców we włóknach nerwowych jest zupełnie niepodobny do rozchodzenia się impulsów w kablach obwodów elektrycznych. Można go raczej porównać ze zjawiskiem zachodzącym w zapalonym na jednym końcu loncie z tą różnicą, iż lont można użyć dla wyzwolenia reakcji (wybuchu) tylko jeden raz, natomiast włókno nerwowe może być użyte natychmiast ponownie po przejściu pojedynczego impulsu. Tak jak wzdłuż zapalonego lontu przesuwa się pewna reakcja chemiczna (spalanie się łatwopalnego materiału), podobnie pobudzenie i wędrówka impulsu wzdłuż nerwu polegają na szybkim przesuwaniu się w określonym kierunku pewnej reakcji fizykochemicznej, wyrażającej się zmianą różnicy potencjału pomiędzy wnętrzem i zewnętrzną powierzchnią błony pokrywającej włókno nerwowe. W stanie spoczynku owa różnica potencjałów wynosi ok. minus 70 miliwoltów. Jest ona wynikiem różnego stężenia jonów sodu i potasu po obu stronach wspomnianej błony (ryc. 193). Mówimy, że błona komórkowa jest spolaryzowana. Powstanie w pewnym punkcie bodźca nerwowego polega na zmniejszeniu owej różnicy potencjałów, czyli depolaryzacji nerwu w danym punkcie poniżej 70 miliwoltów. Następuje przy tym nagła zmiana przepuszczalności błony komórkowej, pozwalająca jonom sodowym wnikać z zewnątrz przez błonę, zaś jonom potasowym wychodzić na zewnątrz błony. To przemieszczanie się jonów sodu i potasu przez błonę przesuwa się wzdłuż włókna nerwowego, powodując depolaryzację coraz to nowego punktu włókna. Ta nienormalna przepuszczalność błony dla jonów sodu i potasu jest zjawiskiem chwilowym, trwającym ułamek sekundy, po którym zostają w danym punkcie odtworzone warunki pierwotne (polaryzacja) i odcinek ten odzyskuje ponownie zdolność pobudzenia. 44ł44ł+łł-H- + +-l--t+4++++4 + +4+4
: 4- -H-*-H-+'-ł ++•!- + <—t- + -n-. -+- -i- —h -ł- -i- + + 4- 4-4- 4- 4- 4- +4- +4- 4-4-4-+ 4y ^ y J p ^ - p ^
Ryc. 193. Schemat wędrówki pobudzenia w nerwie (opis w tekście).
'409
Prędkość, z jaką opisany wyżej bodziec, przesuwa się wzdłuż włókna nerwowego w różnych aksonach jest różna i waha się od 4 do 400 km/h. Bardzo ważną cechą przewodzenia bodźców w aksonie jest to, że pobudzenie w czasie wędrówki wzdłuż aksonu nie zanika ani nie traci swej siły w odróżnieniu np. od sygnałów elektrycznych przesyłanych np. przez kable telefoniczne na duże odległości. Dzieje się tak dlatego, że przewodzenie bodźca jest związane ze zużyciem, minimalnej wprawdzie, energii pochodzącej z procesów chemicznych przemiany, doprowadzających do powstania różnicy stężeń sodu i potasu po obu stronach błony komórkowej. Skomplikowany układ połączeń olbrzymiej liczby neuronów w układzie nerwowym posługuje się zatem jednolitym sposobem przekazywania sygnałów. Rodzaj tych informacji (światło, zapach, smak, dźwięk itd.) oraz rozkazów zostaje odczytany zależnie od tego, jakie neurony czy grupy neuronów zostały pobudzone. Neurony muszą przekazywać jednak nie tylko pobudzenie, ale także jego siłę - potrafimy nie tylko odróżnić światło, dźwięk itp., lecz także ocenić jego natężenie. Natura rozwiązała ten problem w ten sposób, iż siła bodźca zostaje przełożona na częstotliwość impulsów nerwowych. Słabe bodźce wywołują impulsy o małej częstotliwości (kilka impulsów na sekundę), bodźce silne powodują powstawanie impulsów o dużej częstotliwości (kilkaset na sekundę). W ten sposób jeden i ten sam rodzaj impulsów, przekazywany z różną Konwergencja
Dywergencja
A Ryc. 194. Zbieżność i rozbieżność neuronów. A - Aksony trzech neuronów tworzą kontakt synaptyczny z jedną komórką nerwową. S- Jeden akson tworzy połączenia z komórkami trzech neuronów (wg Grollmana).
'410
częstotliwością, dostarcza informacji zarówno o rodzaju sygnału, jak i natężeniu. Zupełnie inny rodzaj zjawisk następuje, gdy bodziec wędrujący wzdłuż włókna nerwowego nadejdzie do miejsca zetknięcia się aksonu jednego neuronu z ciałem komórkowym lub dendrytem innego neuronu. Miejsce takie nazywa się synapsą. Schematyczną budowę synapsy ilustruje ryc. 194. Na rycinie tej widać, że akson neuronu przekazującego bodziec nie styka się z ciałem komórki odbierającej bodziec, lecz istnieje pomiędzy nimi mikroskopijna szczelina. Bodziec nerwowy pokonuje tę szczelinę za pomocą specjalnego przenośnika chemicznego. Każdy nadchodzący do synapsy bodziec elektryczny powoduje wydzielanie się minimalnej ilości pewnej substancji przekaźnikowej, która przepływa na drugą stronę szczeliny synaptycznej, zmieniając w podobny sposób redystrybucję jonów sodu i potasu po obu stronach błony komórkowej ciała neuronu i związaną z tym zmianę ładunku elektrycznego w protoplazmie ciała neuronu. Wiemy już, że taka chwilowa zmiana potencjału jest równoznaczna z impulsem nerwowym. W ciele komórkowym ów impuls rozchodzi się w ciele neuronu w ciągu około milisekundy. Istnieje jednak pewna istotna różnica pomiędzy rozchodzeniem się bodźca w aksonie a pobudzeniem wyzwalanym w synapsie. Pobudzenie aksonu ma charakter zjawiska określanego wfizjologii jako „wszystko albo n i c " - bodziec, jeżeli jest dostatecznie silny, wyzwala zawsze tę samą reakcję; jeżeli jest zbyt słaby, reakcji tej nie jest w stanie zapoczątkować. W synapsie natomiast każdy nadchodzący bodziec powoduje przejście na drugą stronę szczeliny synaptycznej, pewnej porcji substancji przekaźnikowej, która rozkłada się stosunkowo powoli. Rzecz w tym, że na ciele komórkowym jednego neuronu znajduje zakończenie zwykle większa liczba aksonów i rzadko neuron zostaje pobudzony przez bodziec nadchodzący z jednego tylko aksonu. Nadchodzące równocześnie lub w bardzo krótkich odstępach czasu, bodźce, z których każdy powoduje wyzwolenie się pewnej ilości substancji przekaźnikowej utrzymującej się w stanie czynnym ok. 5-10 ms, powoduje zsumowanie się tych pojedynczych efektów. W ten sposób neuron odbiorczy staje się swego rodzaju „wzmacniaczem sumującym" różnych nadchodzących sygnałów, wywołując reakcję wzmocnioną w postaci serii bodźców o zwiększonej częstotliwości, przekazywanej do swego aksonu. Na schematach ilustrujących ten proces przedstawia się zwykle tylko łańcuch złożony co najwyżej z kilku neuronów, w rzeczywistości jednak bodźce czuciowe płyną bezustannie do neuronów pośrednich przez wielką liczbę przyległych włókien nerwowych. Natura zastosowała w układzie nerwowym system przewodzenia przez liczne równoległe kanały, dzięki czemu uszkodzenie kilku komórek nerwowych nie odbija się w istotniejszym stopniu na wykonywanej przez nie czynności, ponieważ mogą one być przejęte przez kanały równoległe. Mówiliśmy dotychczas o pobudzeniu neuronu przez nadchodzący bodziec. Obok tego istnieją w układzie nerwowym neurony, które pod wpływem '411
nadchodzących do nich bodźców zostają jak gdyby „pobudzone negatywnie", tj. reaktywność ich nie zwiększa się, lecz przeciwnie- zmniejsza; mówimy, że następuje w nich hamowanie. Wyobrażamy sobie ów proces w t e n sposób,że nadchodzący do takiego neuronu bodziec wyzwala substancję chemiczną, która nie ułatwia, lecz odwrotnie - utrudnia przechodzenie jonów sodu i potasu przez błonę komórkową. Typowy neuron pośredni otrzymuje bodźce zarówno od licznych neuronów pobudzających, jak i hamujących. Jego aktualny stan - pobudzenie lub hamowanie - zależy od wypadkowej wszystkich nadchodzących bodźców pobudzających i hamujących. Dzięki powyższym właściwościom przewodzenia synaptycznego neuron wykazuje w pewnych okolicznościach właściwości urządzenia o zmiennym progu pobudliwości. Jeżeli szereg słabych sygnałów pobudzających zsumuje się do wartości zbliżonych do progu wyładowania, to wówczas bardzo słaby dodatkowy impuls może wystarczyć dla wywołania powstania potencjału czynnościowego neuronu i odwrotnie - obecność bodźców hamujących z niektórych zakończeń może spowodować, że neuron staje się niewrażliwy na inne sygnały pobudzające, które normalnie wystarczyłyby dla pobudzenia. Widzimy zatem, iż neuron ma zdolność dodawania i odejmowania różnych bodźców przez podwyższanie lub obniżanie progu pobudliwości.
RDZEŃ KRĘGOWY Rdzeń kręgowy (medulla spinalis) ma kształt grubego sznura, nieco spłaszczonego w kierunku strzałkowym, o przeciętnej średnicy 1 cm, barwy białej o ciężarze ok. 30 g. Umieszczony jest w kanale kręgowym. Rozpoczyna się on bez wyraźnej granicy od rdzenia przedłużonego i rozciąga na przestrzeni około 45 cm, od I kręgu szyjnego do górnej krawędzi II kręgu lędźwiowego, gdzie kończy się stożkiem rdzeniowym. Rdzeń nie wypełnia więc kanału rdzeniowego całkowicie, lecz jest od niego krótszy. Od rdzenia odchodzą parzyste nerwy rdzeniowe (nervi spinales). Poszczególne otwory międzykręgowe, przez które wychodzą odpowiednie nerwy rdzeniowe, są przesunięte względem rdzenia ku dołowi. Wskutek tego nerwy wychodzące z dolnych odcinków rdzenia, zanim osiągną właściwe otwory międzykręgowe, biegną na znacznej przestrzeni wewnątrz kanału kręgowego. Zespół tych nerwów, otaczających nić końcową, nosi nazwę ogona końskiego (cauda equina). W zależności od okolicy rdzenia, z której odchodzą nerwy, rozróżniamy w nim części: szyjną, piersiową, lędźwiową i krzyżowo-guziczną. Średnica rdzenia nie jest równomierna na całej jego długości. W odcinkach, z których wychodzą korzenie nerwów przeznaczonych dla kończyn, rdzeń ma dwa wrzecionowate zgrubienia: zgrubienie szyjne - na przestrzeni od II kręgu szyjnego do II kręgu piersiowego, oraz zgrubienie lędźwiowe, ciągnące się od X do XII kręgu piersiowego. Na powierzchni zewnętrznej rdzenia widnieją w linii przyśrodkowej dwie bruzdy: głębsza od przodu, zwana szczeliną pośrodkową przednią, oraz '412
Ryc. 195. Rdzeń kręgowy (1:1): A - w i d o k od przodu, B- widok z boku.
płytsza od tyłu - bruzda pośrodkowa tylna. Dzielą one rdzeń na dwie symetryczne połowy: prawą i lewą. Na powierzchni każdej połowy rdzenia zaznacza się: od przodu podłużna linia, zwana bruzdą boczną przednią, z któret wychodzą korzenie ruchowe nerwów rdzeniowych, oraz od tyłu bruzda boczna tylna, wzdłuz której wchodzą do rdzenia korzenie czuciowe nerwów rdzeniowych. Wymienione bruzdy '413
dzielą każdą połowę rdzenia na trzy sznury: przedni, boczny i tylny. Są one utworzone przez pęczki włókien nerwowych, przebiegające podłużnie w rdzeniu. Korzenie brzuszne (radices ventrales) składają się z włókien ruchowych, zdążających do mięśni szkieletowych. W skład korzeni grzbietowych (radices dorsales) wchodzą włókna czuciowe, idące od komórek położonych w odpowiednich zwojach rdzeniowych (ganglia spinalia). Korzenie brzuszne i grzbietowe łączą się ze sobą bocznie od zwoju rdzeniowego, tworząc mieszany nerw rdzeniowy (nervus spinalis). U człowieka wychodzi z rdzenia 31 par nerwów: 8 szyjnych, 12 piersiowych, 5 lędźwiowych, 5 krzyżowych i 1 para nerwów guzicznych. Każdej parze nerwów odpowiada odcinek, zwany segmentem rdzeniowym. Poszczególne segmenty oraz nerwy rdzeniowe oznacza się pierwszą literą łacińskiej nazwy odcinka z podaniem kolejnej liczby segmentu, np. Ct (cervicalis primus) pierwszy szyjny, Jh3 (thoracalis tertius-trzeci piersiowy), l_5 (lumbalisquintus - piąty lędźwiowy), S3 (sacralis tertius - trzeci krzyżowy), C0 (coccygeus guziczny). Budowa wewnętrzna rdzenia nie jest jednorodna. Na przekroju poprzecznym ukazuje się ciemniejsza część pośrodkowa w kształcie motyla lub litery H, zwana istotą szarą (substantia grisea), oraz otaczająca ją na kształt płaszcza istota biała (substantia alba). Zasadniczym składnikiem istoty szarej są ciała komórek nerwowych; istota biała stanowi skupienie opisanych już wyżej pęczków włókien nerwowych, połączonych w sznury. W istocie szarej rozróżniamy słupy przednie i tylne, które na przekroju mają kształt rogów przednich i tylnych, oraz istotę szarą środkową, skupioną wokół ciągnącego się wzdłuż całego rdzenia kanału środkowego. Kanał ten łączy się u góry z komorami, leżącymi wewnątrz mózgowia. W odcinku piersiowym pomiędzy słupem przednim a tylnym istoty szarej występuje jeszcze słup boczny. W obrębie słupów przednich istoty szarej znajdują się duże komórki jąder ruchowych, których neuryty wychodzą z rdzenia w składzie korzeni brzusznych. W zrębie słupów bocznych odcinka piersiowego występuje znacznie mniejsze skupienie komórek stanowiących jądro współczulne rdzeniowe. U podstawy słupów tylnych leży zespół komórek, tworzących tzw. jądro grzbietowe. Neurony wchodzące w skład tego jądra stanowią człon, włączony pomiędzy neurony czuciowe a ruchowe rdzeniowego łuku odruchowego. Istota biała utworzona jest z włókien nerwowych, skupionych w rdzeniu w tzw. sznury. Istota biała jest najlepiej rozwinięta w odcinku szyjnym i piersiowym. W skład jej wchodzą włókna wstępujące, tj. takie, które przewodzą bodźce w kierunku mózgu, i włókna zstępujące, przewodzące bodźce w kierunku odwrotnym. Zespoły włókien przewodzące identyczne podniety w tym samym kierunku noszą nazwę dróg nerwowych. Odróżniamy wśród nich drogi zstępujące, czyli odmózgowe, i wstępujące, czyli domózgowe. Pierwsze przebiegają w sznurach przednich rdzenia, drugie - w sznurach tylnych; sznury boczne zawierają drogi obu rodzajów. '414
Ośrodki rdzenia kręgowego Ośrodki rdzeniowe są rozmieszczone odcinkowo, tzn. pewne odcinki rdzenia zaopatrują określone odcinki (metamery) ciała w nerwy czuciowe, ruchowe, wydziel nicze i naczynioruchowe. Do najważniejszych ośrodków rdzenia należą ośrodki ruchowe kończyn i tułowia. Rozmieszczenie ich w rdzeniu przedstawia się następująco: a) ośrodek ruchów przepony, tj. zespół komórek, od których odchodzą włókna nerwu przeponowego, leży na wysokości lll-IV segmentu szyjnego; b) ośrodki ruchowe kończyn górnych mieszczą się w V—VI11 segmentach szyjnych i pierwszym segmencie piersiowym; c) w części piersiowej rdzenia kręgowego leżą ośrodki mięśni klatki piersiowej, grzbietu i brzucha; d) w zgrubieniu lędźwiowym leżą ośrodki ruchowe kończyn dolnych. Poza tym w rdzeniu znajdują się ośrodki autonomiczne. W ostatnim odcinku szyjnym i dwóch górnych piersiowych znajduje się ośrodek odruchów źrenicy. Wskutek podrażnienia tego ośrodka występuje odruchowe rozszerzenie źrenicy i wysunięcie do przodu gałki ocznej. W odcinku krzyżowym rdzenia mieszczą się ośrodki oddawania moczu, defekacji i ejakulacji. We wszystkich odcinkach piersiowych i górnych lędźwiowych rozmieszczone są ośrodki naczynioruchowe i wydzielnicze dla gruczołów potowych skóry.
Odruch Czynności układu nerwowego, podobnie jak i jego budowa, są niezwykle złożone. Mimo to jednak można wyodrębnić pewien elementarny akt czynności układu nerwowego, który pozwala ustalić jedną wspólną zasadę czynności tego układu dla wszystkich przedstawicieli świata zwierzęcego, a w tej liczbie i człowieka. Tym zasadniczym aktem jest odruch (ref/ex). Odruchem nazywamy automatyczną reakcję na bodziec zewnętrzny lub wewnętrzny, zachodzącą przy udziale ośrodkowego układu nerwowego. Każdy odruch rozpoczyna się pod wpływem bodźca działającego na zakończenia nerwowe, które znajdują się w narządach nerwowych, zwanych receptorami, i kończy się jakąś zmianą czynności ustroju. Wszystkie receptory można podzielić na trzy grupy: 1) eksteroreceptory, czyli narządy zmysłów, umieszczone na zewnętrznej powierzchni ciała; 2) proprioreceptory, tj. zakończenia nerwowe w narządach ruchu - mięśniach, ścięgnach i stawach; 3) wisceroreceptory, tj. zakończenia nerwowe w narządach wewnętrznych. Zależnie od tego, w której z wyżej wymienionych grup receptorów odruch się rozpoczyna, rozróżniamy odruchy: eksteroceptywne, proprioceptywne i wisceroceptywne. Znamy bardzo dużo najrozmaitszych odruchów, np. przy mechanicznym podrażnieniu podeszwowej powierzchni skóry stopy występuje odruchowe '415
zgięcie stopy - odruch podeszwowy; dotknięcie okolicy ust noworodka wywołuje odruch ssania; dotknięcie jakimś twardym przedmiotem tylnej ściany gardła sprowadza odruch wymiotny; uderzenie w ścięgno mięśnia czworogłowego wywołuje nagłe wyprostowanie w stawie kolanowym; uderzenie w ścięgno Achillesa - odruchowy skurcz mięśnia trójgłowego łydki; nagłe podrażnienie narządów jamy brzusznej wywołuje odruchowe zahamowanie czynności serca (odruch Goltza) i in.
i i %
\
/
Ropi przednie
M dwugłowy— ramienia
_
Staw łokciowy
Kość łokciowa
Odruchowy skurcz
rięśnia
Rozciąganie mięśnia
J
Ryc. 196. Schemat ilustrujący odruch rozciągania.
Wszystkie odruchy odbywają się przy udziale ośrodkowego układu nerwowego. Łatwo się o tym przekonać, wykonując doświadczenia na żabie z wyciętym mózgiem i zachowanym rdzeniem kręgowym. U żaby takiej można wywołać szereg odruchów. Jeżeli np. zanurzymy jej łapkę w słabym roztworze kwasu, następuje odruch zginania, łapka zostaje wyciągnięta z kwasu; jeżeli do skóry bocznej powierzchni tułowia przyłożymy kawałeczek bibuły zwilżonej kwasem, wystąpi odruch pocierania, polegający na skurczach odpowiedniej '416
łapki i pocieraniu jej o tułów. Wystarczy jednak zniszczyć rdzeń kręgowy, np. przez włożenie do kanału kręgowego sondy, a wszystkie odruchy znikają. Każdy odruch rozpoczyna się od podrażnienia zakończeń nerwowych w jakimś receptorze. Powstające tu pobudzenie zostaje przekazane przez włókna nerwowe dośrodkowe do ośrodkowego układu nerwowego. Włókna dośrodkowe są wypustkami komórek nerwowych, skupionych na zewnątrz mózgu i rdzenia kręgowego, w zwojach rdzeniowych. W najprostszym przypadku pobudzenie zostaje przekazane neuronowi ośrodkowemu (ruchowemu), który je doprowadza do odpowiedniego narządu (mięśnia, gruczołu), wprawiając go w stan czynny. Na rycinie 196 przedstawiono schematycznie tzw. odruch rozciągania. Rozciąganie mięśnia połączonego nerwami z rdzeniem powoduje odruchowy jego skurcz. Znajdujące się w mięśniu proprioreceptory w postaci tzw. wrzecion mięśniowych reagują na bodziec rozciągania w ten sposób, że wysyłają impulsy do ośrodka ruchowego w rdzeniu, skąd neuron ruchowy wysyła impulsy przez akson ruchowy do mięśnia. Dla wywołania tego odruchu wystarcza wydłużenie wrzeciona o ułamek centymetra. Odruch ten ma olbrzymie znaczenie dla utrzymania postawy ciała. Drogę, którą wędruje pobudzenie w czasie odruchu, nazywamy łukiem odruchowym. Opisany wyżej łuk odruchowy składa się z dwóch neuronów. Większość fizjologów sądzi jednak, że łuk odruchowy przeważnie musi się składać co najmniej z trzech neuronów; pomiędzy neuron czuciowy a ruchowy zostaje włączony trzeci neuron pośredni (kontaktowy), którego wypustki nie wychodzą poza ośrodkowy układ nerwowy, lecz rozprowadzają pobudzenie w granicach tego układu. Wskutek tego pobudzenie rozszerza się i obejmuje znaczne obszary ośrodkowego układu nerwowego. Nazywamy to promieniowaniem (irradiacją) pobudzenia. Jeżeli w opisanym wyżej doświadczeniu z odmóżdżoną żabą wywołamy odruch pocierania, to ruchy pocierania wykonuje zawsze łapka tej strony, na której przyłożyliśmy bibułę z kwasem. Jeżeli doświadczenie zmienimy w ten sposób, że łapkę tę amputujemy, to po jakiś czasie wystąpi odruch pocierania po stronie przeciwnej. Wytłumaczyć to zjawisko można tylko w ten sposób, że pobudzenie rozeszło się w rdzeniu żaby i objęło ośrodki ruchowe, zarządzające ruchami strony przeciwnej. Im silniejszy jest bodziec i im dłuższy czas jego działania, tym większe obszary ośrodkowego układu nerwowego obejmuje pobudzenie promieniujące z jednego punktu.
Opony rdzenia kręgowego Rdzeń otaczają trzy opony (meriingeś). Wymieniane od zewnątrz są one następujące: opona twarda (dura mater spinalis), pajęczynówka (arachnoidea spinalis) i opona miękka rdzenia (pia mater spinalis). Mocna i gruba opona twarda, będąca przedłużeniem opony twardej mózgowia, ma kształt długiego walcowatego worka, dosyć luźno obejmującego rdzeń, zakończonego ślepo u dołu na wysokości II-III kręgu krzyżowego. 27 - Anatomia i fizjologia
417
Worek ten składa się z dwóch blaszek, przy czym blaszka zewnętrzna stanowi okostną kanału kręgowego. Szczelinowata, lecz dosyć pojemna, przestrzeń zawarta między tymi blaszkami, wypełniona przejrzystym płynem tkankow y m , tkanką tłuszczową i splotami żylnymi, nazywa się jamą nadtwardówkową. Wąska szczelina pomiędzy blaszką wewnętrzną opony twardej a pajęczynówką nazywa się jamą podtwardówkową. Blaszka wewnętrzna opony twardej, umocowana jest w otworach międzykręgowych, otaczając na kształt rękawa wychodzące przez nie korzenie nerwowe. W obrębie otworów międzykręgowych cienka blaszka pajęczynówki łączy się z oponą twardą. Przestrzeń zawarta między pajęczynówką a oponą miękką nazywa się jamą podpajęczynówkową. Wypełnia ją płyn mózgowo-rdzeniowy (liquor cerebrospinalis). Jama podpajęczynówkowa rdzenia łączy się z jamą podpajęczynówkową mózgowia, co ma duże znaczenie dla krążenia płynu mózgowo-rdzeniowego. Rozszerzająca się u dołu część jamy podpajęczynówkowej rdzenia, w której znajduje się tylko ogon koński, nazywa się zbiornikiem końcowym. Ze zbiornika tego można wydobyć płyn mózgowo-rdzeniowy drogą nakłucia lędźwiowego, bez obawy uszkodzenia rdzenia. Opona miękka, która przylega ściśle do rdzenia, zawiera liczne rozgałęzienia naczyń krwionośnych, które wnikają w jego miąższ. Od pokrywającej rdzeń opony miękkiej odchodzą w linii bocznej z każdej strony poprzecznie ustawione wyrostki cienkiej, lecz mocnej, blaszki w kształcie ząbków, zwężających się w kierunku bocznym. Ząbki te, w liczbie 23 z każdej strony, tworzą więzadło ząbkowane (ligamentum denticulatum). Wierzchołki tych ząbków zrastają się z pajęczynówką, ustalając w ten sposób rdzeń. Widzimy więc, że rdzeń kręgowy, zawieszony na więzadle ząbkowanym i korzonkach rdzeniowych, pływa równocześnie w płynie mózgowo-rdzeniow y m , co razem z poduszką z tkanki tłuszczowej i splotów żylnych, wypełniających jamę nadtwardówkową, chroni go od wstrząsów i urazów mechanicznych.
OPONY MÓZGOWIA Mózgowie otaczają trzy opony (meninges encephali): zewnętrzna - opona twarda (dura matei), środkowa - pajęczynówka (arachnoidea) i wewnętrzna, przylegająca bezpośrednio do tkanki mózgowej - opona miękka (p/a mater).
Opona twarda mózgowia Opona twarda (dura mater) jest błoną włóknistą, która równocześnie stanowi okostną jamę czaszki. U osobników dorosłych opona twarda jest ściśle zrośnięta z kośćmi podstawy czaszki, natomiast z kośćmi sklepienia czaszki jest połączona dosyć luźno i łatwo się od nich oddziela. Przez otwór '418
wielki kości potylicznej opona mózgowia przechodzi do kanału kręgowego, przedłużając się w oponę twardą rdzenia. Gładka i błyszcząca powierzchnia wewnętrzna opony twardej oddzielona jest od pajęczynówki szczelinowatą przestrzenią, zwaną jamą podtwardówkową (cavum subdurale). Opona oddaje wewnątrz czaszki trzy płaszczyznowe wyrostki: sierp mózgu, sierp móżdżku i namiot móżdżku. 1. Sierp mózgu (fa/x cerebri), ustawiony w płaszczyźnie pośrodkowej, wchodzi do szczeliny podłużnej mózgu. Górny brzeg sierpa mózgu przyczepia się do wewnętrznej powierzchni kości czaszki wzdłuż bruzdy zatoki strzałkowej, biegnąc od grzebienia koguciegb do guzowatości wewnętrznej kości potylicznej. Wolny brzeg dolny nie dochodzi do ciała modzelowatego. 2. Sierp móżdżku (fa/x cerebelli) stanowi przedłużenie sierpa mózgu od guzowatości potylicznej do otworu wielkiego kości potylicznej. Wolny brzeg sierpa móżdżku wchodzi pomiędzy półkule móżdżkowe. 3. Namiot móżdżku (tentorium cerebelli) jest ustawiony prawie poprzecznie, tworząc jak gdyby dach ponad dołem czaszki tylnym. Namiot móżdżku oddziela półkule móżdżku od płatów potylicznych półkul mózgowych.
Pajęczynówka mózgowia Pajęczynówka (arachnoidea encephali) jest cienką błoną łącznotkankową, którą oddziela od opony twardej włosowata jama podtwardówkowa. Z położoną pod nią oponą miękką zrasta się pajęczynówka tylko na wypukłych powierzchniach zakrętów mózgowych. Nad bruzdami i szczelinami opona ta przerzuca się, nie zagłębiając się w nie, wskutek czego pomiędzy oponą miękką a pajęczynówką powstaje szereg większych i mniejszych przestrzeni, które się ze sobą komunikują i w całości noszą nazwę jamy podpajęczynówkow e j (cavum subarachnoideale). Większe przestrzenie podpajęczynówkowe noszą nazwę zbiorników (cisternae subarachnoideales). Jama podpajęczynówkowa jest wypełniona płynem mózgowo-rdzeniow y m , który wydostaje się z komory czwartej do zbiornika, otaczającego rdzeń przedłużony i móżdżek (zbiornik móżdżkowo-rdzeniowy), przez dwa parzyste otwory boczne i nieparzysty otwór pośrodkowy komory czwartej.
Opona miękka mózgowia Opona miękka (p/a mater encephali) jest zrośnięta z powierzchnią zewnętrzną mózgu i wchodzi we wszystkie zagłębienia, bruzdy i szczeliny. Zawiera ona liczne naczynia krwionośne tętnicze i żylne, które wnikają do tkanki mózgowej.
'419
Płyn mózgowo-rdzeniowy Płynem mózgowo-rdzeniowym (liquor cerebrospinaliś) nazywamy ciecz, wypełniającą komory mózgu oraz przestrzenie podpajęczynówkowe mózgowia i rdzenia kręgowego. Płyn mózgowo-rdzeniowy powstaje w splotach naczyniówkowych w wyniku przesączania się wody i pewnych składników osocza krwi. Objętość płynu mózgowo-rdzeniowego wynosi u człowieka dorosłego od 100 do 250 ml. Płyn jest przejrzysty, bezbarwny, o ciężarze właściwym 1,006. Skład płynu mózgowo-rdzeniowego przedstawia tab. 30. Ciśnienie płynu mózgowo-rdzeniowego wynosi 150 mm słupa wody. Z komór mózgowych płyn mózgowo-rdzeniowy wydostaje się do jamy podpajęczynówkowej przez otwory w komorze czwartej. Głównym miejscem wchłaniania się płynu z jamy podpajęczynówkowej są ziarnistości pajęczynówki (granulationes arachnoideales), wpuklające się do zatoki strzałkowej górnej i jej uwypukleń bocznych. Obecność płynu mózgowo-rdzeniowego przyczynia się do równomiernego rozkładu ciśnienia w jamie czaszki. Możemy powiedzieć, że mózgowie jest zanurzone w płynie mózgowo-rdzeniowym, co też chroni je przed urazami. T a b e l a 30 Składniki
mg%
Białko Glukoza Wapń Potas Chlorek sodowy Fosfor nieorganiczny Wodorowęglany (% obj.)
15-55 50-80 4- 6 12-17 720-750 1,25-2,0 40-60
MÓZGOWIE Mózgowiem (encephalon) nazywamy część ośrodkowego układu nerwowego, umieszczoną w jamie czaszki. W skład mózgowia wchodzi mózg (cerebrum), móżdżek (cerebellum) i rdzeń przedłużony (medulla oblongata). Dla celów fizjologicznych i klinicznych dzielimy mózgowie na mózg (cerebrum) i pień mózgu (truncus cerebri). Mózgiem nazywamy półkule mózgowe i część wzrokową podwzgórza, pniem - pozostałą część mózgowia. Przeciętny ciężar mózgowia wynosi 1375 g u mężczyzn i 1240 g u kobiet. Mózgowie noworodków męskich waży 340 g, żeńskich - 330. Ciężar i wielkość mózgowia podlegają dużym wahaniom osobniczym. Między ciężarem mózgowia a uzdolnieniem jego posiadacza nie ma współzależności. Zestawienie ciężaru mózgowia człowieka z ciężarem mózgowia przedstawicieli rozmaitych gatunków ssaków wykazuje, że tylko u niewielu najpotężniejszych ssaków, takich jak słoń i wieloryb, ciężar bezwzględny mózgowia jest '420
Bruzda środkowa
Bruzda
boczna mózgu
Móżdżek
Rdzeń przedłużony Ryc. 197. Mózgowie - widok od strony prawej.
większy niż u człowieka. Pod względem ciężaru mózgowia człowiek przewyższa znacznie swych najbliższych krewnych z gromady ssaków, inne Naczelne. Duży ciężar bezwzględny i względny mózgowia człowieka jest wyrazem daleko posuniętej centralizacji ośrodkowego układu nerwowego. Mózg ma kształt jajowaty. Jego dolna powierzchnia jest spłaszczona i nosi nazwę podstawy mózgu (basis cerebri). Powierzchnie boczna i górna są wypukłe. W linii pośrodkowej na powierzchni przedniej, górnej i tylnej jest widoczna głęboka szczelina podłużna mózgu, oddzielająca duże półkule mózgu (hemispheria cerebri) - prawą i lewą. Półkule mózgu zakrywają całkowicie od góry pień mózgu. W tyle oddziela je od móżdżku głęboka szczelina poprzeczna mózgu. Powierzchnię półkul mózgu pokrywa cienka warstwa istoty szarej, zwana korą mózgową (cortex cerebri), stanowiąca skupienie komórek nerwowych. Pod korą mózgową znajduje się w półkulach mózgu istota biała (substantia alba), która składa się ze ściśle do siebie przylegających włókien nerwowych, przebiegających głównie między komórkami kory mózgowej. Skupienia komórek nerwowych w obrębie istoty białej noszą nazwę jąder podkorowych. Powierzchnia półkul mózgu wykazuje liczne fałdy, zwane zakrętami mózgu (gyri cerebri), które są porozdzielane bruzdami (sulci) lub szczelinami (f/ssurae) mózgu. '421
Szczelina podłużna mózgu
Ryc. 198. Powierzchnia podstawna mózgu (po usunięciu móżdżku).
Na dnie szczeliny podłużnej mózgu znajduje się spoidło wielkie mózgu, zwane także ciałem m o d z e l o w a t y m ( c o r p u s callosum), składające się z włókien łączących obie półkule. Podstawa mózgowia jest utworzona przez dolne powierzchnie półkul mózgu oraz dolną powierzchnię pnia mózgowego. Posuwając się od tyłu ku przodowi na podstawie mózgowia znajdujemy opisane niżej twory. 1. Rdzeń przedłużony ( m e d u l l a oblongata) w linii pośrodkowej jest rozdzielony szczeliną pośrodkową przednią na dwie podłużne, wałkowate wyniosłości, zwane piramidami ( p y r a m i d e s ) . Bocznie od każdej z piramid leży owalny t w ó r , zwany oliwką (oliva). 2. M o s t ( pons) przylega od przodu do rdzenia przedłużonego jako lity w a ł poprzeczny, zbudowany głównie z włókien nerwowych. 3. Konary mózgu (pedunculi cerebri) wychodzą spod przedniej krawędzi mostu i biegną ku przodowi, bokowi i ku górze, by się zagłębić w półkulach '422
mózgowych. Ograniczają one trójkątne zagłębienie, zwane dołem międzykonarowym, na dnie którego leży istota dziurkowana tylna. Przebijają ją liczne, wnikające tędy do mózgu naczynia krwionośne, po których oderwaniu pozostają drobne otworki. 4. Ciała suteczkowate (corpora mamillaria) - białe wzgórki położone ku przodowi od istoty dziurkowanej tylnej. 5. Guz popielaty (tuber cinereum) jest wypukleniem przylegającym z tyłu do ciał suteczkowatych. Ku dołowi guz popielaty przechodzi w zwężający się lejek ( i n f u n d i b u l u m ) , na którym niejako zawieszona jest przysadka ( h y p o p h y sis cerebri), umieszczona w dole przysadki na górnej powierzchni trzonu kości klinowej. 6. Skrzyżowanie w z r o k o w e ( chiasma opticum) przylega do guza popielatego od przodu; w bok i ku tyłowi z każdej strony odchodzi od skrzyżowania wzrokowego pasmo w z r o k o w e (tractus opticuś), ku przodowi zaś - n e r w w z r o k o w y (nervus opticuś). 7. Nad skrzyżowaniem wzrokowym znajduje się cienka blaszka istoty szarej, zwana blaszką krańcową mózgu ( l a m i n a terminalis). 8. Bocznie i ku przodowi od skrzyżowania wzrokowego znajduje się istota dziurkowana przednia ( substantia perforata anterior). 9. Na powierzchni dolnej płatów czołowych widnieje po każdej stronie podłużne pasmo w ę c h o w e ( tractus olfactorius), biegnące prawie równolegle do szczeliny podłużnej mózgu. Pasmo węchowe rozpoczyna się na przodzie zgrubieniem, zwanym opuszką węchową (bulbus olfactorius) i kończy się w tyle t r ó j k ą t e m w ę c h o w y m . Ciało modzelowate
Rdzeń przedłużony Ryc. 199. Powierzchnia przyśrodkową mózgu.
'423
Rdzeń przedłużony Rdzeń przedłużony (meduila oblongata) łączy się z rdzeniem kręgowym bez wyraźnej granicy. Umowną granicą anatomiczną jest tu płaszczyzna, przechodząca przez brzegi otworu wielkiego kości potylicznej. Przez rdzeń przedłużony przebiegają wszystkie impulsy, idące z mózgu do rdzenia kręgowego i odwrotnie - z rdzenia do mózgu. U góry rdzeń przedłużony ma wyraźną granicę; po stronie przedniej tworzy ją tylny brzeg mostu. Na dolnej (przedniej) powierzchni rdzenia przedłużonego widnieją dwa wypukłe białe pasma, zwane piramidami (pyramides), które są utworzone przez włókna drogi piramidowej ( tractus pyramidalis), łączące komórki nerwowe kory mózgowej z komórkami rogów przednich rdzenia kręgowego. Większa część włókien drogi piramidowej przechodzi na stronę przeciwną, t w o rząc skrzyżowanie piramid, widoczne na dnie szczeliny pośrodkowej przedniej. Sznurom bocznym rdzenia kręgowego odpowiadają takie same sznury rdzenia przedłużonego, kończące się zgrubieniem, zwanym oliwką. Przebiegają tędy włókna drogi móżdżkowo-rdzeniowej grzbietowej. Kanał środkowy rdzenia kręgowego po przejściu do rdzenia przedłużonego stopniowo się rozszerza i wychodzi na tylną jego powierzchnię. Dno tego rozszerzonego kanału ma kształt rombu i nosi nazwę dołu równoległobocznego (fossa rhomboidea). Od tyłu dół równoległoboczny jest pokryty tylko oponą miękką, ponad którą leży móżdżek. Opisane wyżej rozszerzenie kanału środkowego, którego dno stanowi dół równoległoboczny, nosi nazwę k o m o r y czwartej (ventriculus ąuartus). Ku górze komora czwarta zwęża się ponownie i przechodzi w wąski przewód, zwany w o d o c i ą g i e m mózgu, który łączy się z p o n o w n y m rozszerzeniem - komorą trzecią, położoną już w międzymózgowiu. Komora czwarta połączona jestzjamą podpajęczynówkową trzema otworam i ; jeden nieparzysty otwór pośrodkowy komory czwartej leży w kącie d o l n y m dołu równoległobocznego, dwa parzyste otwory boczne znajdują się w kątach bocznych tego dołu. W rdzeniu przedłużonym, podobnie jak w rdzeniu kręgowym, występują skupienia istoty szarej, z których wychodzą na o b w ó d nerwy; są to jądra n e r w ó w czaszkowych. Rozmieszczenie tych jąder i wychodzących z nich n e r w ó w nie ma jednak tej prawidłowości odcinkowej, jaka charakteryzuje rdzeń kręgowy. Spośród 12 par nerwów czaszkowych 8 odchodzi od rdzenia przedłużonego. Są w ś r ó d nich nerwy czuciowe (dośrodkowe), ruchowe (odśrodkowe) i mieszane. Rdzeń przedłużony stanowi skupienie ośrodków nerwowych dla szeregu ważnych czynności odruchowych, które są związane z odchodzącymi od niego w y m i e n i o n y m i wyżej nerwami czaszkowymi. Do od ruchów odbywających się przy udziale rdzenia przedłużonego należą takie czynności, jak ssanie, żucie, '424
połykanie, w y m i o t y , kichanie, kaszel, mruganie powiek, ruchy oddechowe, regulacja czynności serca, rozszerzanie naczyń krwionośnych, wydzielanie potu, a także regulowanie przemiany materii. Odruchy te są wrodzone, tzn. są przekazywane dziedzicznie i występują już u noworodka. Liczne ośrodki rdzenia przedłużonego, jak np. ośrodek oddechowy, są czynne przez całe życie bez przerwy. Niektóre odznaczają się automatyzmem, tj. zostają pobudzane przez zmiany zachodzące w komórkach nerwowych, inne znów zostają pobudzone przez bodźce nadchodzące przez odpowiednie nerwy czuciowe. Czynność oddechową reguluje ośrodek oddechowy, położony w dolnej części dołu równoległobocznego. Uszkodzenie tego ośrodka prowadzi do ustania ruchów oddechowych i śmierci z uduszenia. Ośrodek ten jest pobudzany automatycznie przez rozpuszczony we krwi dwutlenek węgla i odruchowo przez bodźce nadchodzące z płuc za pośrednictwem włókien dośrodkow y c h nerwu błędnego. Ośrodek regulacji czynności serca pracuje zarówno automatycznie, jak też odruchowo. Czynność tego ośrodka przejawia się hamowaniem akcji serca. Pobudzenie automatyczne spowodowane jest zmianą składu krwi. Pobudzenie odruchowe następuje w różnych stanach, w których zostaje pobudzony ośrodek nerwu błędnego. Zwolnienie czynności serca pojawia się np. przy zwiększeniu ciśnienia w łuku aorty, przy ucisku na gałki oczne (odruch oczno-sercowy Aschnera), w tzw. odruchu Goltza, w y w o ł a n y m u żaby uderzeniem w trzewa przez przednią ścianę jamy brzusznej. Ośrodek naczynioruchowy jest pobudzany odruchowo. Pobudzenie jego rozszerza naczynia krwionośne. W rdzeniu przedłużonym znajdują się również ośrodki regulujące przemianę materii. Nakłucie pewnych okolic rdzenia przedłużonego w y w o ł u j e np. przechodzenie cuk»"j do moczu (cukromocz), zaburzenia przemiany wodnej i mineralnej. Za pośrednictwem rdzenia przedłużonego dochodzi również do skutku szereg odruchów orientujących w przestrzeni. Bodźce dośrodkowe w tych odruchach dochodzą przez nerw VIII oraz nerwy czuciowe dążące do mięśni. Powyższy przegląd wykazuje, że rdzeń przedłużony stanowi nadbudowę nad rdzeniem kręgowym - niejako wyższe piętro ośrodkowego układu nerwow e g o , które podporządkowało sobie prymitywniejsze samodzielne czynności rdzenia kręgowego.
Tyłomózgowie wtórne W skład t y ł o m ó z g o w i a w t ó r n e g o ( metencephalon) wchodzi m o s t (pons) i móżdżek ( c e r e b e l l u m ) . Most jest wyraźnie odgraniczony od sąsiednich części mózgowia tylko po stronie podstawnej - ma on kształt grubego walca ułożonego poprzecznie. '425
Część grzbietowa mostu wchodzi w skład dołu równoległobocznego, tworząc jego górną połowę. Most składa się z włókien nerwowych i skupień istoty szarej, tworzących jądra własne mostu. Przez most przebiegają włókna nerwowe, łączące korę mózgową z rdzeniem przedłużonym i kręgowym oraz z móżdżkiem. Móżdżek ( c e r e b e l l u m ) leży nad komorą czwartą i pokrywa dół równoległo: boczny. Od góry móżdżek zakryty jest płatami potylicznymi półkul mózgow y c h , od których oddziela go szczelina poprzeczna mózgu. Móżdżek składa się z części środkowej, zwanej robakiem {vermis) i z dwóch półkul ( h e m i s p h e r i a cerebelli). Kora móżdżku tworzy wąskie podłużne fałdy, porozdzielane licznymi, równolegle przebiegającymi bruzdami. Istota szara wnika dosyć daleko w głąb wspomnianych bruzd. Pod korą móżdżku znajduje się istota biała, utworzona z włókien nerwowych. W obrębie istoty białej leżą skupienia komórek nerwowych, zwane jądrami móżdżku. Największe z nich nazywa się jądrem zębatym ( nucleus dentatus). Czynności móżdżku pozostają w ścisłym związku z ruchami d o w o l n y m i . Móżdżek jest narządem, w którym odbywa się koordynacja, czyli uzgadnianie złożonych ruchów dowolnych. Zwterzę po usunięciu móżdżku: 1) doznaje zaburzeń w rozmieszczeniu napięcia mięśniowego, 2) nie potrafi skoordynować ruchów, 3) nie może utrzymać się w pozycji stojącej, 4) szybko się męczy. O roli móżdżku dają pojęcie zaburzenia występujące po jego usunięciu: 1. Atonia, czyli utrata napięcia mięśniowego, pojawia się bardzo szybko po wycięciu móżdżku i utrzymuje przez kilka dni. Następnie napięcie odzyskują mięśnie wyprostne, wskutek czego kończyny stają się sztywno wyprostowane, głowa odrzucona ku tyłowi. Po p e w n y m czasie zmiany napięcia mięśniow e g o ustępują, ruchy jednak pozostają niezgrabne. Uszkodzenie móżdżku pociąga za sobą zaburzenie rozdziału napięcia mięśniowego na poszczególne grupy mięśniowe. Podobne zmiany występują u człowieka przy chorobach móżdżku. 2. Ataksja - zaburzenie koordynacji ruchów - jest najjaskrawszym objaw e m uszkodzenia móżdżku. Zaburzenie to, określone jako niezborność lub bezład, polega na t y m , że zwierzę z wyciętym móżdżkiem nieprawidłowo ocenia zamierzone ruchy, tzn. siłę i zakres skurczu odpowiednich mięśni, wskutek czego ruchy są niezręczne, mają zasięg zbyt wielki lub zbyt mały. Człowiek z uszkodzonym móżdżkiem ma trudności w trafieniu palcem do nosa lub piętą do kolana drugiej kończyny. Chory taki np. zbyt wysoko unosi ramię, później zgina łokieć, przez co ruch traci właściwy kierunek. 3. Astazja polega na t y m , że zwierzę wykonuje ustawicznie drobne ruchy, które nazywamy drżeniem. Drżenie ustaje w w y g o d n y m położeniu, np. w pozycji leżącej, i nasila się przy zmianie postawy i każdym dodatkowym ruchu. Przy uszkodzeniach móżdżku występuje u ludzi tzw. drżenie za miarowe. Chory przy t y m dobrze trafia do celu, ale w chwili zbliżania np. kończyny do tego celu występuje u niego drżenie. '426
4. Astenia polega na szybkim męczeniu się zwierzęcia. Wynika to stąd, iż ruchy takiego zwierzęcia są nieekonomiczne i są wykonywane nawet w stanie spoczynku. Móżdżek jest więc narządem pomocniczym, który współdziała z korą mózgową w w y k o n y w a n i u ruchów dowolnych.
Śródmózgowie Ś r ó d m ó z g o w i e (mesencephalon) jest krótką częścią pnia mózgowego, łączącą międzymózgowie z mostem i móżdżkiem. W skład śródmózgowia wchodzą dwa konary m ó z g o w e ( pedunculi cerebri) i blaszka p o k r y w y śródm ó z g o w i a ( l a m i n a tecti mesencephali). Konary mózgowe znajdują się po stronie podstawnej - wchodzą one pod kątem ostrym z mostu i pogrążają się w półkulach mózgowych. Pomiędzy konarami znajduje się w s p o m n i a n y dół międzykonarowy (fossa interpeduncularis) i na jego dnie - istota dziurkowana tylna ( substantia perforata posterior). We wnętrzu śródmózgowia przebiega wąski kanał, z w a n y w o d o c i ą g i e m mózgu ( aąueductus cerebri), łączący komorę czwartą z komorą trzecią, położoną w międzymózgowiu. Blaszka p o k r y w y śródmózgowia ( l a m i n a tecti mesencephali) leży po stronie grzbietowej, tj. nad wodociągiem, i jest zakryta sięgającymi jeszcze dalej ku t y ł o w i półkulami mózgowymi. Blaszka ta składa się z dwóch par w z g ó r k ó w : górnych {colliculi superiores) i dolnych (colliculi inferiores). W ścianie dolnej wodociągu mózgu znajdują się jądra ruchowe n e r w ó w III i IV. Nerw III okoruchowy (n. oculomotorius) wychodzi ze śródmózgowia na powierzchni podstawnej mózgu w dole międzykonarowym, natomiast nerw IV, bloczkowy (n. trochlearis) jest jedynym spośród n e r w ó w mózgowych, który ukazuje się na powierzchni grzbietowej pnia mózgu, skąd kieruje się ku podstawie wokół bocznej powierzchni konara mózgowego.
Międzymózgowie i jądra podkorowe kresomózgowia Półkule mózgowe, podobnie jak i pozostałe części mózgowia, zbudowane są z istoty białej i szarej. Istota szara, umieszczona na powierzchni półkul mózgowych, nosi nazwę kory mózgowej, natomiast skupienie tejże istoty we wnętrzu istoty białej półkul mózgowych nazywamy jądrami p o d k o r o w y m i kresomózgowia. Należą one rozwojowo do starszych i prymitywniejszych części ośrodkowego układu nerwowego, w odróżnieniu od kory mózgowej, która zajmuje szczególne miejsce jako najwyższy ośrodek regulujący i kierujący czynnościami całego ustroju. Pozostałość kanału środkowego w obrębie międzymózgowia ma kształt szczeliny, zwanej komorą trzecią, ustawionej w płaszczyźnie strzałkowej. W części tylnej komora trzecia łączy się z w o d o c i ą g i e m mózgu, od przodu zaś komunikuje się za pośrednictwem dwóch o t w o r ó w międzykomorowych z kom o r a m i bocznymi, położonymi we wnętrzu półkul mózgowych. '427
Ściana boczna komory trzeciej utworzona jest po każdej stronie z dużego skupienia istoty szarej o kształcie jajowatym, zwanego w z g ó r z e m (fha/amus). Grzbietowa i przyśrodkowa powierzchnie wzgórza są w o l n e ; powierzchnia boczna, jak już wspomniano, zrasta się z torebką wewnętrzną, zaś powierzchnia dolna łączy się z tzw. okolicą podwzgórzową. Na ścianie bocznej komory trzeciej widnieje rowek, który biegnie od wejścia do wodociągu mózgu ku przodowi do o t w o r u międzykomorowego, zwany bruzdą podwzgórzową ( sulcus hypothalamicus). Część bocznej ściany komory trzeciej położoną poniżej tej bruzdy nazywamy p o d w z g ó r z e m (hypothalamus). W obrębie podwzgórza znajdują się skupienia istoty szarej, z których najważniejsze tworzy niewielkie wypuklenie, zwane guzem popielatym ( tuber cinereum). Guz popielaty przechodzi ku d o ł o w i w lejek (infundibulum), na którym zawieszona jest przysadka (hypophysis) bardzo ważny gruczoł dokrewny, leżący w zagłębieniu siodła tureckiego. Podwzgórze otrzymuje włókna czuciowe od wzgórza, samo zaś ma połączenie z jądrami podkorowymi kresomózgowia oraz ze śródmózgowiem, do których wysyła impulsy. Ścianę dolną, czyli dno komory trzeciej, tworzą, w y m i e n i o n e już przy opisie podstawy mózgu: skrzyżowanie wzrokowe, położony za nim guz popielaty i lejek, dalej ciała suteczkowate i istota dziurkowana tylna. Ścianę tylną komory trzeciej tworzy pasmo włókien poprzecznych, zwane spoidłem t y l n y m mózgu (comissura cerebri posterior), pod którym znajduje się wejście do wodociągu mózgu. Nad t y m spoidłem leży szyszynka ( c o r p u s pineale), którą otacza ze wszystkich stron opona miękka, zawierająca liczne naczynia krwionośne, przechodzące tu z pokrywy komory trzeciej. Ścianę górną komory trzeciej tworzy blaszka naczyniowa nabłonkowa. Jest to nie rozwinięta, bardzo cienka pozostałość ściany pęcherzyka nfiiędzymózgowia, która zrasta się z pokrywą naczyniówkową komory trzeciej utworzoną przez oponę miękką. W komorze trzeciej znajduje się skupienie istoty szarej łączącej oba wzgórza, zwane zrostem m i ę d z y w z g ó r z o w y m (adhesio interthalamica). Bocznie od wzgórza, już w obrębie półkuli mózgowej, leży jądro soczewkow a t e (nuc/eus lentiformis), oddzielone od wzgórza torebką wewnętrzną. Torebka w e w n ę t r z n a (capsula interna) stanowi skupienie włókien nerwowych, które biegną od kory do konarów mózgowych i dalej przez piramidy do rdzenia kręgowego. Przez torebkę wewnętrzną przechodzi ważna droga ruchowa - korowo-rdzeniowa (piramidowa). W jądrze soczewkowatym rozróżniamy dwie części, przedzielone cienkim pasmem istoty białej: położoną bliżej linii środkowej gałkę bladą (globus pallidus) oraz część boczną - skorupę (putamen). Jądro ogoniaste (nucleus caudatus) le^y bocznie i powyżej wzgórza. Ma ono kształt dużego przecinka, w którym rozróżniamy głowę, ciało i ogon. Głowa jądra ogoniastego jest zwrócona ku przodowi, zwężający się ogon kieruje się ku tyłowi, w bok i ku dołowi. Jądro ogoniaste i skorupa ze względu na wspólne pochodzenia nazywane '428
są łącznie ciałem p r ą ż k o w a t y m {corpus striatum) albo prążkowiem {striatum), gałkę bladą nazywa się pallidum. Zarówno striatum jak i pallidum są jądrami ruchowymi. Wychodzą z nich impulsy ruchowe do jąder ruchowych pnia mózgowego, a cały układ jąder podkorowych kresomózgowia w r a z z d r o g a m i , które z nich wychodzą, nosi nazwę układu ruchowego pozapiramidowego. Striatum, jak i pallidum mają też połączenia ze wzgórzem oraz z korą mózgową. Międzymózgowie jest częścią ośrodkowego układu nerwowego, która zwierzętom niższym, ze słabo rozwiniętą korą mózgową, może zapewnić zdolność reagowania i zachowania się w s p o s ó b podobny, w jaki zachowuje się zwierzę nie pozbawione półkul mózgowych. U zwierząt wyższych, którym wycięto korę mózgową, ale pozostawiono międzymózgowie, występuje szereg zaburzeń, dotyczących przede wszystkim ruchów dowolnych. Jak w s p o m n i a n o , wzgórze jest p o d k o r o w y m ośrodkiem czuciowym. Przez wzgórze przechodzą wszystkie bez wyjątku bodźce powstające w receptorach obwodowych. Część tych bodźców zostaje przekazana do kory mózgowej, część zaś jest skierowana przez wzgórze do odpowiednich jąder podwzgórza, jąder podkorowych kresomózgowia, jąder śródmózgowia. Za pośrednictwem wzgórza odbywają się liczne i złożone czynności odruchowe na drogach przechodzących przez wspomniane wyżej jądra ruchowe. Własnym jądrem ruchowym międzymózgowia jest gałka blada ( g l o b u s pallidus), która reguluje i koordynuje czynności niższych odcinków pnia mózgowego - śródmózgowia i rdzenia przedłużonego. Łuki odruchowś, które w swojej części czuciowej mają wzgórze, a w ruchowej - gałkę bladą, są drogą, po której przebiegają bodźce nerwowe, wyzwalające tak złożone czynności odruchowe! jak chód, bieg, różne odruchy obronne, pokarmowe, płciowe. Odruchy te są wrodzone i utrwalone dziedzicznie, tzn. same pojawiają się po urodzeniu i są niezmienne dla pewnego gatunku; noszą one także nazwę o d r u c h ó w bezwarunkowych. Odruchy te znikają u chorych z uszkodzoną gałką bladą; nie występuje u nich odruch obronny na nagły dźwięk, światło itp. W gałce bladej powstają również impulsy nerwowe, które są hamowane przez ośrodek nadrzędny, jakim jest dla niej ciało prążkowane, i które wskutek tego w warunkach prawidłowych nie ujawniają się. Są to stare automatyzmy ruchowe, które dla człowieka straciły swe znaczenie (np. ruchy wykonywane przy wspinaniu się na drzewo). Pojawiają się one w razie uszkodzenia lub zniszczenia ciała prążkowanego i noszą nazwę ruchów atetotycznych. Ruchy te występują również u noworodków, u których prążkowie nie jest jeszcze w pełni rozwinięte. Gałka blada uczestniczy również przy w y k o n y w a n i u ruchów dowolnych, dla których bodźce nerwowe powstają w korze mózgowej, oraz odruchowych. W razie uszkodzenia gałki bladej ruchy stają się niezgrabne, mało sprężyste, jednostajne. Podwzgórze jest ważnym ośrodkiem licznych czynności w e g e t a t y w n y c h . Ma tu niewątpliwie duże znaczenie bliskie sąsiedztwo z tak ważnym gruczołem dokrewnym, jakim jest przysadka mózgowa. Po zniszczeniu lub uszkodzeniu '429
ośrodków guza popielatego występują zaburzenia przemiany materii, zaburzenia naczynioruchowe, poty, skurcze jelit, macicy, pęcherza, zmiany w składzie krwi, zaburzenia regulacji cieplnej. Poznawszy budowę i czynności poszczególnych odcinków ośrodkowego układu nerwowego stwierdzamy, że wyższy odcinek tego układu stanowi jak gdyby nadbudowę złożoną z łuków odruchowych i ośrodków, które stają się coraz bardziej złożone, ale też coraz doskonalsze, i podporządkowują sobie niższe odcinki (piętra). Rdzeń przedłużony jest nadbudową rdzenia kręgowego, śródmózgowie podporządkowuje sobie rdzeń przedłużony, a dzięki ośrodkom słuchowym i w z r o k o w y m umożliwia ono organizmowi doskonalsze reagowanie na zjawiska świata zewnętrznego, niżby to było możliwe tylko za pomocą rdzenia przedłużonego. Jeszcze wyższe piętro pnia mózgowego międzymózgowie i jądra podkorowe - umożliwia wykonywanie już bardzo złożonych i doskonałych czynności, zwanych instynktami i odruchami bezwarunkowymi. Pojawienie się w procesie rozwoju filogenetycznego kory m ó z g o w e j , która najwyższy stopień rozwoju osiągnęła u człowieka, daje nieograniczone w p r o s t możliwości nabywania nowych, indywidualnych reakcji, uwarunkowanych przez przedmioty i zjawiska świata zewnętrznego oraz wewnętrznego i stawia człowieka na szczycie drabiny ewolucyjnej świata istot żywych.
Kreso m ózgo wie Kresomózgowie (telencephalon) jest tą częścią układu nerwowego, która w sposób istotny różni człowieka od pozostałych przedstawicieli świata zwierzęcego. Kresomózgowie osiąga u człowieka najwyższy stopień rozwoju; przewyższa ono wszystkie pozostałe części mózgowia zarówno rozmiarami, jak i różnorodnością funkcji. Powstając z pierwszego pęcherzyka mózgowego, kresomózgowie przybiera w trakcie rozwoju embrionalnego postać dwóch półkul m ó z g o w y c h ( h e m i s p h e r i a cerebri), połączonych ze sobą szerokim i dość g r u b y m pęczkiem istoty białej, zwanej ciałem m o d z e l o w a t y m ( c o r p u s callosum).
Półkule mózgowe
W skład półkul m ó z g o w y c h ( h e m i s p h e r i a cerebri) wchodzi płaszcz m ó z g u (pallium), w ę c h o m ó z g o w i e ( r h i n e n c e p h a l o n ) i o m ó w i o n e już wyżej jądra podkorowe tworzące ciało prążkowane ( c o r p u s striatum). Płaszcz składa się z istoty białej pokrytej od zewnątrz istotą szarą, czyli korą (cortex cerebri). Istota biała utworzona jest z czterech rodzajów włókien nerwowych. Są to włókna: 1) odkorowe lub odśrodkowe, tj. odchodzące od komórek kory mózgowej, 2) dośrodkowe, tj. zdążające do komórek kory, 3) asocjacyjne, łączące różne okolice kory mózgowej i 4) spoidłowe, łączące półkule mózgowe. Kora Kora mózgowa tworzy na powierzchni mózgu liczne fałdy. Gdyby można było je rozprostować i wygładzić, kora zajęłaby powierzchnię odpowiadającą '430
kwadratowi o boku 50 cm i grubości ok. 3 m m . Kora mózgowa składa się z gęsto upakowanych komórek nerwowych różnej wielkości i kształtu, tworzących kilka warstw. Ich liczbę ocenia się na ok. 10 miliardów. Każda z tych komórek otrzymuje informacje od setek innych neuronów przekazywane przez ich aksony. Na każdej półkuli mózgowej odróżniamy trzy powierzchnie: wypukłą ( facies convexa cerebri), przyśrodkową ( facies medialis cerebri) i podstawę mózgu (basis cerebri). Jak już wspomniano, powierzchnię półkul pokrywają fałdy zwane zakrętami m ó z g u (gyricerebri), porozdzielane bruzdami (sulcicerebri) i szczelinami (fissurae). U zarodka czteromiesięcznego powierzchnia półkul jest jeszcze niemal zupełnie gładka i dopiero w 5 - 6 miesiącu zaczyna pokrywać się bruzdami, których liczba wzrasta w miarę dalszego rozwoju. W każdej półkuli rozróżniamy cztery płaty (lobi): 1) płat czołowy ( lobus frontaiis), 2) płat ciemieniowy ( lobus parietalis), 3) płat s k r o n i o w y ( lobus temporalis) i 4) płat potyliczny ( lobus occipitalis). Płat czołowy oddziela od płata ciemieniowego głęboka bruzda środkowa (sulcus ceritraliś). Bruzda ta biegnie na powierzchni wypukłej prawie prostopadle do szczeliny bocznej, do której zbliża się na odległość około 2 cm. Bruzda boczna mózgu (sulcus lateralis) oddziela płat skroniowy od płata czołowego i ciemieniowego. Jest to jedna z najgłębszych bruzd mózgu. Rozpoczyna się na podstawie mózgu w pobliżu istoty dziurkowanej przedniej, skąd przechodzi na powierzchnię wypukłą mózgu, biegnąc najpierw ku górze, a następnie ku tyłowi. Od bruzdy bocznej odchodzi kilka odgałęzień, a mianowicie: gałąź wstępująca, gałąź przednia oraz skierowany ku tyłowi i ku górze odcinek końcowy bruzdy, zwany gałęzią tylną. Płat ciemieniowy zajmuje okolicę położoną ku tyłowi od bruzdy środkowej. Na wypukłej powierzchni półkuli nie ma wyraźnej granicy pomiędzy płatem ciemieniowym a potylicznym. Na powierzchni przyśrodkowej granicę tę tworzy bruzda ciemieniowo-potyliczna. Tylko nieznaczna część tej szczeliny wychodzi na powierzchnię wypukłą półkuli, nacinając jej górną krawędź. W obrębie każdego z wyżej w y m i e n i o n y c h płatów rozróżniamy szereg zakrętów i bruzd. W płacie c z o ł o w y m , ku przodowi od bruzdy środkowej, przebiega równolegle do niej ułożony zakręt p r z e d ś r o d k o w y (gyrus precentralis). Od przodu ogranicza go bruzda przedśrodkowa (sulcus precentralis). W kierunku od góry ku d o ł o w i przebiegają jeden po d r u g i m trzy zakręty czołowe: górny, środkowy i dolny, rozdzielone dwiema bruzdami: czołową górną i czołową dolną. Zakręt c z o ł o w y górny ( g y r u s frontaiis superior) rozpoczyna się na przyśrodkowej powierzchni półkuli i dochodzi do bruzdy czołowej górnej. Zakręt c z o ł o w y ś r o d k o w y ( g y r u s frontaiis medius) leży pomiędzy bruzdą czołową górną a dolną, zakręt czołowy dolny - między bruzdą czołową dolną a szczeliną boczną. Dwie przednie gałęzie szczeliny bocznej wrzynają się w obszar zakrętu c z o ł o w e g o d o l n e g o ( g y r u s frontaiis interior) i dzielą go na trzy części: przednią - część oczodołową (pars orbitalis) położoną pomiędzy gałęzią przednią a przednio-dolnym brzegiem płata czołowego; środkową - część trójkątną ( pars triangularis), pomiędzy gałęzią wstępującą a przednią, oraz tylną - część wieczkową (pars opercularis), ograniczoną od przodu gałęzią wstępującą. Na powierzchni dolnej płata czołowego przebiega równolegle do szczeliny podłużnej mózgu bruzda w ę c h o w a (sulcus olfactorius), do której przylega opuszka w ę c h o w a ( b u l b u s olfactorius)
'431
i p a s m o w ę c h o w e ( t r a c t u s olfactorius). Pomiędzy bruzdę w ę c h o w ą a brzegiem przyśrodkowym półkuli leży zakręt prosty (gyrus rectus), bocznie od bruzdy w ę c h o w e j znajduje się szereg bruzd o c z o d o ł o w y c h , rozdzielających zakręty oczodołowe ( sulci et gyri orbitales). W płacie c i e m i e n i o w y m wzdłuż bruzdy środkowej rozciąga się zakręt z a ś r o d k o w y (gyrus postcentraiis), ograniczony od tyłu bruzdą z a ś r o d k o w ą ( sulcus postcentralis). Za tą bruzdą, prawie równolegle do krawędzi górnej półkuli, biegnie bruzda międzyciemieniowa (su/cus interparietalis), dzieląca tylną okolicę płata ciemieniowego na dwie części: płacik c i e m i e n i o w y górny ( l o b u l u s parietalis superior) i płacik c i e m i e n i o w y dolny ( l o b u l u s parietalis interior). W granicach płacika ciemieniowego dolnego rozróżniamy dwa zakręty: zakręt nadbrzeżny (gyrus supramarginalis), zamykający zakończenie szczeliny bocznej i zakręt k ą t o w y ( g y r u s angularis), otaczający zakończenie bruzdy s k r o n i o w e j górnej (su/cus temporalis superior). Na zewnętrznej powierzchni płata potylicznego występuje kilka bruzd, które są osobniczo zmienne i wskutek tego powierzchnię tę t r u d n o jest podzielić na określone zakręty. W płacie s k r o n i o w y m rozróżniamy d w i e bruzdy skroniowe, biegnące równolegle do szczeliny bocznej. Bruzda skroniowa górna i dolna (sulcus temporalis superior et interior) leżą na powierzchni bocznej płata skroniowego, dzieląc go na trzy zakręty: skroniowy górny, ś r o d k o w y i dolny ( g y r u s temporalis superior, medius et interior). Zakręt s k r o n i o w y dolny przechodzi przez dolną powierzchnię płata skroniowego. Wzdłuż brzegu zewnętrznego płata skroniowego biegnie krótka bruzda potyliczno-skroniowa (sulcus occipitotemporaiiś). Na powierzchni dolnej płata skroniowego widnieje bruzda poboczna ( sulcus collateralis), na zewnątrz od tej szczeliny leży zakręt potyliczno-skroniowy p r z y ś r o d k o w y ( g y r u s occipitotemporaiiś medialis), przyśrodkowo zakręt hipokampa (gyrus parahippocampalis). Przednia część zakrętu hipokampa tworzy zgrubienie, zwane hakiem (uncus). Wpuklona część powierzchni mózgu, w y s p a (insuia), leży w głębi dołu bocznego mózgu i jest całkowicie zakryta przez część płata czołowego, ciemieniowego i skroniowego. Ma ona kształt piramidy, zwróconej podstawą ku górze. Bruzdy i zakręty przyśrodkowej powierzchni półkul mózgu udostępniamy badaniu, oddzielając od siebie obie półkule cięciem strzałkowym w płaszczyźnie pośrodkowej. Rozróżniamy tu bruzdy i zakręty długie i krótkie. Bezpośrednio nad spoidłem w i e l k i m biegnie bruzda ciała m o d z e l o w a t e go (sulcus corporis callosi), a nad nią długi, łukowato w y g i ę t y zakręt obręczy ( g y r u s cinguli), ograniczony od góry bruzdą obręczy (sulcus cinguli). Za pośrednictwem wąskiego pasma, zwanego cieśnią zakrętu obręczy (isthmus gyri cinguli), łączy się on z zakrętem hipokampa. Końcowa część bruzdy obręczy wznosi się ku górze krawędzi półkuli tworząc wygiętą ku t y ł o w i część brzeżną bruzdy obręczy. Nieco ku przodowi odchodzi od bruzdy obręczy ku górze niewielka bruzda okołośrodkowa. Część przyśrodkowej powierzchni półkuli położona pomiędzy t y m i bruzdami nazywa się płacikiem o k o ł o ś r o d k o w y m (lobulus paracentraiis). Ku t y ł o w i od części brzeżnej bruzdy obręczy leży przedklinek ( p r e c u n e u s ) , ograniczony od tyłu głęboką szczeliną ciemieniowo-potyliczną (sulcus parietooccipitaiis). Dalej ku t y ł o w i leży klinek (cuneuś), ograniczony od dołu bardzo głęboką bruzdą o s t r o g o w ą (sulcus calcarinus).
Komory boczne półkul m ó z g o w y c h We wnętrzu każdej półkuli mózgowej znajduje się szczelinowata pozostałość światła pierwotnego pęcherzyka mózgowego, zwana komorą boczną (ventriculus lateraliś). Komora boczna składa się z czterech części, z których każda leży w innym płacie. Są to: 1) róg przedni - w płacie czołowym, 2) część środkowa - w płacie ciemieniowym, 3) róg t y l n y - w płacie potylicznym i 4) róg dolny - w płacie skroniowym. Każda z komór bocznych łączy się z komorą trzecią za pośrednictwem o t w o r u międzykomorowego.
'432
Węchomózgowie W ę c h o m ó z g o w i e ( r h i n e n c e p h a l o n ) składa się z części o b w o d o w e j i części ośrodkowej, czyli korowej. Do części o b w o d o w e j należą: opuszka w ę c h o w a , pasmo w ę c h o w e i trójkąt w ę c h o w y . O b w o d o w a część składa się g ł ó w n i e z włókien n e r w o w y c h , przewodzących bodźce w ę c h o w e do części korowej. Najważniejszymi składnikami części ośrodkowej w ę c h o m ó z g o w i a są: o m ó w i o n y wyżej zakręt obręczy i jego cieśń oraz wpuklający się do wnętrza rogu dolnego k o m o r y bocznej hipokamp. Włókna n e r w o w e , łączące różne okolice w ę c h o m ó z g o w i a o b u półkul, tworzą tzw. sklepienie (fornix). Ma o n o kształt litery X, która swą częścią środkową zrośnięta jest z ciałem m o d z e l o w a t y m i bierze udział w utworzeniu stropu k o m o r y trzeciej.
Ośrodki korowe Liczne obserwacje kliniczne oraz badania polegające na drażnieniu słabym prądem elektrycznym różnych obszarów kory mózgowej odsłoniętych w czasie zabiegów neurochirurgicznych świadczą o tym, że pewne okolice kory są związane z określonymi czynnościami. Okolice takie nazywamy ośrodkami korowymi. Ośrodkami ruchowymi (lub polem ruchowym) kory są te jej części, które po podrażnieniu wywołują reakcję ruchową z grup mięśniowych. Ośrodki ruchowe położone Sq w paśmie kory tworzącym zakręt przedśrodkowy. Znajdują się tu komórki olbrzymie (Betza) o kształcie piramidy, zwane także komórkami piramidalnymi. Aksony tych komórek biegną przez torebkę wewnętrzną, śródmózgowie i most do rdzenia, tworząc szlak mózgowo-rdzeniowy, zwany też piramidowym. Droga ta zawiera ok. 34000 długich włókien mielinowych po każdej stronie. Włókna te krzyżują się, tj. włókna idące od lewej półkuli przechodzą na stronę prawą rdzenia kręgowego. Skrzyżowanie to następuje w obrębie pnia mózgowego. Skutkiem tego mięśnie jednej strony ciała pozostają pod kontrolą ośrodków strony przeciwnej. Dlatego uszkodzenie, np. przez wylew krwi, pola ruchowego lub włókien szlaku korowo-rdzeniowego ponad poziomem ich skrzyżowania powoduje porażenie mięśni po stronie przeciwnej ciała. Obszar kory odpowiadający zakrętowi zaśrodkowemu, zwany polem somestetycznym, zawiera ośrodki czuciowe, odbierające wrażenia dotyku, ciepła, chłodu oraz informacje o ruchach mięśniowych. Czucie bólu prawdopodobnie nie jest reprezentowane w korze mózgowej. Wrażenia słuchowe docierają do płatów skroniowych. Ośrodki słuchowe znajdują się w zakręcie skroniowym górnym. Okolice przylegające do pola słuchowego pełnią funkcję kojarzeniową, związaną z interpretacją dźwięków m. in. rozumieniem mowy. Zniszczenie okolicy tylnej zakrętu skroniowego górnego powoduje utratę zdolności rozumienia słów. Nazywa się to afazją słuchową. Chory słyszy, co się do niego mówi, ale nic z tego nie rozumie, tak jakby słyszał język obcy całkowicie mu nie znany. Płaty skroniowe prawdopodobnie są także siedzibą pamięci. Drażnienie 28-Anatomia i fizjologia
433
odsłoniętej powierzchni kory płata skroniowego wywołuje u badanego żywe obrazy z dalekiej nieraz przeszłości. Również drgawki epileptyczne zaczynające się w tych okolicach kory często są poprzedzone aurą w postaci żywych wspomnień i scen z dawno minionej przeszłości. Pole wzrokowe znajduje się w płacie potylicznym. Bodźce wzrokowe z prawych połówek obu siatkówek docierają do pola wzrokowego prawej półkuli i tak samo jest ze stroną przeciwną, toteż zniszczenie ośrodka wzrokowego po jednej stronie mózgu powoduje ubytek pola widzenia tylko w odpowiednich połówkach siatkówek. Okolice przedczołowe położone ku przodowi od pola ruchowego są uważane za siedlisko wyższych czynności mózgu, związanych z intelektem, inteligencją, sferą moralną człowieka. Człowiek pozbawiony okolic przedczołowych zdolny jest do wykonywania wielu prostych zadań intelektualnych, ma jednak trudności lub w ogóle nie jest zdolny do tworzenia złożonych konstrukcji myślowych prowadzących do wyznaczonego celu, do pogłębionego i abstrakcyjnego myślenia. Przypuszcza się, że w okolicach przedczołowych następuje zapamiętywanie wielkiej liczby informacji i powstawanie na ich podstawie konstrukcji myślowych, takich jak planowanie zachowania się w przyszłości, świadome opóźnianie reakcji, wybór najbardziej odpowiedniej reakcji, przewidywanie skutków czynności ruchowych, rozwiązywanie skomplikowanych zadań logicznych (matematycznych, prawnych, filozoficznych), kontrola zachowania się wg wzorców społecznych i moralnych. Osoby pozbawione okolic przedczołowych działają impulsywnie w sposób nie kontrolowany, zatracają wszelkie zasady moralne, tracą poczucie wstydu, są emocjonalnie bardzo zmienne,łatwo przechodzą od radości do przygnębienia, od uczuć przyjaznych do gniewu i nienawiści. Do bardziej złożonych czynności kory mózgowej należą także czynności poznawcze wyższego rzędu - praksyjne i gnostyczne. Noworodek przynosi ze sobą na świat tylko bardzo prymitywne ruchy wrodzone. Stopniowo dziecko doskonali coraz bardziej swe ruchy na drodze naśladowania, powtarzania i ćwiczenia, dzięki czemu uzyskuje zdolność wykonywania czynności zamierzonych, celowych, czyli praksji. Praksja jest wynikiem współdziałania obszernych terytoriów kory mózgowej, jednakże ostateczny efekt ruchowy czynności praksyjnych dochodzi do skutku za pośrednictwem ośrodków ruchowych. Utratę zdolności wykonywania celowych ruchów złożonych nazywamy aprąksją. Chory z apraksją staje się zupełnie bezradny. Nie potrafi sobie zapiąć guzika, zasznurować butów, posługiwać się widelcem, łyżką itp., gdyż „zapomina", jakie ruchy są potrzebne do wykonywania tych czynności. Gnozją nazywamy zdolność poznawania przedmiotów i zjawisk oraz oceny tych wrażeń, jakich dostarczają nam zmysły. Rozróżniamy różne rodzaje gnozji. Stereognozją nazywamy zdolność rozpoznawania przedmiotów dotykiem. Ośrodki stereognozji mieszczą się w płacie ciemieniowym ku tyłowi od zakrętu '434
zaśrodkowego. Porażenie tych okolic pociąga za sobą tzw. astereognozję, tj. niezdolność rozpoznawania przedmiotów dotykiem. Wspomnieliśmy już, że ośrodki gnozji słuchowej mieszczą się w g ó r n y m zakręcie skroniowym. Gnozją słuchową nazywamy nabytą w życiu osobniczym zdolność rozpoznawania przedmiotów według charakteryzujących je dźwięków. I tak np. zegarek rozpoznajemy po tykaniu, samolot po warkocie motoru, ludzi po głosie itp. Gnozja w z r o k o w a lokalizuje się w płacie potylicznym na jego powierzchni wewnętrznej i zewnętrznej. Gnozją wzrokową nazywamy zdolność poznawania przedmiotów z ich wyglądu. Ośrodki m o w y Mowa jako sposób wyrażania myśli jest cechą wyłącznie ludzką. Mówiąc w y d a j e m y szereg artykułowanych dźwięków, z których składają się słowa i zdania wyrażające m y ś l i - j e s t to ruchowa część m o w y . Słysząc cudzą m o w ę rozumiemy ją, wiążemy pewne słowa z pewnymi przedmiotami i zjawiskami tę część m o w y nazywamy częścią słuchową. Zaburzenia m o w y w y w o ł a n e porażeniem korowych ośrodków m o w y nazywają się afazją. Ośrodki m o w y są umieszczone w korze mózgowej niesymetrycznie: u praworęcznych w lewej półkuli, u leworęcznych - w prawej. Istnieje ośrodek m o w y czuciowy i ruchowy. Ośrodek czuciowy (słuchowy) m o w y (Wernickego) znajduje się w tylnej części zakrętu skroniowego górnego. Przy porażeniu tej okolicy chory traci zdolność rozumienia słów. Nazywamy to afazją słuchową. Ośrodek ruchowy m o w y (Broca) leży w tylnej części zakrętu czołowego dolnego. Po porażeniu tej okolicy występuje afazja ruchowa. Chory nie potrafi mówić, m i m o że pozostaje mu zdolność wydawania dźwięków i modulacji głosu, ponieważ wypadają u niego ruchowe wzory słów i traci on możność zestawiania dźwięków i sylab. Istnieją wreszcie w korze mózgowej ośrodki czytania i pisania. Pierwszy leży w zakręcie kątowym płata ciemieniowego, drugi - w tylnej części środkowego zakrętu czołowego. Elektroencefalografia Czynności ośrodkowego układu nerwowego towarzyszą pewne zjawiska elektryczne, które potrafimy rejestrować i wykorzystać dla celów praktycznych w neurologii. Jeżeli do czaszki lub obnażonego mózgu przyłożymy dwie elektrody i połączymy j e z o d p o w i e d n i m wzmacniaczem i przyrządem rejestrującym, to otrzymamy zapis aktywności elektrycznej mózgu w postaci krzywej, zwanej elektroencefalogramem (eeg). Prawidłowy eeg u człowieka pozostającego w spoczynku z zamkniętymi oczami ma postać krzywej składającej się z fal o częstości 8 - 1 2 Hz, które układają się w postaci wrzecion. Fale takie noszą '435
nazwę fal alfa. Jeżeli badanemu polecimy otworzyć oczy lub wykonać jakiś wysiłek umysłowy, wówczas rytm alfa znika i krzywa eeg przybiera postać drobnych nieregularnych oscylacji. W pewnych chorobach mózgu, jak np. padaczka lub guzy mózgu, w zapisie eeg pojawiają się charakterystyczne zmiany pozwalające na bliższe zlokalizowanie ogniska chorobowego. ODRUCHY WARUNKOWE Badanie czynności tak skomplikowanego narządu, jakim jest mózg, nastręcza duże trudności. Jedną z metod dających wgląd w mechanizm niektórych czynności ośrodkowego układu nerwowego jest metoda Odruchów warunkowych. Metoda ta związana jest z nazwiskiem rosyjskiego fizjologa Pawłowa, który je opracował. W pewnym okresie za pomocą teorii odruchów warunkowych próbowano tłumaczyć wszystkie czynności układu nerwowego nie wyłączając właściwych tylko człowiekowi tzw. wyższych czynności, jak działalność intelektualna i funkcja sfery psychicznej. Chociaż sprawa jest bardziej skomplikowana, odruchy warunkowe stanowią niewątpliwie jeden z lepiej poznanych i doświadczalnie udokumentowanych mechanizmów funkcjonowania układu nerwowego i dlatego pokrótce je tu przedstawimy. Uprzednio omówione odruchy (bezwarunkowe) odbywają się przy udziale ośrodków położonych w rdzeniu kręgowym lub na niższych (podkorowych) piętrach mózgu. Odruchy bezwarunkowe są wrodzone, utrwalone dziedzicznie i właściwe dla całego gatunku. W odruchach warunkowych zasadniczą rolę odgrywa kora mózgowa. Powstają one w trakcie rozwoju i życia osobniczego i mają charakter nabyty, nie są niezmienne - powstają, utrwalają sję lub wygasają w zależności od warunków zewnętrznych - są zawsze w ja kiś sposób uwarunkowane czynnikami zewnętrznymi. Jeżeli psu wlejemy do pyska rozcieńczony roztwór jakiegokolwiek kwasu, wywoła to reakcję obronną, kwas zostanie wyrzucony na zewnątrz, a równocześniezacznie obficie wydzielać się ślina, która rozcieńcza kwas i zmywa go ze śluzówki jamy ustnej. Jest to odruch prosty, czyli bezwarunkowy. Jeżeli tuż przed wlaniem kwasu zadziałamy na psa jakimkolwiek bodźcem zewnętrznym, np. dzwonkiem, i bodziec ten skojarzymy kilkakrotnie z wlewaniem kwasu, to po pewnym czasie reakcja obronna i wzmożone wydzielanie śliny zacznie powstawać na sam dźwięk dzwonka. Ten odruch na bodziec pierwotnie obojętny nazwał Pawłów odruchem warunkowym. Odruchy bezwarunkowe są stałe, niezmienne, ulegają tylko nieznacznym wahaniom i dają się z łatwością wywołać u każdego zwierzęcia. Mają one gotowy układ dróg nerwowych, którymi biegnie pobudzenie - zamknięty układ przewodników; natomiast odruchy warunkowe są mniej lub bardziej przypadkowe, powstanie ich wymaga czynników dodatkowych, dla ich wytworzenia musi nastąpić utorowanie pewnego odcinka drogi pobudzenia. Odcinek ten znajduje się w korze mózgowej, gdzie następuje proces podobny '436
do zamykania obwodu elektrycznego, umożliwiający przejście pobudzenia z dróg czuciowych na drogi odśrodkowe. Odruchy warunkowe zachodzą przy udziale kory mózgowej, tzn. droga łuku odruchowego przebiega przez ośrodki w korze mózgowej. Odruchy bezwarunkowe należą do czynności niższych pięter układu nerwowego ośrodkowego. Wycięcie kory półkul mózgowych u psa pociąga za sobą zniknięcie wszystkich wytworzonych odruchów warunkowych. Nowych wytworzyć się nie udaje. Natomiast odruchy bezwarunkowe po t y m zabiegu nie zmieniają się zupełnie. U zwierząt, które nie mają kory mózgowej, odruchy warunkowe powstają przy udziale najwyższego piętra ich ośrodkowego układu nerwowego. Odruchy warunkowe powstają na podstawie odruchów bezwarunkowych. Do powstania odruchu warunkowego konieczna jest zbieżność w czasie jakiegoś bodźca obojętnego dla ustroju z odbywającym się odruchem bezwarunkowym. Po takim skojarzeniu powstaje między tym bodźcem a czynnością bezwarunkową związek czasowy. Bodziec dotychczas obojętny staje się bodźcem warunkowym. Nabiera znaczenia sygnału, który umożliwia ustrojowi wcześniejszą reakcję na zmiany środowiska zewnętrznego, zanim te zmiany na ustrój bezpośrednio zadziałają.
SEN Sen jest stanem ustroju, w którym występują zmiany szeregu jego czynności. Najistotniejszym objawem snu jest obniżenie aktywności układu nerwowego, a przede wszystkim kory mózgowej, co pociąga za sobą utratę łączności z otoczeniem. Bezsenność jest jednym z często spotykanych zaburzeń czynności mózgu. Sen jest nieodzowną potrzebą ustroju. Człowiek przebywa w stanie snu przez V3 część swego życia. Długość snu u człowieka jest zależna od wieku. Noworodek śpi 21 h na dobę, dziecko 3-miesięczne - 18 h, półroczne - 14 h, 4-letnie - 12 h. W miarę lat życia długość snu stopniowo się zmniejsza. Począwszy od 17-19 roku życia długość snu ogranicza się do 8 h na dobę. Brak snu prowadzi do wystąpienia poważnych zaburzeń w ośrodkowym układzie nerwowym. W doświadczeniach przeprowadzonych na ochotnikach, którzy nie spali po 112 h, spostrzegano po 30-60 h bezsenności utratę pamięci, wzrost pobudzenia, halucynacje, reakcje schizofreniczne. Głębokość snu jest różna w różnych jego okresach. Głębokość snu bada się w ten sposób, że przy śpiącym w pewnym momencie uruchamia się jakiś sygnał dźwiękowy. Miarą głębokości snu jest siła bodźca powodującego przebudzenie. Doświadczenia te wykazują, że u większości ludzi sen jest najgłębszy w pierwszej, rzadziej w drugiej godzinie po zaśnięciu. W czasie snu zmniejsza się napięcie mięśniowe. W stanie skurczu pozostaje jedynie mięsień okrężny oka zamykający powieki i mięsień prosty oka górny, który pociąga gałkę oczną ku górze, podciągając ją pod powiekę górną. Rytm oddechowy w czasie snu ulega zwolnieniu. Przemiana gazowa i podstawowa '437
obniża się o 10-15%, Ciśnienie krwi spada o 10-30 mm Hg. Tętno ulega zwolnieniu. Istnieje kilka teorii snu. Wiadomo, że wszelkie sytuacje sprowadzające do m i n i m u m napływ bodźców do receptorów czuciowych sprzyjają wystąpieniu snu naturalnego (zaciemnienie pomieszczenia, ciepło, cisza, rozluźnienie mięśni). Strach i emocje utrudniają zasypianie. Stąd hipoteza, że sen jest następstwem zmniejszenia napływu bodźców do ośrodkowego układu nerwowego. Biorąc pod uwagę to, że najważniejsze przejawy snu mają niewątpliwy związek ze zmianą czynności mózgu, szukano wyjaśnienia tego stanu w analizie procesów, jakie przy t y m w mózgu zachodzą. Wyniki badań eksperymentalnych Pawłowa dały mu podstawę do stwierdzenia, że stan snu jest identyczny ze stanem hamowania wewnętrznego. Pawłów zauważył mianowicie w czasie badań nad odruchami w a r u n k o w y m i , że psy często zasypiały, gdy działano na nie bodźcami w y w o ł u j ą c y m i w korze mózgowej stan hamowani^. Według Pawłowa sen powstaje wskutek promieniowania hamowania, które obejmuje nie tylko całą korę mózgową, ale przechodzi też i na pień mózgu. Według teorii niedokrwiennej sen następuje wskutek zmniejszenia dopływu krwi do mózgu lub ośrodków świadomości. W ten sposób tłumaczy się senność po spożyciu posiłku, gdy duża ilość krwi gromadzi się w jamie brzusznej. Według teorii chemicznej we krwi mają się gromadzić „toksyny zmęczenia" lub inne produkty chemiczne wprowadzające ustrój w stan snu.
TWÓR SIATKOWATY Na całej przestrzeni pnia mózgowego, a więc w obrębie rdzenia przedłużonego, śródmózgowia, a częściowo także i międzymózgowia rozrzucone są dyfuzyjnie neurony tworzące jednorodną sieć komórek nazwaną układem s i a t k o w a t y m ( f o r m a t i o reticularis). Neurony te nie skupiają się w jądra, chociaż przenikają i otaczają jądra znajdujące się w pniu mózgowym. Układ siatkowaty składa się z małych i dużych neuronów przeważnie o krótkich aksonach. Wszystkie drogi nerwowe dośrodkowe i odśrodkowe przechodzą przez t w ó r siatkowaty, uzyskując z nim połączenia przez odgałęzienia włókien nerwowych. Ponadto neurony t w o r u siatkowatego mają bezpośrednie połączenia z korą mózgową i jądrami podkorowymi. W p ł y w układu siatkowatego może się rozprzestrzeniać w wielu kierunkach: „w d ó ł " - na rdzeń kręgowy i „w g ó r ę " - na struktury węchomózgowia, gdzie powstają afekty i emocje oraz dalej na korę mózgową, która kieruje procesami intelektualnymi, oraz „ n a boki" - przez podwzgórze i przysadkę na czynności narządów wewnętrznych i gruczołów dokrewnych. Twór siatkowaty pełni bardzo ważną, choć jeszcze nie we wszystkich szczegółach poznaną, funkcję: pod jego kontrolą pozostaje ogólny stopień '438
aktywności całego ośrodkowego układu nerwowego, włącznie ze stanem czuwania i snu oraz aktywności swoistych obszarów związanych z naszą świadomością. Porównać go można do elektrowni, która dostarcza prądu dla funkcjonowania sieci przenoszenia informacji - telefonów, telekomunikacji, radia itd. Wyłączenie tego układu, np. zniszczenie przez guz n o w o t w o r o w y , w y l e w lub proces zapalny, powoduje u człowieka stan śpiączkowy-całkowitą utratę zdolności reagowania na bodźce budzące - podobnie jak wyłączenie elektrowni uniemożliwia odbieranie sygnałów radiowych i nadawanie audycji z radiostacji. Drażnienie prądem elektrycznym tworu siatKowatego u śpiącego zwierzęcia powoduje jego natychmiastowe obudzenie. Z drugiej strony powstające w mózgu silne pobudzenie oddziałuje na układ siatkowaty, utrzymując się w nim przez dłuższy czas, nawet jeśli przyczyna wywołująca dawno znikła. Majaczenia i halucynacje mają źródło w takim patologicznym pobudzeniu układu siatkowatego.
NERWY CZASZKOWE N e r w y czaszkowe ( nn. craniales) w liczbie 12 par odchodzą symetrycznie od mózgu i opuszczają czaszkę przez otwory w jej podstawie. Poza nerwem X i XI unerwiają one głównie narząd głowy. Noszą one następujące nazwy: I. nn. w ę c h o w e (nn. olfactorii), II. n. wzrokowy (n. opticuś), III. n. okoruchowy (n. oculomotorius), IV. n. bloczkowy (n. trochlearis), V. n. trójdzielny (n. trigeminus), VI. n. odwodzący (n. abducens), VII. n. twarzowy (n. facialis), VIII. n. przedsionkowo-śl i makowy (n. vestibulocochlearis), IX. n. językowo-gardłowy (n. glossopharyngeus), X. n. błędny (n. vaguś), XI. n. dodatkowy ( n. accessorius), XII. n. podjęzykowy (n. hypoglossus).
Nerwy węchowe (I) N e r w y w ę c h o w e (nn. olfactorii) składają się z ok. 20 nici węchowych, które odchodzą od komórek węchowych, mieszczących się w błonie śluzowej okolicy węchowej jamy nosa. Nici węchowe wchodzą do jamy czaszki przez blaszkę sitową kości sitowej, by zakończyć się w opuszce węchowej. Przerwanie nici węchowych powoduje utratę powonienia. Zdarza się to niekiedy w następstwie urazów czaszki lub wskutek ucisku guza na nerwy węchowe. '439
N. wzrokowy (II) N. odwodzący (VI)
zwaczowy
Ryc. 200. Rozmieszczenie nerwów czaszkowych. Kolorem czerwonym oznaczono nerwy ruchowe, kolorem niebieskim - nerwy czuciowe.
'440
Ryc. 201. Nerw węchowy (I).
Nerw wzrokowy (II) N e r w w z r o k o w y (n. opticuś) tworzą włókna nerwowe, odchodzące od komórek zwojowych siatkówki. Nerw przebija zewnętrzną warstwę siatkówki, naczyniówkę i twardówkę i opuszcza gałkę oczną w miejscu położonym około 3 mm przyśrodkowo od bieguna tylnego gałki ocznej. Stąd kieruje się ku kanałowi wzrokowemu, przez który wchodzi do dołu czaszkowego środkowego. W odległości około 1 cm za gałką oczną nerwu wzrokowego wchodzi tętnica i żyła środkowa siatkówki (a. et v. centralis retinae). Po wejściu do jamy czaszki nerw wzrokowy kończy się w skrzyżowaniu w z r o k o w y m ( chiasma opticum), położonym na trzonie kości klinowej w rowku skrzyżowania n e r w ó w wzrokowych. Ze skrzyżowania wychodzi po każdej stronie pasmo wzrokowe ( tractus opticuś), doprowadzające włókna wzrokowe do poduszki wzgórza, ciała kolankowatego bocznego i wzgórków górnych blaszki pokrywy śródmózgowia. W skrzyżowaniu w z r o k o w y m tylko część (mniej więcej połowa) włókien nerwowych przechodzi na stronę przeciwną. Włókna idące od bocznej połowy siatkówki nie ulegają skrzyżowaniu, lecz przechodzą do pasma wzrokowego tej samej strony. Włókna biegnące od przyśrodkowej strony siatkówki przechodzą w obrębie skrzyżowania wzrokowego na stronę przeciwną. Każde pasmo wzrokowe ma zatem połączenie z obu gałkami ocznymi. Lewe pasmo wzrokowe zawiera zatem włókna z lewej połowy każdego oka, prawe z prawej. Przecięcie pasma wzrokowego po jednej stronie (np. lewej) powoduje '441
Ryc. 202. Nerw wzrokowy.
połowicze niedowidzenie w obu oczach (wypadnięcie prawej części pola widzenia w każdym oku). Choroby nerwu wzrokowego (zanik n. wzrokowego) prowadzą do upośledzenia i utraty wzroku.
Nerw okoruchowy (III) Nerw o k o r u c h o w y (n. oculomotorius) jest nerwem ruchowym gałki ocznej. Unerwia on mięsień dźwigacz powieki górnej oraz wszystkie zewnętrzne mięśnie oka, z wyjątkiem mięśni prostego bocznego i skośnego górnego. W skład nerwu okoruchowego wchodzą także włókna przywspółczulne, unerwiające zwieracz źrenicy. Uszkodzenie nerwu III pociąga za sobą porażenie wszystkich ruchów gałki ocznej z wyjątkiem odwodzenia i nikłego ruchu ku dołowi i do wewnątrz. Gałka oczna ustawia się w pozycji zwróconej w kierunku kąta zewnętrznego oka. Towarzyszy temu opadnięcie powieki górnej. Źrenica jest rozszerzona i nie reaguje na światło ani na zbieżność.
'442
Nerw bloczkowy (IV) N e r w bloczkowy (n. trochlearis) jest najcieńszym nerwem m ó z g o w y m i unerwia tylko jeden mięsień skośny górny oka. Wychodzi z pnia mózgu po stronie grzbietowej, tuż poniżej wzgórków dolnych blaszki pokrywy, po czym owija się wokół konara mózgowego i ukazuje się na podstawie mózgu tuż za pasmem wzrokowym. Dalej biegnie ku przodowi w bocznej ścianie zatoki jamistej ku szczelinie oczodołowej górnej, przez którą wchodzi do oczodołu i kończy się w mięśniu skośnym górnym. Porażenie nerwu IV powoduje występowanie zeza podczas spoglądania w górę; gałka oczna skręca się do wewnątrz.
Nerw trójdzielny (V) Nerw trójdzielny (n. trigeminus) jest najgrubszym nerwem mózgowym. Tworzą go włókna czuciowe, unerwiające powłoki i części głębokie twarzy oraz włókna ruchowe dla mięśni poruszających żuchwę. Wychodzi on z mózgu na granicy pomiędzy mostem a ramieniem móżdżkowym środkowym d w o m a korzeniami: przednim - r u c h o w y m ( p o r t i o minor) i t y l n y m - czuciowym (portio major). Przebija oponę twardą w pobliżu wierzchołka kości skalistej.
łowy dolny Ryc. 203. N e r w t r ó j d z i e l n y (V)
'443
gdzie część czuciowa tworzy duży zwój nerwu trójdzielnego (ganglion trigeminale), położony w wycisku zwoju nerwu trójdzielnego na części skalistej kości skroniowej. Od zwoju tego odchodzę trzy duże gałęzie: 1) nerw oczny (n. ophthalmicus), 2) nerw szczękowy (n. maxillaris), 3) nerw żuchwowy (n. mandibularis), do którego dołącza się w całości część ruchowa ( p o r t o minor). 1. N e r w oczny (n. ophthalmicus) biegnie ku przodowi w kierunku oczodołu, do którego wchodzi przez szczelinę oczodołową górną. Po drodze mija zatokę jamistą, przebiegając w jej ścianie bocznej. Oddaje tu gałązkę czuciową do opony twardej. W pobliżu szczeliny oczodołowej górnej dzieli się na trzy gałęzie: nerw czołowy, nerw nosowo-rzęskowy i ń e r w ł z o w y . 2. N e r w szczękowy [n. maxillaris) po wyjściu ze zwoju biegnie w bocznej ścianie zatoki jamistej w kierunku o t w o r u okrągłego, przez który wchodzi do dołu skrzydłowo-podniebiennego (fossa pterygopalatina), a stąd przez szczelinę oczodołową dolną do kanału podoczodołowego. Unerwia czuciowo zęby górnego łuku zębowego. 3. N e r w ż u c h w o w y (n. mandibularis) jest największą gałęzią nerwu trójdzielnego. Oprócz włókien czuciowych zawiera także włókna ruchowe, jest zatem nerwem mieszanym. Opuszcza jamę czaszki przez otwór owalny. Unerwia czuciowo zęby dolnego łuku zębowego.
Nerw odwodzący (VI) N e r w odwodzący (n. abducens) unerwia tylko jeden mięsień oka. Wychodzi z mózgu za tylną krawędzią mostu, kieruje się ku przodowi i ku górze. Po drodze przebija zatokę jamistą, przez którą przechodzi bocznie od tętnicy szyjnej wewnętrznej. Wchodzi do oczodołu przez szczelinę oczodołową górną, gdzie unerwia mięsień prosty boczny gałki ocznej (m. rectus lateralis). W przypadku porażenia nerwu VI gałka oczna jest skierowana do wewnątrz (zez zbieżny).
Nerw twarzowy (VII) N e r w t w a r z o w y (n. facialiś) jest nerwem mieszanym; składa się on z dwóch części: większej - ruchowej, i mniejszej - czuciowo-wydzielniczej, zwanej również n e r w e m pośrednim (n. intermedius). Nerw twarzowy wychodzi z rdzenia przedłużonego w kącie pomiędzy tylną krawędzią mostu a oliwką, w sąsiedztwie n e r w ó w pośredniego i statyczno-słuchowego. Razem z t y m i nerwami wchodzi do przewodu słuchowego wewnętrznego, a następnie biegnie we w ł a s n y m kanale kości skalistej i wychodzi na podstawę czaszki przez otwór rylcowo-sutkowy, ale już bez nerwu pośredniego, który odłącza się od niego jako n e r w skalisty większy (n. petrosus majoi) i struna bębenko'444
Ryc. 204. Nerw twarzowy (VII).
wa ( c h o r d a tympani). Po wyjściu z czaszki krótki pień nerwu twarzowego dzieli się wachlarzowato na szereg gałązek końcowych, które przebijają śliniankę przyuszna.i unąrwjają wszystkie mięśnie fnimiczne twarzy, mięsień szeroki szyi i brzusiec tylny mięśnia dwubrzuścowego. Struna bębenkowa ( c h o r d a tympani) odłącza się od nerwu twarzowego przed jego wyjściem z kanału kostnego i podąża do jamy bębenkowej, z której wychodzi przez szczelinę skalisto-bębenkową, by dołączyć się do nerwu językowego (p. wyżej). Porażenie o b w o d o w e n. VII pociąga za sobą porażenie wszystkich mięśni twarzy po stronie uszkodzenia, łącznie z mięśniem szerokim szyi. Twarz staje się niesymetryczna, oko po stronie uszkodzonej nie zamyka się. Niemożliwe jest zmarszczenie czoła.
Nerw przedsionkowo-ślimakowy (VIII) N e r w przedsionkowo-ślimakowy (n. vestibulocochlearis) jest nerwem czuciowym. Składa się z dwóch części - przedsionkowej ( pars yestibularis) i ślimakowej ( pars cochlearis). '445
{
Część przedsionkowa utworzona jest z włókien wychodzących z dwubiegunowych komórek z w o j u przedsionkowego ( g a n g l i o n vestibulare), pozostającego w związku z narządem równowagi - kanałami półkolistymi; część druga składa się z włókien odchodzących od z w o j u spiralnego (ganglion spirale), związanego z receptorem słuchowym - narządem spiralnym (Cortiego) (organum spirale). Nerw statyczno-słuchowy wychodzi z mózgu bocznie od nerwu twarzowego przy tylnej krawędzi mostu i kieruje się do przewodu słuchowego wewnętrznego, gdzie dzieli się na dwie części górną - przedsionkową i d o l n ą - ś l i m a k o w ą . Uszkodzenie części ślimakowej powoduje osłabienie słuchu lub głuchotę. Porażenie części przedsionkowej - zawroty głowy, nudności, w y m i o t y , zaburzenia równowagi.
Nerw językowo-gardłowy (IX) N e r w językowo-gardłowy (n. głossopharyngeus) jest nerwem mieszanym. Większą część nerwu tworzą włókna czuciowe, unerwiające gardło i język; niewielką część stanowią włókna ruchowe, przeznaczone dla mięśni gardła i wydzielnicze dla ślinianki przyusznej. Nerw ten odchodzi od rdzenia przedłużonego 5 - 6 korzonkami, które ukazują się w bruździe bocznej tylnej za oliwką. Opuszcza czaszkę przez otwór żyły szyjnej. W obrębie otworu tworzy z w ó j górny (ganglion superius), a po wyjściu z czaszki - z w ó j dolny (ganglion inferius), położony w dołku skalistym. Po wyjściu z czaszki biegnie pomiędzy żyłą szyjną wewnętrzną a tętnicą szyjną wewnętrzną, a następnie kieruje się ku nasadzie języka i bocznej ścianie gardła.
Nerw błędny (X) N e r w błędny (n. vagus) jest nerwem mieszanym. Większość jego włókien należy do części przywspółczulnej układu nerwowego autonomicznego. Nazwa jego wywodzi się stąd, że obszar jego unerwienia sięga daleko poza głowę. Nerw błędny unerwia opony mózgowe, narządy szyi, serce, narządy oddechowe i znaczną część przewodu pokarmowego. Odchodzi od rdzenia przedłużonego za pośrednictwem kilku korzonków. Opuszcza czaszkę przez otwór żyły szyjnej. W obrębie tego otworu część czuciowa nerwu tworzy niewielki z w ó j górny, a po wyjściu z o t w o r u - drugi, większy z w ó j dolny. Na szyi nerw błędny biegnie pomiędzy tętnicą szyjną wspólną a żyłą szyjną wewnętrzną we wspólnej pochewce powięziowej. Z szyi przechodzi do śródpiersia górnego, a następnie tylnego, przy czym nerw błędny lewy krzyżuje się od przodu z łukiem aorty, prawy - z tętnicą podobojczykową prawą. Dalej ku dołowi nerwy błędne (trunci vagales) przechodzą na ścianę przełyku, gdzie tworzą splot przełykowy. Pień błędny lewy stopniowo przemieszcza się na '446
przednią ścianę przełyku, prawy - na tylną. Po przejściu do jamy brzusznej pnie błędne rozgałęziają się w ścianie żołądka. Na przebiegu nerwu błędnego rozróżniamy część g ł o w o w ą , szyjną, piersiową i brzuszną. Zwój górny Zwój dolny Gałąź gardłowa N. krtaniowy górny T. szyjna wspólna N btodny (X)
N. błędny (X) 1 N. krtaniowy wsteczny Tchawica
-r——Oskrzele główne ; lewe
Splot płucny
_
Splot przełykowy
.Gałąź żołądkowa przednia
Gałęzie wdtiobnwu
Ryc. 205. Nerw błędny (X).
Nerw dodatkowy (XI) N e r w d o d a t k o w y (n. accessorius) jest nerwem ruchowym. Składa się z dwóch części: jedna jest utworzona z korzeni czaszkowych ( radices craniales), odchodzących od rdzenia przedłużonego, drugą tworzą korzenie rdzeniowe (radices spinałeś), odchodzące od górnej części rdzenia kręgowego. Korzenie rdzeniowe wchodzą do jamy czaszki przez otwór wielki kości potylicznej, gdzie łączą się z korzeniami czaszkowymi we wspólny pień. Po wyjściu z czaszki przez otwór żyły szyjnej nerw dzieli się na dwie gałęzie - wewnętrzną '447
i zewnętrzną. Pierwsza dołącza się w całości do nerwu błędnego, a włókna jej unerwiają wewnętrzne mięśnie krtani i mięśnie- zwieracze gardła. Gałąź zewnętrzna biegnie do mięśnia mostkowo-obojczykowo-sutkowego, przebija go i zdąża do mięśnia czworobocznego grzbietu. Unerwia ruchowo te dwa mięśnie.
N. trójdzielny
N. odwodzący Nn. twarzowyi słuchowy
N. językowo-garałowy N. błędny
Oliwka Piramida
Otwór szyjny Zwój górny n. błędnego
I. dodatkowy (XI) Ryc. 206. Nerw dodatkowy (XI).
Nerw podjęzykowy (XII) N e r w podjęzykowy (n. hypoglossus) jest nerwem ruchowym. Zaopatruje wszystkie mięśnie języka wewnętrzne i zewnętrzne, z wyjątkiem mięśnia podniebienno-językowego. Wychodzi z rdzenia przedłużonego kilkoma korzeniami w bruździe bocznej przedniej. Opuszcza czaszkę przez kanał nerwu podjęzykowego, po czym zatacza łuk, skierowany wypukłością ku dołowi i wchodzi do języka licznymi gałązkami językowymi. '448
Po drodze n. podjęzykowy tworzy zespolenie z nerwami szyjnymi pierwszym i d r u g i m tworząc na szyi tzw. pętlę szyjną (ansa cervicalis), od której odchodzą gałęzie ruchowe do mięśni podgnykowych.
NERWY RDZENIOWE N e r w rdzeniowy (n. spinalis) powstaje przez połączenie odchodzącego od rdzenia kręgowego ruchowego korzenia brzusznego (radix ventralis) z czuciow y m korzeniem g r z b i e t o w y m (radixdorsalis), związanym ze z w o j e m rdzeniow y m (ganglion spinale). Nerwy rdzeniowe są nerwami mieszanymi i unerwiają mięśnie i skórę. Układ nerwowy
Narząd efektorowy
Acetylocholina
M. szkieletowy
Zwój
Włókno przedzwojowe
Acetylocholina
/
<
Włókno zazwojowe wtó
Noradrenalina Acetylocholina \
Adrenalina Część rdzeniowa nadnerczy Zwój
Acetylocholina
Włókno przedzwojowe
K< fen
Włókno zazwojowe
Acetylocholina
Ryc. 207. Schemat o b w o d o w e j części o b w o d o w e g o układu n e r w o w e g o . W układzie somatyczn y m w y p u s t k i o s i o w e komórek ruchowych biegną bez przerwy do mięśni szkieletowych, gdzie wyzwala się substancja przekaźnikowa - acetylocholina. W układzie a u t o n o m i c z n y m droga pobudzenia z ośrodka do efektora składa się z d w ó c h n e u r o n ó w - przedzwojowego i zazwojowego. W układzie w s p ó ł c z u l n y m w ł ó k n a przedzwojowe są krótkie; w układzie przywspółczulnym zwoje są położone w pobliżu efektorów i włókna zazwojowe są krótkie. Na zakończeniach n e r w ó w współczulnych wyzwala się noradrenalina, włókien p r z y w s p ó ł c z u l n y c h - a c e t y l o c h o l i n a . Przekaźnikiem w zwojach (między w ł ó k n e m przedzwojowym i z a z w o j o w y m ) w o b u układach jest acetylocholina. 29 - Anatomia i fizjologia
449
W skład nerwu rdzeniowego wchodzą włókna czuciowe, ruchowe oraz autonomiczne. Wspólny pień nerwu rdzeniowego dzieli się na cztery gałęzie: 1) gałąź grzbietową (ramus dorsalis), 2) gałąź brzuszną (ramus ventralis), 3) gałąź o p o n o w ą ( ramus meningeus), 4) gałąź łączącą do pnia nerwu współczulnego (ramus communicans). Od rdzenia kręgowego odchodzi 31 par n e r w ó w rdzeniowych. W zależności od odcinka rdzeniowego, do którego te nerwy należą, rozróżniamy: 8 par n e r w ó w szyjnych (nn. cervicales) Ci-C 8 , 12 par piersiowych (nn. thoracici) Thi-Thi2, 5 par lędźwiowych (nn. lumbales) L-|-L 5 , 5 par n e r w ó w krzyżowych (nn. sacrales) S ^ S s i 1 nerw guziczny (n. coccygeus) Co.
Gałęzie grzbietowe nerwów rdzeniowych Gałęzie grzbietowe (rami dorsales) n e r w ó w rdzeniowych unerwiają głębokie mięśnie grzbietu oraz skórę całej okolicy grzbietowej, od potylicy aż do okolicy guzicznej. Każda z gałęzi grzbietowych dzieli się na gałąź przyśrodkową i gałąź boczną. Budowa i rozmieszczenie tych n e r w ó w na całym t u ł o w i u powtarzają się bez większych zmian. Do ważniejszych n e r w ó w odchodzących od gałęzi grzbietowych należą: n e r w potyliczny większy (n. occipitalis major), który jest gałęzią grzbietową drugiego nerwu szyjnego (C2), unerwiającą mięśnie i skórę okolicy potylicznej, n e r w y górne pośladków (nn. clunium superiores), odchodzące od gałązek grzbietowych ostatnich n e r w ó w lędźwiowych i dążące do skóry w okolicy pośladkowej, n e r w y środkowe pośladków (nn. clunium medii), odchodzące od n e r w ó w krzyżowych i również unerwiające okolicę pośladkową.
Gałęzie brzuszne n e r w ó w rdzeniowych Gałęzie brzuszne (rami ventrales) n e r w ó w rdzeniowych unerwiają mięśnie oraz skórę przedniej i bocznej okolicy szyi i tułowia, a także kończyn. Gałęzie brzuszne unerwiają znacznie większe okolice ciała, są też dużo grubsze od gałęzi grzbietowych. Najgrubsze są nerwy biegnące do kończyn. W odróżnieniu od gałęzi grzbietowych w nerwach odchodzących od gałęzi brzusznych okolic szyjnej, lędźwiowej, krzyżowej i guzicznej układ odcinkowy się zatarł. Zachował się on natomiast wyraźnie w obrębie klatki piersiowej. Gałęzie brzuszne wspomnianych wyżej odcinków rdzenia wymieniają ze sobą włókna nerwowe, tworząc w pobliżu kręgosłupa sploty (plexus). Pnie nerwowe odchodzące od splotów zawierają włókna pochodzące z szeregu sąsiednich segmentów rdzenia. '450
Splot szyjny Splot szyjny (plexus cervicalis) powstaje z zespolenia gałęzi brzusznych czterech pierwszych n e r w ó w szyjnych (C1-C4) i leży na wysokości czterech pierwszych kręgów szyjnych. Od splotu szyjnego odchodzą: 1. Krótkie gałęzie mięśniowe biegnące od mięśni szyi (pętla szyjna - p. wyżej). 2. N e r w p r z e p o n o w y (n. phrenicus) unerwia najważniejszy mięsień wdechowy - przeponę. Nerw przeponowy powstaje z włókien pochodzących z czwartego i trzeciego szyjnego segmentu rdzeniowego. Po wyjściu ze splotu nerw przeponowy biegnie po przedniej powierzchni mięśnia pochyłego przedniego pionowo w dół. W klatce piersiowej schodzi ku przeponie, biegnąc pomiędzy blaszką ścienną osierdzia i blaszkę ścienną opłucnej. Nerw przepon o w y oprócz włókien ruchowych zawiera także włókna czuciowe, którymi zaopatruje osierdzie oraz opłucną śródpiersiową i przeponową w najbliższym sąsiedztwie. Jednostronne porażenie nerwu przeponowego daje niewielkie objawy. Porażenie obustronne powoduje duszność wskutek ucisku przepony na płuca i wyłączenia jej udziału w oddychaniu. 3. N e r w y skórne, które unerwiają czuciowo skórę szyi i górne okolice klatki piersiowej, odchodzą od splotu szyjnego i ukazują się spod tylnej krawędzi mięśnia mostkowo-obojczykowo-sutkowego. Są t o : a) nerw potyliczny mniejszy (n. occipitalis minor), b) nerw uszny wielki (n. auricularis magnus), c) nerw poprzeczny szyi (n. transversus colli), d) nerwy nadobojczykowe (nn. supraclaviculares).
Splot ramienny Splot ramienny (plexus brachialis) tworzą gałęzie brzuszne czterech dolnych n e r w ó w szyjnych (C 5 -C 8 ) i większa część pierwszego nerwu piersiowego (Thi). Splot ten przeznaczony jest dla skóry i mięśni kończyny górnej. Każda z tych gałęzi dzieli się na dwie części - przednią i tylną, które łącząc się ze sobą tworzą trzy główne pęczki n e r w ó w długich unerwiających kończynę górną: pęczek boczny (fasciculus lateraliś), pęczek przyśrodkowy (fasciculus madialis) i pęczek tylny (fasciculus posterior). Pod względem topograficznym splot ramienny dzieli się na część nadobojczykową (pars supraclavicularis), położoną w dole nadobojczykowym, i część podobojczykową (pars infraclavicularis), leżącą poniżej obojczyka. Nerwy odchodzące od splotu ramiennego dzielimy na dwie grupy: gałęzie krótkie, unerwiające mięśnie okolicy barku, i gałęzie długie, unerwiające pozostałe części kończyny. Pierwsze odchodzą od części nadobojczykowej splotu, drugie - od części podobojczykowej. '451
~!Hp|jgi
Gałęzie długie powstają z trzech pęczków splotu ramiennego. Od pęczka bocznego odchodzi nerw mięśniowo-skórny i boczna część nerwu pośrodkowego, od pęczka przyśrodkowego - p'rzyśrodkowa część nerwu pośrodkowego, nerw łokciowy, nerw skórny ramienia przyśrodkowy i nerw skórny przedramienia przyśrodkowy. Z pęczka tylnego powstaje nerw promieniowy. 1. N e r w mięśniowo-skórny (n. musculocutaneus) po odejściu od splotu ramiennego kieruje się ku przedniej okolicy ramienia, przebijając mięsień kruczo-ramienriy, a następnie biegnie pomiędzy mięśniami ramiennym a d w u g ł o w y m ramienia. Unerwia ruchowo te trzy mięśnie. Po stronie bocznej ramienia kończy się gałęzią czuciową, zwaną nerwem przedramienia bocznym, unerwiającym skórę powierzchni bocznej przedramienia. Uszkodzenie tego nerwu powoduje porażenie wspomnianych trzech mięśni i osłabienie zginania przedramienia, które jest możliwe jeszcze tylko dzięki skurczowi mięśnia ramienno-promieniowego. '452
2. N e r w skórny przyśrodkowy ramienia (n. cutaneus brachii medialis) unerwia skórę dołu pachowego i przyśrodkowej powierzchni ramienia do kłykcia przyśrodkowego kości ramiennej. 3. N e r w skórny przyśrodkowy przedramienia (n. cutaneus antebrachii medialis) unerwia czuciowo skórę przedramienia po stronie łokciowej powierzchni przedniej. 4. N e r w p o ś r o d k o w y (n. medianus) jest nerwem mieszanym. Odchodzi od splotu za pośrednictwem d w u ramion obejmujących widełkowato tętnicę pachową. Na ramieniu biegnie najpierw razem z tętnicą ramienną we wspólnej pochewce łącznotkankowej, następnie się od niej nieco oddala. W dole ł o k c i o w y m nerw pośrodkowy leży pod rozcięgnem mięśnia d w u g ł o w e g o ramienia ( a p o n e u r o s i s m. bicipitis brachii). Na przedramieniu biegnie w kierunku dłoni pomiędzy warstwą głębokich a powierzchownych zginaczy palców. Na dłoń dostaje się przez kanał nadgarstkowy pod troczkiem zginaczy. W obrębie ramienia i stawu łokciowego nerw pośrodkowy nie oddaje żadnych odgałęzień. W górnej części przedrarriienia wysyła szereg gałęzi mięśniowych (rr. musculares) do wszystkich mięśni strony przedniej przedramienia, z wyjątkiem zginaczy nadgarstka łokciowego i części łokciowej zginacza palców głębokiego. Na dłoni nerw pośrodkowy rozgałęzia się ha trzy n e r w y d ł o n i o w e w s p ó l n e palców (nn. digitales palmares communes), do palców pierwszego, drugiego, trzeciego i strony promieniowej palca czwartego. 5. N e r w ł o k c i o w y (n. ulnaris) jest nerwem mieszanym; odchodzi od pęczka przyśrodkowego splotu ramiennego i biegnie po przyśrodkowej stronie tętnicy ramiennej. W okolicy łokcia odłącza się od tętnicy i okrąża od tyłu kłykieć przyśrodkowy kości ramiennej, biegnącw rowku nerwu łokciowego, gdzie jest pokryty tylko powięzią i skórą okolicy łokciowej i daje się łatwo wymacać;"stąd nerw łokciowy przechodzi na przednią powierzchnię przedramienia, gdzie biegnie w towarzystwie tętnicy łokciowej pomiędzy mięśniem zginaczem palców głębokim a mięśniem zginaczem łokciowym nadgarstka. W obrębie ramienia nerw łokciowy gałęzi nie oddaje. Na przedramieniu unerwia te mięśnie zginacze, które nie zostały unerwione przez nerw pośrodkowy, a więc zginacz łokciowy nadgarstka i połowę zginacza głębokiego palców. 6. N e r w p r o m i e n i o w y (n. radialis) stanowi przedłużenie pęczka tylnego splotu ramiennego, jest on najgrubszym nerwem splotu. Pień nerwu promieniowego po odłączeniu się od splotu ramiennego okrąża spiralnie od tyłu kość ramienną, biegnącw bruździe nerwu promieniowego, przykryty głową boczną mięśnia trójgłowego. W tej okolicy nerw promieniowy oddaje szereg gałęzi mięśniowych do mięśnia trójgłowego ramienia oraz gałąź czuciową - n e r w skórny t y l n y ramienia (n. cutaneus brachii posterior), unerwiający skórę pokrywającą głowę boczną tego mięśnia. Po wydostaniu się z obrębu mięśnia trójgłowego nerw p r o m i e n i o w y biegnie po stronie bocznej ramienia. W okolicy łokciowej znajduje się pomiędzy mięśniami ramienno-promieniowym a ramiennym. '453
Od pnia nerwu promieniowego oddziela się jeszcze w obrębie ramienia n e r w skórny t y l n y przedramienia (n. cutaneus antebrachii posterior), który przebija powięź ponad stawem łokciowym i unerwia skórę tylnej powierzchni przedramienia. W okolicy stawu łokciowego nerw promieniowy oddaje odgałęzienia do mięśnia ramienno-promieniowego, a następnie dzieli się na dwie gałęzie końcowe - głęboką i powierzchowną. Gałąź głęboka (r. profundus n. radialis) przechodzi ną stronę tylną przedramienia, gdzie unerwia wszystkie bez wyjątku prostowniki przedramienia i mięsień łokciowy. Gałąź powierzchowna (r. superficialis n. radialis) biegnie najpierw po stronie promieniowej przedramienia i przechodzi na grzbiet ręki, gdzie dzieli się na pięć n e r w ó w grzbietowych palców (nn. digitales dorsales), które unerwiają czuciowo stronę promieniową grzbietowej powierzchni ręki oraz częściowo grzbiet i zwrócone ku sobie powierzchnie palców pierwszego, drugiego i trzeciego. '454
Nerwy międzyżebrowe Gałęzie brzuszne n e r w ó w piersiowych, w liczbie 12 par, noszą nazwę n e r w ó w międzyżebrowych (nn. intercostales). Nerwy te nie tworzą splotów, lecz są ułożone odcinkowo. Jedynie pierwszy i ostatni nerw międzyżebrowy nawiązują łączność z sąsiednimi splotami. Nerwy międzyżebrowe unerwiają mięśnie ściany klatki piersiowej i j a m y brzusznej, blaszką ścienną opłucnej żebrowej i otrzewnej oraz skórę bocznej i przedniej powierzchni tułowia. Każdy z n e r w ó w międzyżebrowych biegnie razem z tętnicą i żyłą międzyżebrową w odpowiedniej przestrzeni międzyżebrowej, wzdłuż dolnej krawędzi żebra. Dolne nerwy międzyżebrowe wychodzą spod łuku żebrowego i dążą do mięśni brzucha. Nerwy międzyżebrowe oddają gałęzie mięśniowe, gałęzie skórne, gałęzie czuciowe głębokie do opłucnej i otrzewnej oraz gałęzie łączące z układem współczuł nym. Najgrubsze są gałęzie skórne. Od pnia n. międzyżebrowego odchodzą dwie gałęzie skórne: gałąź skórna boczna (r. cutaneus lateralis), wychodząca do skóry pomiędzy linią pachową a sutkową, oraz gałąź skórna przednia (r. cutaneus anterior), która wychodzi do skóry w pobliżu linii przymostkowej. Stanowią one liejako zakończenie nerwów międzyżebrowych. Od gałęzi skórnych II, III i IV nerów międzyżebrowych odchodzą gałęzie od gruczołów sutkowych. Również od n e r w ó w II i III odchodzą gałęzie skórne do kończyny górnej, noszące nazwę n e r w ó w międzyżebrowo-ramiennych (nn. intercostobrachiales).
Splot lędźwiowy Splot l ę d ź w i o w y (plexus lumbalis) jest utworzony przez gałęzie brzuszne I, II, III i częściowo IV nerwu lędźwiowego, jak również drobną gałąź od nerwu podżebrowego (Thi2>. Gałęzie splotu łędźwiowego unerwiają część mięśni i skóry jamy brzusznej, mięśnie i skórę przyśrodkowej i przedniej powierzchni uda oraz przyśrodkową powierzchnię podudzia. Od splotu lędźwiowego odchodzą: a. Krótkie gałęzie mięśniowe, unerwiające mięsień czworoboczny lędźwi, mięsień biodrowo-lędźwiowy i mięsień lędźwiowy mniejszy. b. Gałęzie długie: 1) n e r w biodrowo-podbrzuszny, 2) n e r w biodrowo-pac h w i n o w y , 3) n e r w płciowo-udowy, 4) n e r w skórny uda boczny, 5) n e r w u d o w y , 6) n e r w zasłonowy. N e r w u d o w y (n. femoralis) jest najgrubszym nerwem splotu lędźwiowego. Po wyjściu ze splotu biegnie ku dołowi pomiędzy mięśniem lędźwiowym większym a mięśniem b i o d r o w y m . Wychodzi na udo pod więzadłem pachwin o w y m przez rozwór mięśniowy. Około 3 cm poniżej więzadła pachwinowego pień nerwu udowego rozpada się na szereg gałęzi. Zespół tych gałęzi można podzielić na grupę powierzchowną, przeznaczoną dla skóry, i głębszą - dla mięśni przedniej powierzchni uda. '455
Gałęzie ruchowe odchodzą od nerwu udowego w miednicy małej albo po wyjściu na udo i unerwiają mięśnie: biodrowy, lędźwiowy większy, krawiecki i czworogłowy uda. Porażenie nerwu udowego uniemożliwia prostowanie kończyny dolnej w stawie kolanowym wskutek porażenia mięśnia czworogłowego uda (niemożność wchodzenia po schodach). N e r w zasłonowy (n. obturatorius) po odejściu od splotu zdąża wraz z tętnicą i żyłą zasłonową w kierunku o t w o r u zasłonionego i kanału zasłonowego, przez który opuszcza miednicę. Przed wejściem do kanału lub w s a m y m kanale nerw. ten wysyła gałąź ruchową do mięśnia zasłonowego wewnętrznego. Po wyjściu z kanału albo jeszcze w kanale dzieli się na dwie gałęzie - przednią i tylną, które unerwiają skórę powierzchni przyśrodkowej uda oraz grupę mięśni przywodzicieli.
Splot krzyżowy Splot krzyżowy ( plexus sacralis) powstaje z gałęzi brzusznych IV i V nerwu lędźwiowego i wszystkich n e r w ó w krzyżowych. Pnie nerwowe tworzące splot odznaczają się szczególną grubością, gdyż włókna ich unerwiają znaczną część kończyny dolnej. 4 - 5 pni nerwowych po wyjściu z kanału krzyżowego zespala się, tworząc trójkątną blaszkę, skierowaną wierzchołkiem ku o t w o r o w i kulszow e m u większemu. Przedłużeniem tej blaszki jest nerw k u I s z o w y - największy nerw splotu krzyżowego i całego organizmu ludzkiego. Od splotu krzyżowego odchodzą: 1. Krótkie gałęzie do mięśnia zasłonowego zewnętrznego, gruszkowatego, mięśni bliźniaczych i mięśnia czworobocznego uda. 2. N e r w pośladkowy górny (n. gluteus superior), który wychodzi z miednicy małej razem z tętnicą pośladkową górną przez górną część o t w o r u kulszow e g o większego (otwór nadgruszkowaty) i unerwia ruchowo mięśnie pośladkowe średni i mały oraz mięsień naprężacz powięzi szerokiej. 3. N e r w pośladkowy dolny (n. gluteus interior) wychodzi przez dolną część otworu kulszowego większego (otwór podgruszkowy) razem z tętnicą pośladkową dolną i rozgałęziając się wachlarzowato unerwia mięsień pośladkowy wielki. 4. N e r w kulszowy (n. ischiadicus) jest najdłuższym i najgrubszym nerwem organizmu ludzkiego. Utworzony jest przez włókna pochodzące od wszystkich gałęzi tworzących splot krzyżowy. Opuszcza miednicę przez o t w ó r podgruszkowy, biegnie pomiędzy guzem kulszowym a krętarzem wielkim, przykryty mięśniem pośladkowym wielkim, a następnie zdąża po tylnej stronie uda do dołu podkolanowego, biegnąc pomiędzy mięśniem przywodzicielem uda wielkim a grupą zginaczy. Zwykle w kącie g ó r n y m dołu podkolanowego dzieli się na dwa główne pnie: grubszy, przyśrodkowy - nerw piszczelowy, i cieńszy, boczny - nerw strzałkowy wspólny. Na udzie odchodzą od pnia kulszowego gałęzie ruchowe do '456
w Splot lędźwiowy -
Splot krzyżowy-
-Splot krzyżowy
iiiii — N. kulszowy
i
• N. udowy -
M. piszczelowy M. strzałkowy wspólny
N. strzałkowy powierzchowny^ N. strzałkowy głęboki
N. udowo-goleniowy
-N. łydkowy
Ryc. 210. Nerwy kończyny dolnej.
,
,
mięśni półbłoniastego, półścięgnistego, d w u g ł o w e g o uda i przywodziciela wielkiego. N e r w piszczelowy (n. tibialis) po oddzieleniu się od pnia nerwu kulszowego biegnie przez środek dołu podkolanowego, gdzie przechodzi pomiędzy dwiema g ł o w a m i mięśnia brzuchatego łydki i wnika pod mięsień płaszczkowaty, kierując się pod nim wzdłuż całego podudzia w stronę guza piętowego. N e r w strzałkowy w s p ó l n y (n. peroneus communis) po oddzieleniu się od pnia nerwu kulszowego kieruje się ukośnie ku dołowi i bokowi, owija się dookoła g ł o w y kości strzałkowej i przebija mięsień strzałkowy długi, w którym dzieli się na dwie gałęzie - nerwy strzałkowe: głęboki i powierzchowny. Jeszcze w dole podkolanowym od nerwu strzałkowego w s p ó l n e g o odchodzi n e r w skórny boczny łydki (n. cutaneus surae lateralis), który unerwia skórę bocznej powierzchni goleni. 457
i
i
-
N e r w strzałkowy p o w i e r z c h o w n y (n. peroneus superficialis) biegnie po oddzieleniu się od nerwu strzałkowego głębokiego pomiędzy mięśniami strzałkowymi długim a krótkim. Unerwia obydwa te mięśnie. N e r w strzałkowy głęboki (n. peroneus profundus) przebiega po stronie przedniej goleni wspólnie z tętnicę piszczelową przednią, z którą schodzi na stopę. Unerwia na goleni mięsień piszczelowy przedni, prostownik długi palców, prostownik długi palucha.
Splot sromowy Odróżniany przez niektórych autorów splot s r o m o w y (plexus pudendus), będący właściwie częścią splotu krzyżowego, utworzony jest przez gałęzie brzuszne III i IV n e r w ó w krzyżowych. Splot ten tworzy źespolenie ze splotami sąsiednimi. Gałęzie mięśniowe splotu sromowego unerwiają dźwigacz odbytu i mięsień guziczny. Gałęzie trzewne, w liczbie 4 - 5 , razem ze splotem współczulnym biorą udział w unerwieniu narządów miednicy małej.
Splot guziczny Splot guziczny ( p l e x u s coccygeus) jest utworzony przez nerw guziczny i gałąź przednią V nerwu krzyżowego. Oddaje on gałęzie unerwiające czuciowo okolicę odbytu i kości guzicznej.
UKŁAD NERWOWY AUTONOMICZNY Na wstępie wspomnieliśmy, że układ n e r w o w y za pośrednictwem włókien odśrodkowych wykonuje dwojakiego rodzaju czynności: 1) powoduje skurcze mięśni szkieletowych i 2) reguluje czynności życiowe samego organizmu i przemianę materii. Ta część układu nerwowego, od której zależą czynności mięśni poprzecznie prążkowanych, nazywa się układem somatycznym. Druga część, od której idą na o b w ó d włókna odśrodkowe do narządów wewnętrznych i która reguluje czynności serca, naczyń krwionośnych, gruczołów oraz mięśni gładkich w całym ustroju nazywa się układem n e r w o w y m autonomicznym (systema nervosum autonomicum). Nazwa pochodzi stąd, że zwoje tego układu są położone poza układem n e r w o w y m ośrodkowym, jak również dlatego, że szereg narządów unerwionych przez ten układ po usunięciu z ustroju i umieszczeniu w płynie fizjologiczn y m zachowuje przez pewien czas samoistną rytmiczną aktywność, oraz dlatego, że układ ten zawiaduje czynnościami niezależnymi od naszej woli. Układ n e r w o w y autonomiczny różni się od układu somatycznego zarówno pod względem budowy, jak i czynności. Grupy komórek nerwowych, od których odchodzą o b w o d o w e włókna nerwowe autonomiczne, są rozmieszczone w ośrodkowym układzie nerwo'458
N. okoruchowy Zwój rzęskowy ^ — Z w ó j klinowo-podniebienny Struna bębenkowa Zwój podżuchwowy i. błędny
Odcinek szyjny rdzenia Gałąź łącząca biała
Gałąź łącząca biała Gałąź łącząca szara
Odcinek i lędźwiowy "I
Odcinek I krzyżowy \
i •V
.
V
>
' +
;
/
I
L
Splot pęcherzowy
Ryc. 211. Schemat układu nerwowego wegetatywnego. Kolorem czerwonym oznaczono układ współczulny, żółtym - układ przywspółczulny, fioletowym - ośrodkową część układu.
'459
w y m nierównomiernie. Są one skupione w czterech okolicach ośrodkowego układu nerwowego, a mianowicie: 1) w ś r ó d m ó z g o w i u , 2) w rdzeniu przedłużonym, 3) odcinku piersiowo-lędźwiowym rdzenia kręgowego (na przestrzeni od I kręgu odcinka piersiowego do II lędźwiowego), 4) w odcinku krzyżowym rdzenia kręgowego. Ośrodki autonomiczne, położone w śródmózgowiu, rdzeniu przedłużonym i części krzyżowej rdzenia kręgowego wraz z odchodzącymi od nich nerwami stanowią część przywspółczulną (parasympatyczną) układu autonomicznego. Ośrodki autonomiczne części piersiowo-lędźwiowej rdzenia oraz odchodzące od nich włókna tworzą część współczułną (sympatyczną) układu nerwowego autonomicznego. Istotną cechą odróżniającą układ autonomiczny od somatycznego jest to, że droga nerwowa odśrodkowa w układzie autonomicznym składa się zawsze z d w ó c h neuronów. Włókna nerwowe somatyczne odchodzące od komórek rogów przednich rdzenia kręgowego biegną bez żadnej przerwy do mięśnia szkieletowego, natomiast każde włókno wychodzące z ośrodka autonomicznego w drodze na o b w ó d ulega przerwaniu w j e d n y m ze z w o j ó w o b w o d o w y c h , w których rozpoczyna się drugi neuron, doprowadzający swe wypustki do komórek efektora. Innymi s ł o w y każdy nerw autonomiczny (współczulny lub przywspółczulny) składa się z wypustek dwóch połączonych ze sobą neuronów, z których jeden leży w układzie n e r w o w y m ośrodkowym i kończy się w zwoju o b w o d o w y m , drugi w t y m zwoju się rozpoczyna i kończy w tkankach unerwianego narządu. Łuk odruchowy
Neuron dośrodkowy
Neuron dośrodkowy
Neuron pośredni
mammmmmm Neuron przedzwojowy
Neuron ruchowy
•
Neuron zazwojowy
Ryc. 212. Schematyczne porównanie przebiegu neuronów w łuku odruchowym somatycznym (niebieski) i wegetatywnym (czerwony).
'460
Włókna nerwowe autonomiczne, wychodzące z ośrodkowego układu nerw o w e g o i kończące się na komórkach położonych w zwoju o b w o d o w y m , nazywają się w ł ó k n a m i p r z e d z w o j o w y m i , natomiast włókna odchodzące od zwoju noszą nazwę w ł ó k i e n zazwojowych. Włókna nerwowe autonomiczne sącieńsze (2-7 firn) niż włókna somatyczne (12-14 (im) i są pozbawione całkowicie osłonki mielinowej lub mają osłonkę bardzo cienką. Włókna autonomiczne różnią się także od somatycznych pod względem czynnościowym. Pobudliwość ich jest znacznie niższa. Dla podrażnienia nerwu autonomicznego potrzebny jest znacznie silniejszy bodziec. Włókna autonomiczne przewodzą pobudzenie dużo wolniej (1-3 m/s) niż somatyczne (70-120 m/s). Inną cechą odróżniającą układ n e r w o w y autonomiczny od somatycznego jest t o , że układ autonomiczny nie ma własnych d r ó g dośrodkowych, tj. własnych włókien czuciowych. Ponieważ jednak czynności tego układu mają charakter odruchowy, potrzebny im jest do rozpoczęcia czynności bodziec czuciowy. Bodźce takie w odruchach autonomicznych są przekazywane przez włókna czuciowe, należące do układu somatycznego.
Zwoje autonomiczne W układzie autonomicznym skupienia komórek nerwowych, leżące na zewnątrz ośrodkowego układu nerwowego, czyli z w o j e autonomiczne, leżą na drodze przebiegu n e r w ó w autonomicznych w różnych odległościach od układu nerwowego ośrodkowego. Zależnie od ich położenia rozróżniamy: 1) z w o j e przykręgowe, 2) zwoje przedkręgowe i 3) z w o j e śródścienne. 1. Z w o j e przykręgowe należą do układu współczułnego. Leżą one po obu stronach kręgosłupa, tworząc pień współczulny ( truncus sympathicus). 2. Z w o j e przedkręgowe, które nieraz są znacznie bardziej oddalone od kręgosłupa, znajdują się jednak jeszcze dość daleko od unerwionych narządów. Leżą one na tylnej ścianie jamy brzusznej pozaotrzewnowo, w sąsiedztwie gałęzi aorty brzusznej. Zwoje te powstały wskutek wywędrowania na o b w ó d komórek nerwowych z ośrodkowego układu nerwowego w okresie embrionalnym. Do z w o j ó w przedkręgowych dochodzi stosunkowo niewiele włókien przedzwojowych, natomiast liczba włókien zazwojowych opuszczająca zwój jest bardzo duża. W zwojach przedkręgowych następuje zatem zwielokrotnienie włókien nerwowych dążących do narządów wewnętrznych. Włókna zazwojowe tworzą zawiłe sploty. Do splotów i z w o j ó w przedkręgow y c h należą: splot t r z e w n y {plexus celiacus), zawierający szereg zwojów, z których największym jest parzysty z w ó j t r z e w n y ( g a n g l i o n celiacum), położony w sąsiedztwie pnia trzewnego, splot i zwój nerkowy, splot krezkowy górny i dolny, splot nasienny, splot tętnicy głównej, splot podbrzuszny. 3. Trzecią grupę stanowią zwoje śródścienne, położone na powierzchni lub w ścianie narządów wewnętrznych. Zwoje i sploty tego rodzaju występują '461
w mięśniu sercowym, gruczołach, w ścianie przełyku, żołądka i jelit, w pęcherzu moczowym, macicy itd. Do komórek tych z w o j ó w dochodzą przeważnie przedzwojowe w ł ó k n a przywspółczulne.
Część współczulna układu autonomicznego Współczulne włókna nerwowe wychodzą z rdzenia kręgowego w składzie korzonków brzusznych w odcinku piersiowo-lędźwiowym. Komórki macierzyste tych włókien są skupione w rdzeniu kręgowym pomiędzy rogami przednimi a t y l n y m i istoty szarej rdzenia, tworząc na przestrzeni od pierwszego odcinka piersiowego do drugiego lędźwiowego słup boczny (columna lateralis).
Zwoje współczulne Zwój gwiazdzisty I zwój oiersiowy
N. trzewny większy T 5, 6, 7, 8, 9
N. trzewny mniejszy T 10,
Splot i zwój trzewny Splot i zwój krezkowy górny
Splot krezkowy dolny
"—Splot podbrzuszny
Ryc. 213. Układ nerwowy współczulny (wg Jacoba i Falcone'a).
'462
Po wyjściu z o t w o r u międzykręgowego wraz z nerwem rdzeniowym włókna współczulne odłączają się od niego, tworząc tzw. gałąź łączącą białą ( r a m u s communicans albus), która wchodzi do zwoju pnia współczułnego. Pień współczulny ( truncus sympathicus) z każdej strony tworzy 21 do 24 z w o j ó w współczułnych, połączonych ze sobą włóknami nerwowymi. Leży on na bocznej powierzchni kręgosłupa. W skład jego wchodzą 3 zwoje szyjne, 11-12 zwojów piersiowych, 4 - 5 zwojów lędźwiowych i 3 - 4 zwoje krzyżowe. Włókna nerwowe, które wchodzą do pnia nerwu współczulnego przez gałąź łączącą białą, biegną w n i m ku górze i ku dołowi i kończą się wokół komórek nerwowych, bądź to pierwszego napotkanego zwoju, bądź dopiero drugiego lub trzeciego. Są to włókna przedzwojowe. Od komórek zwojowych odchodzą włókna zazwojowe. Większość tych włókien dołącza się do n e r w ó w rdzeniowych, do których dochodzi przez gałęzie łączące szare (ramicommunicantes grisei). Wraz z nerwami rdzeniowymi włókna te biegną do unerwianych narządów. Część włókien zazwojowych dołącza się do naczyń tętniczych, z którymi dochodzi do narządów. Pień współczulny zawiera w odcinku szyjnym trzy zwoje: górny, ś r o d k o w y i dolny, który najczęściej łączy się ze zwojem piersiowym pierwszym, tworząc z w ó j szyjno-piersiowy albo gwiaździsty. Od zwoju górnego odchodzą włókna unerwiające narząd wzroku, gruczoły łzowe, ślinowe i serce. Zwoje środkowy i gwiaździsty oddają gałązki do splotu płucnego i sercowego. Od części piersiowej pnia współczulnego odchodzą dwa n e r w y t r z e w n e (nn. splanchnici) - większy i mniejszy. Nerwy te przebijają przeponę i kończą się w zwojach przedkręgowych splotu trzewnego. Pobudzenie włókien współczulnych idących do narządu wzrokowego powoduje rozszerzenie się źrenicy i szpary ocznej, wskutek czego oczy wydają się większe („oczy gwiazdy filmowej"). Przecięcie włókien współczulnych idących do oka w y w o ł u j e wystąpienie tzw. zespołu Homera, tj. zwężenia źrenicy, opadnięcia powieki górnej i zwężenia szpary powiekowej. W skórze pobudzenie ukłądu współczulnego powoduje wystąpienie „gęsiej skórki" polegające na skurczu mięśni gładkich przywłosowych wywołujących erekcję włosów. Widać to wyraźnie u zwierząt w postaci najeżonych w ł o s ó w na grzbiecie podczas silnego pobudzenia. W narządach wewnętrznych układ współczulny wzmaga skurcze mięśni gładkich, zwiększa częstość skurczów serca, pobudliwość i przewodnictwo. W płucach następuje rozszerzenie oskrzeli i zwężenie tętnic.
Część przywspółczulna układu autonomicznego Włókna nerwowe układu przywspółczulnego, jak już wspominaliśmy, opuszczają ośrodkowy układ nerwowy w obrębie czaszki i odcinka krzyżowego rdzenia kręgowego. Podobnie jak w układzie współczulnym, tak i tu rozróżniamy włókna przedzwojowe i zazwojowe. Zwoje współczulne jednak leżą w pobliżu rdzenia kręgowego, a ich włókna przedzwojowe są krótkie, natomiast w układzie przywspółczulnym włókna przedzwojowe są długie, a zwoje leżą '463
daleko od ośrodków, w pobliżu unerwianych narządów lub w ścianie tych narządów. Część czaszkowa układu przywspółczulnego pozostaje w związku z III, VII, IX i X nerwami czaszkowymi. Włókna przywspółczulne, idące z nerwem III (okoruchowym), zdążają do z w o j u rzęskowego ( g a n g l i o n ciliare), położonego w oczodole. Od zwoju tego odchodzą włókna zazwojowe do mięśnia rzęskowego i mięśnia zwężającego źrenicę. Włókna przedzwojowe, biegnące z nerwem VII (twarzowym), dochodzą do z w o j u skrzydłowo-podniebiennego ( g a n g l i o n pterygopalatinum), położonego w dole o tej samej nazwie oraz do z w o j u p o d ż u c h w o w e g o (ganglion submandibulare). Włókna zazwojowe, wychodzące ze zwoju skrzydłowo-podniebiennego, unerwiają gruczoł łzowy, błonę śluzową nosa i podniebienie. Od zwoju podżuchwowego włókna przywspółczulne dochodzą do ślinianek podżuchwowej i podjęzykowej. Włókna przedzwojowe, idące z nerwem IX (językowo-gardłowym), kończą się w z w o j u usznym (ganglion oticum), który leży pod o t w o r e m owalnym. Włókna zazwojowe unerwiają śliniankę przyuszną. Największe skupienie przedzwojowych włókien przywspółczulnych w odcinku czaszkowym stanowi n e r w błędny. Włókna te dochodzą w klatce piersiowej do s p l o t ó w sercowego i płucnego i kończą się w zwojach przywspółczu lnych tych narządów. W jamie brzusznej przedzwojowe włókna nerwu błędnego dochodzą do z w o j ó w przełyku, żołądka, w ą t r o b y , trzustki, jelita cienkiego i nerek. Nawiązują one łączność ze splotem n e r w o w y m błony mięśniowej i podśluzowym, zawartymi w ścianie przewodu pokarmowego na całej jego długości od przełyku do odbytu. Włókna przywspółczulne odcinka krzyżowego wychodzą z rdzenia wespół z korzonkami brzusznymi ll-IV n e r w ó w krzyżowych jako n e r w miedniczny (n. pelvicus), który dochodzi do splotu miednicznego. Włókna te unerwiają odbytnicę oraz wszystkie narządy miednicy małej.
Autonomiczne unerwienie narządów i tkanek Układ n e r w o w y autonomiczny doprowadza do unerwianych narządów bodźce, które zmieniają - osłabiają lub wzmacniają - przemianę materii, napięcie oraz czynności automatyczne tkanek i narządów. Większość narząd ó w pozostaje pod kontrolą o b y d w u części układu autonomicznego - współczulnego i przywspółczulnego. W p ł y w n e r w ó w współczulnych i przywspółczulnych na unerwione narządy jest niejednakowy, a bardzo często przeciwstawny. W wielu przypadkach nerwy współczulne działają pobudzająco na czynności narządów, gdy tymczasem nerwy przywspółczulne czynności te hamują i odwrotnie. Taki czynnościowy antagonizm n e r w ó w współczulnych i przywspółczulnych obserwuje się wyraźnie w wielu narządach. Na przykład pobudzenie '464
układu współczulnego powoduje rozszerzenie źrenicy, pobudzenie układu przywspółczulnego pociąga za sobą jej zwężenie. Rytm skurczów serca przyspiesza się przy pobudzeniu nerwów współczulnych i zwalnia wskutek pobudzenia nerwów błędnych (przywspółczulnych). Mięśnie oskrzeli kurczą się pod w p ł y w e m nerwu błędnego i rozluźniają przy pobudzeniu nerwów współczulnych. Układ przywspółczulny (n. błędny) wzmacnia skurcze żołądka i jelit, nerwy współczulne - osłabiają.
30 - Anatomia i fizjologia
UKŁAD NARZĄDÓW ZMYSŁÓW
W ciągu życia organizm ludzki jest w stałym kontakcie z otoczeniem, które wywiera znaczny w p ł y w na przebieg czynności życiowych. Niektóre czynniki wywołują odruchy obronne. W czasie upałów gruczoły potowe zwiększają ilość wydzieliny, która ochładza organizm i zabezpiecza go przed przegrzaniem. Przy jaskrawym świetle bezwiednie przymrużamy oczy, natomiast przy słabym oświetleniu źrenice się rozszerzają, czego zupełnie nie odczuwamy. Aby należycie reagować na otoczenie, człowiek musi się z nim dokładnie zapoznać. Do tego-celu służą mu narządy zmysłów, które syngalizują odbierane z otoczenia wrażenia. Powonienie informuje go o składzie powietrza, jak również o jakości przyjmowanych pokarmów, wzrok koryguje kierunek ruchu itp. Niektóre zmysły rozpoznają te same szczegóły otoczenia. Na przykład wzrok pozwala nam przy dostatecznym oświetleniu określić kształt czy wielkość przedmiotu oglądanego, natomiast przy braku oświetlenia, np. w ciemni, możemy poznać te cechy dotykiem, co w pewnych okolicznościach może mieć pierwszorzędne znaczenie. W niektórych przypadkach ten sam narząd jest siedliskiem kilku zmysłów, np. za pomocą języka rozpoznajemy smak pokarmu, jego konsystencję, temperaturę, ciała obce, które do niego mogą trafić itp. Z otoczenia odbieramy bardzo różnorodne wrażenia dzięki temu, że zakończenia nerwowe w poszczególnych narządach zmysłów mają rozmaitą budowę, która pozwala im na odbieranie jednego tylko rodzaju podniet. Jak wspomniano, narząd może reagować na różne podniety, ale zawsze w podobny sposób. Oko odbiera różne' bodźce, ale odczuwa je zawsze jako wrażenie światła, zarówno przy działaniu promienia słonecznego, jak i przy urazie fizycznym. Ucho odbiera tylko fale dźwiękowe i jest niewrażliwe, np. na barwy. Dotykiem czy węchem nie potrafimy odróżnić roztworu soli ód roztworu chininy, a zmysłem smaku nie jesteśmy w stanie np. wyczuć domieszki gazu świetlnego w powietrzu. Położenie narządów zmysłów w organizmie jest związane z ich czynnościami. Umieszczenie oczu na głowie, jako najwyższej części ciała, rozszerza zasięg wzroku, a położenie ich na twarzy pozwala na rozpoznawanie drogi przy najczęstszym posuwaniu się ciała do przodu. Uszy położone na powierzchniach bocznych głowy pozwalają na odbieranie fal głosowych ze wszystkich kierunków, a zwłaszcza z obu stron osobnika stojącego czy idącego. Narząd powonienia leży na początku dróg oddechowych i sygnalizuje ewentualne zanieczyszczenia powietrza lub produktów pokarmowych, jeszcze przed ich pobraniem. Również położenie języka, jako narządu smaku, na początku drogi '466
pokarmowej pozwala na ocenę pokarmu, jeszcze nie pobranego do przewodu trawiennego, a więc jego rodzaju, stopnia świeżości itd. Poza t y m sąsiedztwo narządu powonienia pozwala na ocenę pewnych wartości smakowych pożywienia, chociaż w życiu codziennym m ó w i się o roli podniebienia, co jedynie w m a ł y m stopniu odpowiada rzeczywistości, jak to dalej wyjaśnimy. Czynnikiem odbierającym bezpośrednio podniety, dochodzące do narząd ó w zmysłów, jest nabłonek nerwowy (neuroepithelium). W niektórych narządach zmysłów odbiera on podniety przy bezpośrednim zetknięciu się z ich źródłem. Takie stosunki spotykamy w niższych narządach zmysłów, jak smak czy powonienie. Natomiast wyżej zorganjzowane narządy, jak wzrok i słuch, wymagają dodatkowych urządzeń, położonych na drodze promieni światła lub fal dźwiękowych do zakończeń nerwowych, leżących w głębi oka czy ucha. Urządzenia te przepuszczają jedynie bodźce, które działają na ten narząd, przetwarzając je we właściwy sobie sposób, jak to dzieje się np. w gałce ocznej przy załamywaniu promieni światła wpadających do wnętrza gałki. Ponieważ każdy narząd zmysłu służy do celów kontaktu organizmu ze światem zewnętrznym i pozwala na poznawanie jego cech, wszystkie te narządy, chociaż bardzo różniące się od siebie, u j m u j e m y w jeden układ narządów z m y s ł ó w (systema organorum sensuum). Poza zmysłami m a m y jeszcze szereg czuć, jak np. odczuwanie ciężaru, odbierane przez czucie mięśniowe i stawowe, lub odczuwanie charakteru powierzchni ciał (gładkość, twardość itd.) przez czucie dotyku i in. Zawdzięczamy je różnie zbudowanym zakończeniom n e r w ó w czuciowych. Jedne z nich leżą w skórze - i te zaliczamy do narządów czucia powierzchniowego. Natomiast położone w narządach w głębi ciała zaliczamy do narządów czucia głębokiego.
NARZĄDY CZUCIA POWIERZCHNIOWEGO I GŁĘBOKIEGO Narządy czucia powierzchniowego występują w postaci tzw. ciałek krańcow y c h , rozmieszczonych w skórze. Zbudowane w różny sposób, ciałka te odbierają wrażenia dotykowe, ciepła, zimna, bólu, pieczenia, swędzenia itp. Głębiej pod skórą leżą narządy czucia głębokiego, które mają nieraz budowę podobną do ciałek krańcowych czucia powierzchniowego. W torebkach staw o w y c h i mięśniach leżą liczne zakończenia nerwowe, wspomagające nieraz czucie powierzchniowe, które trudno oddzielić od głębokiego, np. przy czynnościach ręki, wykonywanych w celu zbadania otoczenia dotykiem. Zakończenia czuciowe w mięśniach są też niezbędnym warunkiem występowania odruchów mięśniowych. Ciałka te spotykamy również we wszystkich tworach łącznotkankowych, jak okostna, omięsna, więzadła, powięzie. One właśnie powodują odczuwanie bólu przy urazach czy stanach zapalnych wymienionych narządów. Dzięki nim również oceniamy np. kształt ujmowanego ręką przedmiotu, jego ciężar, twardość, elastyczność, odporność itp. '467
NARZĄD S M A K U Przy opisie j a m y ustnej była mowa o brodawkach błony śluzowej języka. Rozróżniamy na niej brodawki stożkowate, nitkowate orazgrzybowate, okolone i liściaste. Pierwsze spełniają czynności mechaniczne, drugie są siedliskiem narządu smaku (organon gustus) dzięki rozmieszczonym w nich kubkom smakowym. Kubki te znajdują się również w otoczeniu cieśni gardzieli - na podniebieniu miękkim, na nagłośni i na ścianie tylnej gardła, a więc w miejscach, w których błona śluzowa styka się z kęsem p o k a r m o w y m w początkow y m okresie aktu połykania. Kubek s m a k o w y ( c a l y x gustatorius) jest t w o r e m kształtu jajowatego lub beczułkowatego, leżącym w błonie śluzowej. Długość jego wynosi około 70 (im, szerokość około 40 (im. Jest on zbudowany z podłużnych komórek nabłonkowych, przylegających do siebie jak klepki beczki. Koniec górny kubka ma otwór smakowy, który leży na poziomie powierzchni brodawki językowej. Niektóre komórki są zaopatrzone na końcu w pręcik smakowy, wystający na podobieństwo włoska przez otwór smakowy. Pręciki smakowe, stykające się z rozpuszczonym pokarmem, odbierają podniety smakowe. Od podstawy komórek nabłonka nerwowego odchodzą włókna nerwowe, biegnące do ośrodków smakowych mózgowia. Brodawki grzybowate występują głównie na końcu i bokach języka. Na ich powierzchni leżą po 3 - 4 kubki smakowe. Część tych brodawek (około 20%) nie ma zakończeń smakowych. Brodawki liściaste składają się z fałdów pionowych błony śluzowej, ułożonych jeden na drugim na bocznych stronach języka, bliżej jego nasady. W bruzdach brodawek liściastych leżą również brodawki smakowe. Brodawki okolone, w liczbie 8-12, układają się na powierzchni górnej języka w kształcie litery V, zwróconej ostrzem ku tyłowi. Środek brodawki okolonej jest podobny do brodawki grzybowatej,ztym że jest on otoczony w a ł e m błony śluzowej. Na bocznych ścianach brodawki oraz na powierzchni wewnętrznej wału są umieszczone kubki smakowe. Na dnie rowka, otaczającego środkową część brodawki, uchodzą gruczoły surowicze, których wydzielina wypłukuje resztki rozpuszczonych pokarmów, trafiających do rowka. Muszą one być stamtąd usunięte, aby dopuścić do zakończeń nerwowych następne próbki pokarmów*. Człowiek rozróżnia cztery podstawowe rodzaje smaku: gorzki, słodki, kwaśny i słony. Należy sądzić, że poszczególne kubki odczuwają tylko jeden rodzaj smaku, jak również, że liczne miejsca języka odbierają wrażenie jednego rodzaju. Smak słodki jest odbierany przez brodawki grzybowate przedniej części języka, kwaśny - przez części boczne i środek grzbietu języka, natomiast gorzki przez brodawki okolone. To ostatnie przypuszczenie potwierdzają spostrzeżenia, że przy porażeniu nerwu językowo-gardłowego roztwórchininy * Unerwienie brodawek okolonych i liściastych pochodzi z nerwu językowo-gardłowego. Brodawki grzybowate zaopatruje struna bębenkowa, gałąź nerwu twarzowego.
'468
może być przełknięty bez odczucia jego goryczy. Udało się również wyizolować włókna nerwowe wrażliwe jedynie na sól kuchenną i chininę. Jak w s p o m n i a n o wyżej, odczuwanie smaku potraw w dużej mierze zależy od powonienia. Człowiek pozbawiony węchu (np. przez znieczulenie okolicy węchowej kokainą) nie potrafi z zamkniętymi oczami odróżnić smaku cebuli od jabłka, oczywiście niezbyt kwaśnego ani słodkiego. W t y m stanie odróżnia on jedynie smaki podstawowe. Zmysł smaku jest znacznie mniej czuły od powonienia. Na przykład chinina jest wyczuwana w roztworze 1:2 000 000, kwas s o l n y - 1 : 1 3 0 000, sól kuchenna - 1:400, a cukier zaledwie w stężeniu - 1:200, podczas gdy zapach merkaptanu w y c z u w a m y w powietrzu w stężeniu 1:30 000 000 000.
NARZĄD POWONIENIA Nos, uważany w życiu codziennym za narząd powonienia, jest właściwie ochroną początkowego odcinka dróg oddechowych. Zabezpiecza on drożność nozdrzy, będących stale otwartą drogą powietrza zarówno do j a m y nosa, w której ulega ono klimatyzacji, jak i do dalszych odcinków drogi oddechowej. Jedynie w g ó r n y m odcinku jamy nosa znajduje się okolica w ę c h o w a błony śluzowej (regio olfactoria), posiadająca odmienną od okolicy oddechowej budowę i unerwienie, dzięki czemu człowiek może odbierać wrażenia węchowe. Okolica węchowa zajmuje błonę śluzową, pokrywającą z każdej strony powierzchnię przyśrodkową małżowiny nosowej górnej oraz odpowiadającą jej powierzchnię przegrody nosa. Ponieważ na każdej z tych kości powierzchnia węchowa zajmuje około 1 cm 2 , cała okolica węchowa mierzy około 4 cm 2 . U czworonogów na ogół powierzchnia ta jest znacznie większa, co wiąże się z niepomiernie lepiej rozwiniętym u nich węchem, potrzebnym przy poszukiw a n i u i zdobywaniu pokarmu. Stąd też zaliczamy ich do ssaków makrosmatycznych, podczas gdy człowiek należy do mikrosmatycznych. Zanik węchu u człowieka nastąpił prawdopodobnie w związku z jego postawą pionową, przy której polował on kierując się bardziej wzrokiem niż węchem, którym kierują się zwierzęta drapieżne tropiące ślady lub trawożerne, wietrzące grożącego drapieżnika. Komórki odbierające podniety węchowe, zwane k o m ó r k a m i w ę c h o w y m i , leżą w błonie śluzowej okolicy węchowej i oddają dwie wypustki. Jedna biegnie w kierunku powierzchni błony śluzowej i dochodząc do niej oddaje ze swego końca kilkaset drobnych włosków węchowych o długości 1 [im. Druga wypustka komórki węchowej biegnie w głąb błony śluzowej, stanowiąc neuryt tej komórki. W błonie śluzowej neuryty komórek w ę c h o w y c h tworzą splotowatą sieć, od której odchodzi około dwudziestu n e r w ó w w ę c h o w y c h (nerw olfactorii). Nerwy te dzielą się na dwie grupy, leżące po obu stronach przegrody nosa. Biegnąc ku górze dochodzą one do blaszki sitowej kości tej samej nazwy i przez '469
otworki wchodzą do jamy czaszki, gdzie tuż nad t y m i otworkami, po każdej stronie grzebienia koguciego ( crista galli) leży opuszka w ę c h o w a (bulbus olfactorius), należąca już do mózgowia. W opuszce węchowej nerwy tworzą kuliste ciałka o średnicy 0:1 m m , zwane kłębuszkami węchowymi. Powstają one w ten sposób, że nerwy w ę c h o w e rozgałęziają się drzewkowato i tworzą synapsy z p o d o b n y m i rozgałęziami wypustek tzw. komórek mitralnych, które stanowią drugi neuron drogi węchowej. Rozbudowane synapsy pierwszego i drugiego neuronu w ę c h o w e g o tworzą właśnie wspomniane kłębuszki węchowe. Łączące się ze sobą neuryty komórek mitralnych tworzą na powierzchni dolnej każdej półkuli mózgu pasma w ę c h o w e ( tractus olfactorii), biegnące do trójkąta węchowego na podstawie mózgu. Między komórkami w ę c h o w y m i leżą nieco niższe komórki podstawne. Poza t y m w błonie śluzowej znajdują się gruczoły węchowe, wydzielające śluz, który rozpuszcza substancje węchowe, zwilża i oczyszcza powierzchnię okolicy węchowej, jak również przyczynia się do nawilżania powietrza wdychanego. Zapachy są powodowane przez cząsteczki związków chemicznych unoszące się w powietrzu. Działają one trafiając na błonę śluzową okolicy węchowej. Ogromna różnorodność zapachów utrudnia ich klasyfikację. Zapachy eteryczne, np. o w o c ó w , w o n n e lub balsamiczne, np. kwiatów,ambrozja, np.piżma, sprawiają przyjemne wrażenie. Zapachy czosnkowe, spalenizny, kozłkowe, są znoszone rozmaicie, natomiast wyraźnie przykre są zapachy odrażające (kału, gnijącego mięsa czy produktów roślinnych). Zmysł węchu należy do najczulszych. Do badań węchu służą przyrządy zwane olfaktometrami. Czułość węchu u różnych ludzi jest niejednakowa. Przebywając w atmosferze jakiegoś zapachu przyzwyczajamy się do niego i po p e w n y m czasie przestajemy go odczuwać. Jeżeli zmieszamy dwie substancje wonne, z których jedna ma zapach silniejszy, to składnik o słabszym zapachu przestaje być odczuwany. Na t y m opiera się używanie perfum, trociczek czy kadzideł. Dzięki zmysłowi powonienia powstaje szereg odruchów warunkowych w układzie n e r w o w y m autonomicznym, wywierających w p ł y w na wydzielanie soków trawiennych i na cały proces trawienia. Powonienie ulega zaostrzeniu przy głodzeniu.
NARZĄD PRZEDSIONKOWO-ŚLIMAKOWY Narząd ten dawniej nazywano narządem słuchu i równowagi. Określenie to jest słuszne jedynie w swej pierwszej części, natomiast pojęcie narządu równowagi należałoby zastąpić określeniem czucia położenia i ruchów głowy, co nie byłoby wygodne. Ponieważ narząd słuchu mieści się w ślimaku, a narząd czucia położenia i ruchów głowy jest położony w przedsionku, oba narządy objęto wspólną nazwą z ich umiejscowieniem. '470
Ryc. 214. Narząd przedsionkowo-ślimakowy (półschematycznie).
Narząd ślimakowy Narząd słuchu, jak w i a d o m o , odbiera ze świata zewnętrznego wrażenia akustyczne, jak.tony, dźwięki, szmery i huki. Są one odbierane przez zakończenia nerwowe, umiejscowione w głębi czaszki, w uchu wewnętrznym, czyli błędniku kości skroniowej. Tony są w y w o ł y w a n e przez drgania różnych ciał, jak: struna, pręt, płyta itp., wywołujące powstawanie w powietrzu fal głosow y c h o określonej częstotliwości. Fale głosowe nie docierają bezpośrednio do zakończeń nerwowych, a są przewodzone najpierw przez ucho zewnętrzne i przenoszone na błonę bębenkową, z której przechodzą przez łańcuch kosteczek słuchowych, zawieszonych w uchu ś r o d k o w y m , na środowisko płynne w uchu wewnętrznym, które działa bezpośrednio na zakończenia nerwów. Od zakończeń n e r w ó w tych bodźce słuchowe biegną do mózgu i drogą słuchową do ośrodków słuchu, leżących w korze mózgowej płata skroniowego.
Ucho zewnętrzne Ucho zewnętrzne składa się z małżowiny usznej i przewodu słuchowego zewnętrznego. Małżowina ( auricula) jest wydłużoną, powyginaną płytką, otaczającą wejście do kanału słuchowego zewnętrznego ( meatus acusticus externus). Zadaniem jej jest kierowanie fal głosowych do przewodu. Tę rolę spełnia ona szczególnie u zwierząt, posiadających zdolność zwracania swych znacznie większych niż u człowieka uszu w kierunku źródła dźwięku. U człowieka małżowina uszna jest mało ruchoma z p o w o d u uwstecznienia mięśni. Małżowina uszna jest bardzo elastyczną miękką płytką, posiadającą szkielet zbudowany z chrząstki sprężystej. Złamania małżowiny zdarzają się wyjątko'471
wo i to pod w p ł y w e m silnych urazów. Kształty małżowiny są zmienne, jednak szereg jej cech jest dziedzicznie uwarunkowany. U bliźniąt jednojajowych małżowiny uszne są identyczne. Różnice budowy małżowiny łatwo zaobserwować, porównując jej kształty u paru osobników jednocześnie. Dolna część małżowiny; jej płatek, nie ma chrząstki. Wielkość jego jest bardzo różna, bywa on w o l n y albo przyrośnięty do skóry policzka. Małżowiny są zabarwione na różowo dzięki bogatemu ukrwieniu. Przy zaburzeniach krążenia i niedotlenieniu krwi występują na nich objawy sinicy, podobnie jak na wargach i paznokciach. Przewód słuchowy zewnętrzny w bocznej trzeciej części jest zaopatrzony w szkielet chrząstkowy, w dalszym zaś przebiegu wchodzi do przewodu kostnego słuchowego w kości skroniowej. Koniec wewnętrzny przewodu zamyka błona bębenkowa. Przewód słuchowy zewnętrzny jest pokryty skórą, zaopatrzoną zwłaszcza na początku, w grube, krótkie włosy, które stanowią do pewnego stopnia przeszkodę dla przenikania ciał obcych. W skórze znajdują się gruczoły łojowe, których wydzielina, mieszając się ze złuszczonym nabłonkiem, tworzy tzw. w o s k o w i n ę (cerumen). Rozrzedza ją wydzielina gruczołów apokrynowych, zwanych gruczołami w o s k o w i n o w y m i . Zatrzymując się w przewodzie słuchow y m , może ona zamknąć jego światło i spowodować znaczne osłabienie słuchu, a czasem również inne dolegliwości, jak szum w uszach, bóle itp. Chrząstka przewodu słuchowego tworzy w górnej ścianie szczelinę zamkniętą tkanką łączną. Dzięki temu głowa żuchwy, leżąca do przodu od przewodu, zmienia przy ruchach stawu skroniowo-żuchwowego światło przew o d u , co można wyczuć przy wprowadzeniu do niego opuszki małego palca (sprawdź!). Z tego też p o w o d u przy zapaleniu ślinianki przyusznej, która sięga do dołu zażuchwowego, chorzy odczuwają przy żuciu silne bóle ucha, które utrudniają przyjmowanie pokarmów, zwłaszcza stałych. W przewodzie słuchowym brak obfitszej tkanki podskórnej, dlatego też wszelkie toczące się w nim stany zapalne, zwykle połączone z obrzękiem, powodują silne bóle. Gałązki czuciowe przewodu słuchowego pochodzą od nerwu uszno-skroniowego oraz nerwu błędnego, dzięki któremu podrażnienia mechaniczne przewodu mogą na drodze odruchowej spowodować kaszel.
Błona bębenkowa Błona bębenkowa (membrana tympani) ma kształt owalu o wymiarach średnio 8,5 mm x 10 m m ; grubość jej wynosi około 0,2 m m . Jest ona zbudowana z włókien tkanki łącznej, przebiegającej okrężnie i promienisto, dzięki czemu ma dużą odporność przy zachowaniu dużej elastyczności. Powierzchnia błony jest pokryta od strony zewnętrznej nabłonkiem wielowars t w o w y m płaskim, zaś od strony wewnętrznej, skierowanej do jamy bębenkowej - błoną śluzową, wyściełającą tę jamę. '472
Od strony wewnętrznej do błony bębenkowej przylega młoteczek, którego koniec rękojeści zrasta się z błoną. Powoduje to pewne wciągnięcie błony do jamy bębenkowej. Błona bębenkowa jest zaopatrzona w naczynia krwionośne, dochodzące do niej od naczyń przewodu słuchowego zewnętrznego i j a m y bębenkowej. Obfite unerwienie czuciowe błony pochodzi od nerwu uszno-skroniowego i nerwu błędnego. Procesy ropne, toczące się w uchu środkowym, są bardzo bolesne i mogą doprowadzić do przebicia błony bębenkowej i wyjścia ropy do przewodu słuchowego. Dla zapobieżenia zniszczeniom błony w pewnych przypadkach wykonuje się przecięcie jej i otwarcie drogi dla odpływu ropy (paracenteza). Po zlikwidowaniu sprawy ropnej w uchu środkowym następuje wygojenie się rany i może wrócić zdolność słuchu.
Ucho środkowe Ucho środkowe składa się z jamy bębenkowej, kosteczek słuchowych i trąbki słuchowej. Jama bębenkowa Jama bębenkowa ( cavum tympani) jest ograniczona od strony zewnętrznej błoną bębenkową, zaś od strony wewnętrznej - ścianą kostną ucha wewnętrznego. Jest to wąska, szczelinowata przestrzeń, która w najciaśniejszym miejscu mierzy około 2 m m . Jama bębenkowa zawiera powietrze i jest połączona z przestrzeniami pneumatycznymi wyrostka sutkowatego kości skroniowej, jamą sutkową i komórkami sutkowymi. Stany zapalne ucha środkowego mogą przechodzić na te przestrzenie i spowodować konieczność umożliwienia o d p ł y w u ropy przez wydłutowanie ściany bocznej wyrostka sutkowego (trepanacja).
Kosteczki słuchowe W jamie bębenkowej są zawieszone na mięśniach i więzadłach trzy kosteczki słuchowe. Są to młoteczek (malleus), kowadełko ( incus) i strzemiączko (stapes). Jak w s p o m n i a n o , są one składnikiem biernym narządu słuchu, przenoszącym drgania błony bębenkowej na ucho wewnętrzne. Są to drobne i delikatne t w o r y - długość młoteczka wynosi 8 - 9 m m , tyle samo mniej więcej - długość kowadełka, strzemiączko jest o połowę mniejsze i najdelikatniejsze. Szczegóły budowy są podane na ryc. 215. Kosteczki słuchowe są połączone w ten sposób, że tworzą dźwignię dwuramienną, która przenosi drgania błony bębenkowej na podstawęstrzemiączka, wzmacniając je dwudziestokrotnie. Podstawa strzemiączka łączy się z błoną strzemiączkową, która zamyka okienko przedsionka; jest to otwór w ścianie '473
Kowadełko
/
Głowa młoteczkaMłoteczek-
•
Rękojeśi młotecz
I
/
Podstawa strzemiączka
Strzemiączko Ryc. 215. Układ kosteczek słuchowych.
przyśrodkowej jamy bębenkowej, który łączy ją z uchem wewnętrznym. Przy drganiach błony bębenkowej podstawa strzemiączka wielokrotnie szybko wchodzi do okienka przedsionka i cofa się z niego dzięki elastyczności błony strzemiączkowej. Ruchy te można porównać do ruchów tłoka, z t y m że są one bardzo płytkie lecz bardzo częste, gdyż odpowiadają falom akustycznym.
Trąbka słuchowa Jak już w i e m y , jama bębenkowa jest wypełniona powietrzem. Wiadomo jednak również, że powietrze, które dostaje się do jam ciała, zostaje - jak m ó w i m y - wessane, a właściwie rozpuszczone we krwi. Z doświadczenia własnego w i e m y też, że przy nagłych zmianach ciśnienia powietrza, jak np. przy starcie samolotu, zaczynamy gorzej słyszeć. Dzieje się tak dlatego, że dla prawidłowej czynności błony bębenkowej musi panować w przewodzie słuchowym i w jamie bębenkowej jednakowe ciśnienie, gdyż w przeciwnym razie elastyczna błona bębenkowa zostanie wciągnięta do wewnątrz w przypadku mniejszego ciśnienia w jamie bębenkowej lub odwrotnie - uwypuklona do przewodu przy niższym ciśnieniu powietrza na zewnątrz (jak przy starcie). Zapobiega temu trąbka słuchowa ( tuba auditiva), stanowiąca połączenie jamy bębenkowej z jamą gardła, w której ciśnienie powietrza jest mniej więcej równe panującemu ciśnieniu atmosferycznemu, dzięki otwartemu połączeniu j a m y gardła z jamą nosową. Ponieważ jednak ujście gardłowe trąbki słuchow e j może być z różnych p o w o d ó w zamknięte, wykonujemy parę ruchów połykania i prawidłowy słuch zostaje przywrócony, gdyż mięśnie podniebienia miękkiego, czynrie przy połykaniu, jednocześnie rozszerzają ujście trąbki '474
słuchowej. Taki jest p o w ó d podawania pasażerom przez obsługę samolotu kwaśnych cukierków do ssania, które wzmagają wydzielanie śliny i zmuszają do jej częstszego połykania. Również z doświadczenia w i e m y , że przy silnym katarze gorzej słyszymy. Dzieje się to z p o w o d u zajęcia przez stan zapalny również śluzówki części nosowej gardła i obrzęku ujścia trąbki słuchowej, co prowadzi do niedostatecznej wentylacji j a m y bębenkowej.
Ucho wewnętrzne Ucho wewnętrzne składa się z szeregu połączonych ze sobą przestrzeni w postaci jam i kanałów, wydrążonych w części skalistej (piramidzie) kości skroniowej, tworzących błędnik (labirynt) kostny (labyrinthusosseus). Składa się nań większa przestrzeń, zwana przedsionkiem (vestibulum), kanały półkoliste ( canales semicirculares) i ślimak (cochlea). Błędnik kostny jest wypełniony płynem, zwanym przychłonką ( p e r i l y m p h a ) , który licznymi drogami komunikuje się z płynem podpajęczynówkowym czaszki. Wewnątrz błędnika kostnego mieści się błędnik błoniasty ( l a b y r i n t h u s membranaceus), zawieszony na licznych pasemkach łącznotkankowych, umocowanych na ścianach kostnych i otoczony przychłonką. Jest to zamknięty worek łącznotkankowy, nieco mniejszy od błędnika kostnego, posiadający uchyłki, wchodzące do poszczególnych części błędnika kostnego. W przedsionku leżą woreczek ( sacculus) i łagiewka (utriculus), w innych częściach odpowiednio kanały półkoliste błoniaste i ślimak błoniasty. Błędnik błoniasty jest wypełniony śródchłonką (endolympha), która jest znacznie gęstsza od przychłonki i nigdzie się z nią nie komunikuje, natomiast od przestrzeni endolimfatycznych przedsionka prowadzi przewód śródchłonki do jej zbiornika, leżącego na tylnej powierzchni części skalistej kości skroniowej w przestrzeni podpajęczynówkowej. Nosi on nazwę w o r k a śródchłonki ( saccus endolymphaticus). Opiszemy kolejno części ucha wewnętrznego, rozpoczynając od jego najbardziej skomplikowanej części - ślimaka.
Ślimak Ślimak kostny tworzy dwa i pół skrętu i leży do przodu od przedsionka. Wewnątrz ślimaka wzdłuż jego osi długiej mieści się'.kostne wrzecionko (modiolus), mające postać krótkiego stożka. Dokoła wrzecionka owija się wyrastająca z niego kostna blaszka spiralna (lamina spiralis ossea), kierująca się ku górze. Blaszka ta nie dochodzi do przeciwległej ściany kanału ślimaka, ale kończy się ostrym brzegiem, od którego rozpoczyna się błona spiralna (membrana spiralis), łącząca się z boczną ścianą ślimaka kostnego. Błona ta jest częścią ślimaka błoniastego. '475
Blaszka spiralna kostna i błona spiralna tworzą razem blaszkę podstawną (lamina basilaris), która dzieli poprzecznie kanał spiralny ślimaka (canalis spiralis cochleae) na dwa piętra, zwane schodami. Niższe piętro, bliższe podstawy ślimaka, nosi nazwę schodów bębenka (scala tympani), piętro wyższe stanowią schody przedsionka (scala vestibuli). Schody te są ze sobą połączone szparą osklepka, leżącą przy wierzchołku ślimaka. W świetle schodów przedsionka przebiega błona przedsionkowa (membrana vestibularis), oddzielająca dolną zewnętrzną część ich przestrzeni, która tworzy przewód ślimakowy (ductus cochlearis). W przewodzie ślimaka na błonie podstawnej mieści się narząd ś l i m a k o w y (Cortiego), mający zakończenia nerwowe słuchowe. Jak już w i e m y , przewód ślimakowy jest wypełniony śródchłonką. ^ Narząd ś l i m a k o w y jest skomplikowanym t w o r e m , położonym na blaszce podstawnej przewodu ślimakowego. Zasadniczą jego częścią są komórki nerwowe zmysłowe, zwane komórkami rzęsatymi albo słuchowymi. Są one ułożone w komórkach zrębowych podpórkowych, których rozróżniamy trzy rodzaje: komórki filarowe, komórki Deitersa* i komórki Hensena**. Komórki filarowe, odsunięte od siebie podstawami, tworzą tzw. tunel wewnętrzny Cortiego, po obu stronach którego leżą komórki rzęsate (słuchowe) wewnętrzne i zewnętrzne. Pierwsze tworzą jeden rząd, drugie od trzech do pięciu rzędów. Do każdej komórki rzęsatej dochodzi włókienko nerwowe. Komórki rzęsate otrzymały swą nazwę od wystających z ich wolnego końca rzęsek, czyli pręcików. Liczba tych rzęsek w komórkach wewnętrznych wynosi 30-60, natomiast w zewnętrznych do 100. Od blaszki spiralnej odchodzi w kierunku filarów błona pokrywająca (membrana tectoria), której w o l n y brzeg układa się nad komórkami rzęsatymi. Prześledźmy teraz drogę fal głosowych, poczynając od okienka przedsionka. Drgania strzemiączka powodują powstawanie fal w przychłonce, otaczającej kanał ślimakowy, a więc na schodach przedsionka i na połączonych z nimi szparą osklepka schodach ślimaka. Ponieważ przychłonka, jak każdy płyn, jest praktycznie nieściśliwa, dla umożliwienia jej drgań powinno się znaleźć w ścianach kostnych przedsionka jeszcze jakieś miejsce elastyczne, wrażliwe na działanie strzemiączka, wciskanego na podobieństwo tłoka do okienka przedsionka. Takim miejscem jest okienko ślimaka, leżące w ścianie przyśrodkowej ucha środkowego, położone nieco niżej od okienka przedsionka. Jest ono zamknięte tzw. błoną bębenkową w t ó r n ą , po wewnętrznej stronie której mieszczą się schody bębenka. Kiedy strzemiączko wchodzi do okienka przedsionka, uwypukla się w stronę jamy bębenkowej błona bębenkowa wtórna, zamykająca okienko ślimaka. Odwrotnie, cofanie się strzemiączka do jamy bębenkowej powoduje wpuklenie się błony bębenkowej wtórnej do schodów ślimaka. Fale akustyczne, przebiegające w przychłonce schodów przedsionka i bębenka, powodują powstawanie drgań błony podstawnej kanału spiralnego, * Komórki falangowe zewnętrzne. ** Komórki graniczne zewnętrzne.
'476
przez który komórki rzęsate stykają się z błoną pokrywającą i zostają przez to mechanicznie podrażnione, wskutek czego w komórkach tych powstają impulsy elektryczne. Impulsy te biegną drogą włókien nerwowych do ośrodków słuchowych w korze mózgowej płata skroniowego, gdzie zostają odczute, jako dźwięki. Wspomniane włókna nerwowe komórek rzęsatych pochodzą z części ślimakowej nerwu VIII, czyli przedsionkowo-ślimakowego. Badania ruchów kosteczek słuchowych na zwierzętach i zwłokach ludzkich wykazały, że ruchy te muszą pokonywać pewne opory, wzrastające w miarę zwiększania ich częstotliwości. Opór kosteczek słuchowych przy 2000 drgań na sekundę staje się tak duży, że praktycznie uniemożliwia normalne przenoszenie drgań drogą tych kosteczek. Człowiek słyszy dźwięki o częstotliwości od 16 do 20 000 drgań na sekundę. Ucho ludzkie jest najbardziej wrażliwe na dźwięki o drganiach między 500 a 5000/s. M o w a ludzka obejmuje zakres od 100 do 800 drgań, instrumenty muzyczne od 30 do 4000, a więc dźwięki, które najczęściej słyszymy, mieszczą się w zakresie drgań, przewodzonych przez kosteczki słuchowe. Należy jednak sądzić, że są jeszcze inne drogi przewodzenia dźwięków. Otóż znanym zjawiskiem jest przewodnictwo kostne. Jeżeli zatkamy przewody słuchowe i dotkniemy widełkami stroikowymi głowy, to usłyszymy dźwięk docierający do ucha wewnętrznego drogą kostną. Sądzi się więc na ogół, że dźwięki o niższej częstotliwości są odbierane drogą kosteczek słuchowych, natomiast o wyższej - przez przewodnictwo kostne. Niektórzy autorzy przyjmują możliwość przewodnictwa drgań błony bębenkowej przez jamę bębenkową drogą powietrzną na błonę bębenkową wtórną i następnie do przewodu ślimakowego.
Kanały półkoliste Jak już w i e m y , w błędniku kostnym przebiegają trzy kanały półkoliste (canales semicirculares), które leżą mniej więcej w trzech płaszczyznach zasadniczych. Kanał przedni układa się nieco skośnie w stosunku do płaszczyzny strzałkowej, tylny - dó czołowej, boczny leży w płaszczyźnie poziomej. W kanałach kostnych mieszczą się kanały półkoliste błoniaste, które kanałów kostnych nie wypełniają. Mają one 0,5 mm średnicy i zajmują zaledwie 1 / 5 powierzchni przekroju kanału kostnego. Przewody błoniaste są otoczone przychłonką i powiązane ze ścianami kanałów kostnych cienkimi pasemkami tkanki łącznej. Każdy kanał półkolisty kończy się bańką, w której leży sierpowaty t w ó r , zwany grzebieniem bańkowym, ustawiony poprzecznie do płaszczyzny odpowiedniego przewodu półkolistego. Grzebienie są pokryte nabłonkiem nerwow y m z m y s ł o w y m ( n e u r o e p i t h e l i u m ) , wykazującym zdolność odbierania podniet. Komórki tego nabłonka mają na powierzchni włoski i są jak gdyby oplecione drobnymi rozgałęzieniami części przedsionkowej nerwu VIII (przedsionkowo-ślimakowego). '477
Przewód półkolisty przedni (pionowy)
Bańka błoniasta przednia IM. twarzowy Woreczek j
dl
A
/ -
Przewód półkolisty boczny (poziomy) Przewód półkolisty --„ / t y l n y (pionowy)
m
Ślimak Część ślimakowa f n. VIII Część przedsionkowa n. VIII Ryc. 216. Błędnik błoniasty prawego ucha.
Powierzchnia wolna grzebienia ma kształt siodełkowaty. Leży na niej galaretowaty t w ó r , zwany osklepkiem, do którego wnikają włoski odchodzące od komórek nabłonka nerwowego, pokrywających grzebień bańkowy. Jak już w s p o m n i a n o wyżej, kanały półkoliste są wypełnione śródchłonką, która zmienia położenie przy ruchach głowy dzięki swej bezwładności. Na przykład przychłonką kanałów półkolistych bocznych, ułożonych na płaszczyźnie poziomej, przy obrocie g ł o w y w lewo przemieści się w stosunku do ścian kanałów półkolistych w prawo. Prąd śródchłonki porusza osklepek grzebieni bańkowych i podrażnia włoski nabłonka nerwowego w obu uszach, skąd stany pobudzenia zostają przeniesione drogą gałązek nerwu VIII do ośrodkowego układu nerwowego i odczute, jako obrót g ł o w y w lewo. Podobnie bywa przy innych ruchach głowy, jak np. przy ruchu potakiwania, tylko że w t y m przypadku ruch śródchłonki odbywa się w kanałach półkolistych przednich, a przy kiwaniu głowy na boki - w kanałach tylnych. Z ruchami głowy są połączone niektóre odruchy gałek ocznych, jak np. oczopląs (szybkie m i m o w o l n e ruchy gałek na boki).
Łagiewka i woreczek W przedsionku kostnym ucha wewnętrznego znajdują się jeszcze dwie części błędnika błoniastego, a mianowicie wydłużona łagiewka (utriculus) i bardziej kulisty woreczek (sacculus). Podobnie jak inne części błędnika błoniastego są one otoczone śródchłonką, która tworzy tu stosunkowo duży '478
zbiornik, połączony z przestrzenią podpajęczynówkową mózgowia przewodem przychłonkowym, wychodzącym z ucha wewnętrznego na powierzchni dolnej części skalistej kości skroniowej. W łagiewce i woreczku znajdują się powierzchnie pokryte nabłonkiem n e r w o w y m , które noszą nazwę plamek. Plamka łagiewki jest płaszczyzną poziomą o wymiarach około 3 x 2 m m ; plamka woreczka jest nieco mniejsza i umieszczona w płaszczyźnie strzałkowej. Nabłonek n e r w o w y jest podobny do nabłonka grzebieni bańkowych. Zamiast osklepka jest on pokryty galaretowatą błoną kamyczkową, do której wchodzą włoski komórek nabłonka nerwowego. Błona otrzymała swą nazwę z powodu licznych skupień kryształków węglan ó w i fosforanów wapnia, zwanych kamyczkami błędnikowymi. Kamyczki te mają wysoki ciężar właściwy i uciskając lub pociągając włoski komórek nabłonka nerwowego wywołują w nim stany pobudzenia, które drogą gałęzi nerwu VIII są przenoszone do ośrodkowego układu nerwowego, gdzie są przetwarzane na wrażenia o położeniu ciała w przestrzeni. W postawie stojącej kamyczki błędnikowe uciskają na błonę kamyczkową łagiewki, natomiast w pozycji leżącej na grzbiecie uciskten ustaje, a powstaje pociąganie w kierunku brzegu tylnego plamki. W woreczku przy zmianie postawy stojącej na pozycję leżącą na boku występują stosunki odwrotne.
NARZĄD WZROKU Wśród narządów zmysłów najbardziej złożony jest narząd w z r o k u (organum visus). Jest on przystosowany do odbierania wrażeń wzrokowych w najrozmaitszych warunkach. Widzimy zarówno w południe, jak i o zmierzchu, a nawet w nocy, dzięki urządzeniu regulującemu ilość światła, wpadającego do wnętrza oka. Rozróżniamy szereg barw o bogatej skali odcieni, co prawda jedynie przy należytym oświetleniu: o zmierzchu tracimy tę zdolność. Dzięki specjalnej budowie oka widzimy wyraźnie przedmioty bliskie i dalekie. Mamy zdolność widzenia przestrzennego (stereoskopowego), jak również możemy ocenić odległość oglądanych przedmiotów. Dalej, dzięki wybitnej ruchliwości gałki ocznej, możemy oglądać szybko poruszające się przedmioty, duże obrazy, nie mieszczące się w jednym polu widzenia itp. Do tych rozmaitych czynności narząd wzroku ma specjalne urządzenia.
Budowa ogólna Najważniejszą częścią narządu wzroku jest gałka oczna ( bulbus oculi) z n e r w e m w z r o k o w y m (nervus opticuś). Leży ona w czaszce ukryta w kostnej puszce zwanej oczodołem (orbita), otwartym od przodu, który komunikuje się z leżącą do tyłu od niego jamą czaszki o t w o r e m dla nerwu wzrokowego i szczelinami oczodołowymi. Ze względu na dużą wrażliwość gałki ocznej '479
ochrania ją przed urazami ciało tłuszczowe oczodołu, stanowiące miękkie łożysko, które nie hamuje jej ruchów. W oczodole mieszczą się również mięśnie gałki ocznej i narząd łzowy, należące do narządów dodatkowych oka. Od przodu gałka oczna jest zasłonięta powiekami, które jednocześnie zmywają gałkę oczną, rozprowadzając po niej ciecz łzową.
Gałka oczna ma kształt zbliżony do spłaszczonej nieco kuli. Jej średnica poprzeczna wynosi około 24 m m , zaś pionowa o jeden milimetr mniej. W y m i a r przednio-tylny jest nieco większy od poprzecznego, gdyż gałka w swej przedniej części jest bardziej wypukła. Wypukłość ta stanowi 1 / 6 część kuli o promieniu nieco mniejszym niż reszta gałki ocznej. To uwypuklenie łatwo stwierdzić dotykiem, jak również oglądając zamknięte powieki, zwłaszcza u małych dzieci podczas snu. Najbardziej wysunięte do przodu i do tyłu punkty gałki ocznej nazywamy biegunami - przednim i t y l n y m (polus anterior et posterior). Linia łącząca te bieguny nosi nazwę osi optycznej gałki ( axis opticus). Linię największego o b w o d u gałki nazywamy jej r ó w n i k i e m ( equator ). Od powierzchni tylnej gałki ocznej, nieco w stronę przyśrodkową od osi optycznej, odchodzi nerw wzrokow y , który jest g r u b y m pęczkiem włókien nerwowych, biegnących do podstaw y mózgu. Przy ucisku na gałkę oczną odczuwamy jej podatność, uwarunkowaną t y m . '480
że jej wnętrze jest wypełnione galaretowatą masą, zwaną ciałem szklistym (corpus vitreum) i cieczą wodnistą ( humor aąuosus). We wnętrzu gałki leży również aparat załamujący promienie świetlne.
Gałka oczna Ściana gałki ocznej składa się z trzech błon, ściśle przylegających do siebie w części tylnej oka, natomiast częściowo rozdzielonych w odcinku przednim gałki. Warstwa zewnętrzna, najmocniejsza, zbudowana z tkanki łącznej, nosi nazwę błony włóknistej. Warstwa pośrednia - to błona naczyniowa, ogólnie biorąc odżywiająca gałkę. Wreszcie najgłębsza, siatkówka, czyli błona wewnętrzna oka, to warstwa przeważnie światłoczuła.
Błona włóknista gałki ocznej Błona w ł ó k n i s t a (tunica fibrosa oculi), mocna i odporna, dzieli się na dwie części. Tylna, biaława, stanowiąca 4 / 5 powierzchni całej błony włóknistej, nosi nazwę t w a r d ó w k i . Część przednią, przeźroczystą, zajmującą V 5 wspomnianej powierzchni, nazywamy rogówką.
Twardówka Jak już w s p o m n i a n o , twardówka stanowi warstwę ochronną gałki ocznej. Grubość jej nie wszędzie jest jednakowa. W części tylnej, w otoczeniu nerwu wzrokowego, osiąga ona 1 - 2 m m , na równiku wynosi 0,4-0,5 m m , w części przedniej około 0,3 m m . W miejscach przyczepu mięśni gałki ocznej grubość twardówki dochodzi do 0,6 m m . Twardówka jest zbudowana z włókien tkanki łącznej, przebiegających falisto, co ma duże znaczenie dla gałki ocznej. Chodzi o to, że oko może działać prawidłowo jedynie przy określonym ciśnieniu wewnątrz gałki ocznej. Otóż w przypadkach wzrostu czy spadku ciśnienia śródocznego twardówka może się nieco rozciągać lub kurczyć, przyczyniając się do wyrównania tego ciśnienia. Od zewnątrz twardówka jest pokryta warstwą tkanki łącznej wiotkiej, zwanej blaszką nadtwardówkową, natomiast na powierzchni przylegającej do błony naczyniowej występują liczne komórki barwnikowe, tworzące blaszkę brunatną twardówki. W tylnej części twardówki znajduje się powierzchnia pokryta otworkami, zwana polem s i t o w y m . Przez tę blaszkę przechodzą pęczki włókien nerwowych, które tworzą wspomniany już nerw wzrokowy, biegnący do podstawy mózgu. Poza t y m w innych ścianach twardówki występuje szereg otworków, przez które przechodzą naczynia i nerwy, o czym będzie m o w a dalej. 31 - Anatomia i fizjologia
481
Barwa twardówki zmienia się z wiekiem. U dzieci jest ona niebieskawa, u dorosłych szarawa, w wieku starczym żółtawa - z p o w o d u odkładania się w niej złogów tłuszczowych. W przedniej części gałki ocznej, wzdłuż granicy między twardówką i rogówką, przebiega kanał, zwany zatoką żylną twardówki. Kanał ten zawiera ciecz wodnistą oka, która tą drogą odpływa do żył rzęskowych i spojówkowych. W b r e w nazwie w zatoce nie ma krwi, natomiast jej zawartość uchodzi do krwi. Rogówka Rogówka (cornea) jest całkowicie przeźroczystą, gładką, lśniącą częścią błony włóknistej, łączącą się z twardówką. W miejscu tego połączenia przebiega kolista bruzda, zwana bruzdą twardówki. Powstaje ona z tego powodu, że rogówka jest bardziej wypukła niż twardówka. Rogówka jest jak gdyby wprawiona w twardówkę na podobieństwo szkła od zegarka, albo też łączy się z nią jak kości w szwie łuskowym. Rogówka od tyłu jest okrągła, natomiast oglądana od przodu ma kształt elipsy ze względu na to, że od dołu i od góry na jej powierzchnię przednią zachodzi twardówka. Odbija się to na wymiarach rogówki, która w linii poziomej mierzy około 11,6 m m , zaś w linii pionowej - 11,2 m m . Część środkowa rogówki, zwana jej szczytem, ma grubość około 8 m m , zaś na obwodzie grubość ta wynosi 1,1 m m . U dzieci cała rogówka jest zupełnie przejrzysta, natomiast u osób starszych w górnej części rogówki widoczny jest zmętniały łuk sierpowatego kształtu. Przebiega on równolegle do brzegu w odległości 1 - 2 mm i nosi nazwę łuku starczego (arcus senilis). Z biegiem czasu podobny łuk powstaje na dolnej krawędzi rogówki. Łącząc się ze sobą oba łuki zamykają pierścień zmętniały, zwany o b w ó d k ą starczą (gerontoxon). Wspomniana przejrzystość rogówki występuje w wyniku braku w niej naczyń krwionośnych, jak również osłonek dość licznych nerwów. Warunkiem jej przejrzystości, a więc i należytego spełniania swego zadania, jest pozostawienie jej w środowisku w o d n y m , zawierającym określone stężenie soli kuchennej (0,9 %). Zapewnia jej to ciecz łzowa, stale wydzielana przez gruczoły łzo we. Powierzchnia przednia rogówki jest pokryta tzw. nabłonkiem przednim, obficie unerwionym, dzięki czemu rogówka jest bardzo wrażliwa na urazy mechaniczne (obce ciała, jak kurz, piasek), podrażnienie chemiczne (gazy, dym) oraz wysychanie. Przy podrażnieniach rogówki następuje odruchowe zamknięcie powiek i łzawienie. Przy uszkodzeniu mechanicznym nabłonka przedniego regeneruje się on w ciągu kilku godzin, natomiast uszkodzenia głębsze mogą prowadzić do powstawania blizn, powodujących niedowidzenie. Główną warstwę rogówki stanowi jej istota właściwa, stanowiąca 9 /io jej grubości. Warstwie tej rogówka zawdzięcza swą odporność i zachowanie kształtu. '482
Błona naczyniowa gałki ocznej Jak w s p o m n i a n o wyżej, pod błoną włóknistą leży następna warstwa gałki ocznej - błona naczyniowa ( tunica vascularis bulbi). Dzieli się ona na trzy części: naczyniówkę, ciało rzęskowe i tęczówkę. Naczyniówka Naczyniówka (chorioidea) stanowi tylną część błony naczyniowej, pokrywającą od zewnątrz część wzrokową siatkówki. Składa się ona głównie z sieci naczyń włosowatych, które mają stosunkowo dużą średnicę i mogą pomieścić kilka leżących obok siebie krwinek czerwonych. Naczynia te tworzą gęstą sieć, której oczka są mniejsze od średnicy naczyń. Między naczyniami biegną włókna sprężyste, które powodują napięcie naczyniówki. Z naczyń gałki ocznej zbierają się zazwyczaj cztery duże żyły wirowate, które przebijają twardówkę do tyłu od równika i uchodzą do żyły wrotnej. Krew tętniczą dochodzi do gałki ocznej drogą tętnicy ocznej od tętnicy szyjnej wewnętrznej. Ciało rzęskowe Ciało rzęskowe ( c o r p u s ciliare) jest to twór w kształcie pierścienia, który powstaje z przedniego odcinka błony naczyniowej. Ciało rzęskowe leży między tęczówką i rąbkiem zębatym siatkówki, o czym będzie m o w a niżej. Pierścieniowate ciało rzęskowe otacza soczewkę, która jest umocowana do jego brzegów licznymi cienkimi nićmi łącznotkankowymi, tworzącymi o b w ó d k ę rzęskową soczewki ( zonula ciliaris). Powierzchnia zewnętrzna ciała rzęskowego przylega do twardówki, wewnętrzna jest skierowana do ciała szklistego, wypełniającego gałkę oczną. Na powierzchni tylnej ciała rzęskowego leży promienisto szereg drobnych fałdów (70-80) wysokości 0,6-0,8 m m , długości około 2 m m . Są to w y r o s t k i rzęskowe (processus ciliares), w których występują liczne naczynia krwionośne. W skład ciała rzęskowego wchodzą również włókna mięśniowe gładkie, które leżąc na jego obwodzie przebiegają okrężnie i południkowo. Tworzą one mięsień rzęskowy (musculus ciliaris), który odgrywa zasadniczą rolę w procesie akomodacji oka (p. dalej). Mięsień rzęskowy leży między twardówką a wyrostkami rzęskowymi. Jego włókna południkowe rozpoczynają się w okolicy równika w przestrzeni nadnaczyniówkowej i biegną w kierunku kąta tęczówkowo-rogówkowego. Włókna okrężne leżą najgłębiej, w płaszczyźnie obwódki rzęskowej. Przy skurczu włókien okrężnych zmniejsza się o b w ó d wyrostków rzęskowych i przez to aparat wieszadłowy się rozluźnia. Włókna południkowe pociągają naczyniówkę do przodu razem z ciałem szklistym, co również rozluźnia napięcie obwódki rzęskowej. Poza t y m działanie mięśnia rzęskowego, a więc jego skurcze i rozkurczę, powodują odpływ cieczy wodnistej z komory przedniej oka przez więzadło grzebieniaste (p. Komora przednia gałki ocznej.). '483
Tęczówka Tęczówka (/r/s) jest najbardziej wysuniętą do przodu częścią błony naczyniowej. Jest ona widoczna przez rogówkę i zawdzięcza swą nazwę wielkiej rozmaitości zabarwienia. Barwa tęczówki przechodzi rozmaite odcienie - od żółtej przez zieloną i niebieską do fiołkowej lub przez brązową do prawie czarnej. Barwa ta zależy od ilości barwnika i od b u d o w y powierzchni przedniej tęczówki. Powierzchnia tylna tęczówki jest zabarwiona na czarno. Tęczówka jest błoną okrągłego kształtu, ma średnicę ok. 12 mm i grubość ok. 0,5 m m , w środku ma otwór o zmiennej wielkości, zwany źrenicą (pupilla). Tęczówka zamyka od tyłu komorę przednią oka i oddziela ją od komory tylnej. Brzeg źreniczny (wewnętrzny) tęczówki otacza źrenicę i ślizga się po powierzchni przedniej soczewki, która od tyłu zamyka źrenicę. Brzeg rzęskowy (zewnętrzny) tęczówki jest połączony z ciałem rzęskowym. Tęczówka, jako bogato ukrwiona część błony naczyniowej, bierze udział w wytwarzaniu cieczy wodnistej, a poza t y m jest ważnym urządzeniem, regulującym ilość światła wchodzącego dó wnętrza gałki ocznej. Jeżeli porównamy gałkę oczną z aparatem fotograficznym, to tęczówka odpowiada przysłonie. Do naświetlenia kliszy, to znaczy do wykonania zdjęcia, m u s i m y użyć odpowiedniej p r z y s ł o n y - t y m szerszej, im słabsze jest oświetlenie przedmiotu fotografowanego. Podobnie też źrenica oka rozszerza się przy słabym oświetleniu. W jasny, słoneczny dzień źrenice są silnie zwężone, aby nie dopuścić nadmiaru światła do siatkówki oka. Zwężenie źrenic ma też inne znaczenie. W i e m y , że im mniejszy jest otwór przesłony przy zdjęciu, t y m ostrzejszy obraz powstaje na kliszy. Podobnie dzieje się z okiem człowieka, używającego szkieł. Bez okularów odczuwa on bardziej dotkliwie wadę swego wzroku o zmierzchu, kiedy źrenice jego są bardziej rozszerzone lub w pochmurny dzień. W ciemności, przy maksymalnie rozszerzonych źrenicach, średnica ich wynosi 7 - 8 m m , przy silnym oświetleniu 3 - 2 m m , a nawet mniej. Zmiana średnicy źrenic zachodzi dzięki działaniu mięśni antagonistycznych - zwieracza i rozszerzacza źrenicy (mm. sphincter et dilatator pupillae). Pierwszy tworzą włókna mięśniowe, przebiegające przy brzegu rzęskowym tęczówki, natomiast włókna ułożone promienisto dokoła źrenicy tworzą mięsień rozszerzacz. Mięśnie źrenicy są unerwione przez układ autonomiczny: zwieracz - przez włókna przywspółczulne, rozszerzacz - przez współczulne. Błona w e w n ę t r z n a gałki ocznej Błona w e w n ę t r z n a gałki ocznej (tunica interna bulbi), stanowiąca światłoczułą warstwę, wyścieła powierzchnię wewnętrzną gałki. Ponieważ załamane w soczewce promienie światła trafiają tylko na ścianę tylną gałki, warstwa ta, zwana również siatkówką (retina), zachowała wrażliwość na światło jedynie w części leżącej ku tyłowi od równika gałki - w przeciwieństwie do leżącej do '484
przodu części ślepej. Część tylna siatkówki nosi nazwę części w z r o k o w e j ( pars optica retinae). Kończy się ona rąbkiem zębatym (ora serrata) w odległości około centymetra do przodu od równika, gdzie przechodzi na ciało rzęskowe, jako część rzęskowa siatkówki, oraz na tęczówkę - jako część tęczówkowa. Część wzrokowa siatkówki jest cienką (0,2-0,4 m m ) przeźroczystą błoną barwy różowej, którą zawdzięcza ona barwnikowi rodopsynie, zawartej w komórkach. Na zwłokach siatkówka przybiera barwę szarą wskutek rozkładu rodopsyny i zmętnienia komórek światłoczułych. Między siatkówką a przylegającą do niej błoną naczyniową leży ciemna warstwa barwnikowa, pochłaniająca światło, dzięki czemu, jak w s p o m n i a n o wyżej, źrenice są czarne, gdyż widzimy w nich właśnie tę warstwę. Siatkówka jest związana z podłożem dość luźno, dzięki czemu może się na nim przesuwać. Jest ona przymocowana jedynie przy wyjściu nerwu wzrokow e g o z gałki ocznej i na rąbku zębatym, w miejscu przejścia w część rzęskową. Na ogół utrzymuje się ona przy błonie naczyniowej dzięki ciału szklistemu i cieczy wodnistej, które w gałce wytwarzają ciśnienie śródoczne. Przy jego spadku w pewnych stanach patologicznych może nastąpić tzw. odklejenie siatkówki ( ablatio retinae) z groźnymi dla zdolności widzenia objawami. Siatkówka ma bardzo skomplikowaną budowę. Składa się ona z dziesięciu warstw, w ś r ó d których znajdujemy warstwę komórek wrażliwych na światło i barwy. Istnieją dwa rodzaje tych komórek. Jedne z nich - wydłużone, są wrażliwe na stopień natężenia światła, dające obrazy czarno-białe oglądanych przedmiotów. Noszą one nazwę pręcików [bacilli); liczbę ich szacuje się na 125 min. Drugi rodzaj komórek siatkówki - to tzw. czopki (coni). Mają one kształt buteleczek, szyjkami skierowanych do warstwy barwnikowej. Czopki są wrażliwe na barwy, jednak mogą je odbierać tylko przy d o b r y m oświetleniu. O zmroku i w nocy barw już nie rozróżniamy. Możemy jedynie odróżnić kolory silnych źródeł światła, jak neony, światła sygnałowe itp. Ogólna liczba czopków jest szacowana na 7 min. Część wzrokowa siatkówki nie wszędzie jest jednakowo czuła. W części tylnej siatkówki, na osi wzrokowej, leży miejsce, zwane plamką ( macula), dawniej nazywane plamką żółtą z powodu odmiennego zabarwienia. Jest ono najwrażliwszym miejscem siatkówki dzięki szczególnemu zagęszczeniu komórek światłoczułych. Natomiast w miejscu wyjścia nerwu wzrokowego znajduje się część siatkówki całkowicie niewrażliwa na światło. Jest to małe białawe pole o średnicy ok. 1,5 m m , które leży w odległości około 3 mm od bieguna tylnego gałki w kierunku przyśrodkowym i nieco wyżej od jego poziomu. Nosi ono nazwę tarczy n e r w u w z r o k o w e g o ( discus nervi optici), ponieważ w obrębie tarczy nie ma komórek światłoczułych, siatkówka w t y m miejscu nie odbiera wrażeń wzrokowych. Łatwo to sprawdzić na doświadczeniu Mariotte'a (ryc. 218). Jak wspomniano, rozmieszczenie pręcików i czopków nie jest jednakowe. W dołku środkowym plamki znajdują się wyłącznie czopki, dokoła niego pojawiają się poszczególne pręciki, których liczba rośnie ku o b w o d o w i . Z tego '485
Ryc. 218. Doświadczenie Mariotta. Zasłoń lewe oko i skieruj wzrok na krzyż. Przesuwaj rysunek powoli od i do oka. W odległości ok. 15-20 cm ciemne koło przestaje być widoczne.
wynika, że najwrażliwszym miejscem na barwy jest dołek środkowy plamki, natomiast o zmroku jest to najmniej wrażliwe miejsce siatkówki. W pręcikach gromadzi się barwnik, zwany rodopsyną, który podnosi wrażliwość tych komórek na światło. Rodopsyna syntetyzuje się w ciemności, natomiast na świetle się rozkłada. Siatkówkę można oglądać u osobnika żywego przy użyciu przyrządu zwanego oftalmoskopem, złożonego z urządzenia oświetlającego i soczewki. Badanie za pomocą oftalmoskopu pozwala na obejrzenie tarczy nerwu wzrokowego, plamki i rozgałęzień naczyń tętnicy i żyły środkowej siatkówki (tętnicy i żyły), co ma duże znaczenie diagnostyczne w niektórych chorobach.
Nerw wzrokowy N e r w w z r o k o w y ( nervus opticuś) tworzy się z włókien nerwowych warstwy zwojowej siatkówki, które wychodzą z gałki ocznej przez otworki w blaszce sitowej twardówki. Rozwojowo jest on częścią mózgowia i dlatego zawiera wypustki opon mózgowych, które go okrywają na jego przebiegu wewnątrzoczodołowym od gałki ocznej do kanału wzrokowego. Na t y m odcinku nerw jest wygięty, dzięki czemu ma większą długość, a więc nie napina się przy ruchach gałki ocznej i tych ruchów nie hamuje Po przejściu przez kanał wzrokowy do jamy czaszki nerwy biegną na podstawie mózgu, kierując się ku płaszczyźnie pośrodkowej, w której tworzą skrzyżowanie n e r w ó w w z r o k o w y c h (chiasma nervorum opticorum). Krzyżując się każdy z n e r w ó w oddaje na stronę przeciwną tylko połowę włókien. Jest to więc skrzyżowanie nie pełne, ale tylko częściowe (ryc. 219). Od skrzyżowania wzrokowego odchodzą biegnące rozbieżnie ku tyłowi pęczki, prowadzące połowę włókien każdego z n e r w ó w wzrokowych. Razem ze zmianą składu włókien zmieniła się też nazwa tych t w o r ó w . Noszą one nazwę pasm w z r o k o w y c h (tractus optici), prawego i lewego, które wchodzą w skład drogi wzrokowej.
Zawartość gatki ocznej Gałka oczna, jak w i e m y , jest wypełniona ciałem szklistym i cieczą wodnistą. Wnętrze jej nie stanowi jednej wspólnej przestrzeni. W części przedniej gałki, ograniczonej od przodu rogówką, mieści się komora przednia oka (camera oculi anteriot); wypełniona cieczą wodnistą, zamkniętą od tyłu soczewką '486
kory mózgowej
Ryc. 219. Schemat dróg wzrokowych.
i tęczówką. Z tyłu tęczkówki leży komora tylna oka, wypełniona - podobnie jak przednia - cieczą wodnistą. Pozostałą, największą część gałki ocznej ( 4 / 5 ) wypełnia ciało szkliste. Wymienione przestrzenie nie są od siebie odcięte - do wszystkich dochodzi ciecz wodnista, zapewniająca jednolite ciśnienie w całej gałce ocznej.
Komora przednia gałki ocznej
i
i !
Oprócz wymienionych wyżej ścian stałych komora przednia ma jeszcze ścianę tylną w pewnym stopniu ruchomą. W skład jej wchodzi tęczówka i rogówka, które mogą się bardziej lub mniej wpuklać do komory przedniej 487
i wpływać na głębokość tej komory. Głębokość jej, czyli w y m i a r przednio-tylny, nie jest stały i zależny od budowy oka i jego stanu fizjologicznego. U dalekowidzów komora jest płytsza, natomiast głębsza u krótkowidzów. W komorze przedniej, w miejscu zetknięcia się rogówki, twardówki, ciała rzęskowego i tęczówki, zwanym kątem tęczówkowo-rogówkowym, leży tzw. więzadło grzebieniaste, złożone z pasm i błon łącznotkankowych. Stąd ciecz wodnista przesącza się do wspomnianej wyżej zatoki żylnej twardówki. Jest to droga o d p ł y w u tej cieczy, którą wytwarza tęczówka i ciało rzęskowe. Ciecz w o d n i s t a , której w komorze przedniej mieści się ok. 125 cm 3 , jest w ciągłym ruchu, gdyż jej warstwa, stykająca się z rogówką, oziębia się i opada ku dołowi jako cięższa; natomiastwarstwa leżąca bezpośrednio przy tęczówce zostaje przez nią ogrzana i jako lżejsza przechodzi do najwyższych części komory. Składa się ona głównie z w o d y z domieszką białka i chlorku sodowego.
Komora tylna gałki ocznej Komora tylna ( camera oculi posterior) jest to przestrzeń między tęczówką i ciałem szklistym. Środek jej zajmuje soczewka. W komorze tej leży układ delikatnych włókienek, łączących ciało rzęskowe z soczewką. Komora tylna jest wypełniona cieczą wodnistą. Łączy się ona z komorą przednią przez szczelinę między soczewką i brzegiem źrenicznym tęczówki oraz drogą przestrzeni kąta tęczówkowo-rogówkowego.
Soczewka Jeżeli w zamkniętej szczelnie skrzynce zrobimy mały otwór w jednej ze ścian, to w jej wnętrzu na ścianie przeciwległej zarysuje się zmniejszony odwrócony obraz przedmiotu, umieszczonego przed skrzynką. Ta właściwość była znana d a w n y m fizykom, nie można było jednak zastosować podobnego urządzenia do fotografii, gdyż przy bardzo m a ł y m otworze ilość światła wpadającego do wnętrza aparatu była tak mała, że czas naświetlania kliszy byłby bardzo długi. Przy powiększeniu o t w o r u powstawał obraz zamazany, nieostry, gdyż dostawało się do wnętrza skrzynki dużo promieni brzeżnych, nie skupiających się należycie. Wady te zostały usunięte przez wstawienie do otworu w skrzynce soczewki skupiającej. W oku, które już porównywaliśmy z aparatem fotograficznym, istnieje również soczewka ( lens ), zbudowana z przeźroczystych włókien tkanki łącznej. Jest to soczewka dwuwypukła, której powierzchnia przednia ma większy promień krzywizny, to znaczy, że jest mniej wypukła od powierzchni tylnej. Średnica soczewki mierzy około 9 m m , grubość jest zmienna i w y n o s i średnio 4 m m . Kształt soczewki zmienia się również z wiekiem: u noworodka jest ona prawie kulista, u dorosłego staje się mniej wypukła, u osobników starszych jeszcze bardziej spłaszczona. Poza t y m kształt jej zmienia się w zależności od '488
odległości oglądanych przedmiotów, o czym będzie m o w a w rozdziale o akomodacji. Te zmiany kształtu są możliwe dzięki specjalnej budowie soczewki. Jej warstwy zewnętrzne, zwane korą soczewki, są miękkie, jedynie leżące wewnątrz niej jądro soczewki jest twardsze. U ludzi młodych jądro jest małe i miękkie. Powiększa się ono z biegiem lat, a w wieku starczym stwardnienie obejmuje całą soczewkę. Czynnościowe zmiany kształtu soczewki zależą od otaczającej ją torebki, która stanowi cienką, bardzo sprężystą błonę. Na obwodzie jest ona grubsza dzięki wzmacniającym ją pasmom okrężnym na powierzchni przedniej i tylnej, do których przyczepiają się liczne cienkie włókienka obwódkowe. Łączą one torebkę soczewki z wyrostkami rzęskowymi, tworząc o b w ó d k ę rzęskową (zonula ciliaris), na której soczewka jest zawieszona. W wieku starczym może nastąpić różnego stopnia zmętnienie soczewki, zwane zaćmą (kataraktą). Usunięcie zmętniałej soczewki przywraca zdolność widzenia przy zastąpieniu jej czynności przez użycie odpowiednich szkieł (okulary).
Ciało szkliste Ciało szkliste ( c o r p u s vitreum) leży do tyłu od soczewki, wypełniając resztę gałki ocznej. W przedniej części ciała szklistego znajduje się wgłębienie, zwane dołem ciała szklistego, które wypełnia soczewka. Jak w s p o m n i a n o wyżej, jest to twór galaretowaty, przeźroczysty, silnie łamiący światło. Wewnątrz ciała szklistego drobne włókienka łącznotkankowe tworzą zrąb (stroma). Między włókienkami znajduje się ciecz szklista ( humor vitreus), która ma znaczenie dla utrzymania ciśnienia śródocznego. Skład jej jest podobny do cieczy wodnistej. Ciało szkliste zawiera 98,5 % w o d y i nieco białka, które tworzy jego zrąb. Wewnątrz ciała szklistego znajduje się kanał ciała szklistego o średnicy 1 - 2 m m , w którym w życiu p ł o d o w y m przebiega tętnica środkowa siatkówki (a. centralis retinae), kierująca się od tarczy nerwu wzrokowego do soczewki. W czasie rozwoju płodowego wstępuje ona wraz z towarzyszącą żyłą do nerwu wzrokowego i, wchodząc razem z nerwem do gałki ocznej, rozgałęzia się w siatkówce. Jeszcze u płodu tętnica ta zanika i nie przeszkadza w czynnościach soczewki i ciała szklistego. Wspomniany kanał jest wypełniony cieczą szklistą i delikatnymi włókienkami.
Czynności optyczne gałki ocznej Promienie światła, wchodzące do wnętrza gałki ocznej, przechodzą przez skomplikowany aparat optyczny, w którym zachodzą procesy ich załamania. Promienie trafiają do ośrodków o różnych właściwościach optycznych, załamując się przy każdym takim przejściu. Pewna część promieni zostaje odbita '489
Gałka oczna prawidłowa Soczewka dwuwklęsła
Korekcja
Soczewka dwuwypukła
Gałka oczna prawidłowa
Soczewka cylindryczna
Korekcja Ryc. 220. A. Krótkowzroczność (gałka oczna za długa). Korekcja za pomocą soczewki dwuwklęsłej. Ryc. 220. B. Nadwzroczność (gałka oczna za krótka). Korekcja za pomocą soczewki dwuwypukłej. Ryc. 220. C. Astygmatyzm (nieprawidłowa krzywizna rogówki lub soczewki). Korekcja za pomocą odpowiedniej soczewki cylindrycznej.
od powierzchni rogówki, jak również od soczewki i te pominiemy w naszych rozważaniach. Omówimy tu promienie dochodzące do siatkówki. Promienie dochodzące do oka załamują się w wielu miejscach: na przedniej powierzchni rogówki, dalej na tylnej, następnie przy przejściu do ośrodka odmiennego optycznie, jak ciecz wodnista, dalej na obu powierzchniach soczewki o różnych promieniach krzywizny, wreszcie na siatkówce. Nie możemy zająć się tymi skomplikowanymi zjawiskami, a dla ich zrozumienia, przynajmniej w ogólnych zarysach, skorzystamy ze schematu tzw. oka zredukowanego, nie uwzględniającego załamania wewnątrz soczewek i przy przechodzeniu promienia przez różne ośrodki optyczne. Przyjmiemy jedynie załamanie się światła w soczewce dwuwypukłej (ryc. 220). Już na podstawie tego uproszczonego schematu możemy zrozumieć zasad'490
nicze wady wzroku, jak krótkowzroczność i nadwzroczność. Jeżeli gałka oczna jest za krótka lub za długa, obraz będzie nie ostry, gdyż nie będzie leżał na siatkówce, lecz za nią lub przed nią. Dopiero ustawienie przed okiem odpowiedniej soczewki spowoduje powstanie ostrego obrazu na siatkówce. Przy krótkowzroczności należy użyć soczewki dwuwklęsłej (pomniejszającej), zaś przy nadwzroczności - dwuwypukłej (powiększającej). Nieostrość obrazu może zależeć nie tylko od wymiarów gałki ocznej, lecz również od niewydolności akomodacyjnej oka.
Akomodacja Wiemy, że oko męczy się przy dłuższym oglądaniu przedmiotów bliskich, np. przy pracy ręcznej, jak haftowanie, przy czytaniu, zwłaszcza drobnego druku, przy kreśleniu itp. W t y c h warunkach musimy co pewien czas przerwać pracę i zatrzymać wzrok na przedmiotach odległych. Mówimy, że wtedy oczy odpoczywają. Tak jest w rzeczywistości. Soczewka oka, jako ciało sprężyste, w stanie spokoju (np. we śnie) zachowuje swój określony kształt. Ten kształt zachowuje ona przy oglądaniu przedmiotów odległych, wysyłających, praktycznie biorąc, promienie równoległe, które soczewka oka skupia i rzuca na siatkówkę. Jeżeli jednak oglądamy przedmioty bliskie, to w układzie optycznym muszą zajść pewne zmiany. W aparacie fotograficznym, w którym ostrość nastawiona jest na „nieskończoność", tzn. na przedmioty odległe, do zdjęć przedmiotów bliskich należy albo zmienić obiektyw (czego na ogół się nie robi), albo przesunąć go do przodu, powiększając odległość między nim a kliszą. Te same zasady obowiązują i w oku. Jednak nie możemy przesunąć soczewki, która jest obiektywem oka, jak to robią pewne gatunki ryb. Nie możemy również zamienić soczewki, ale konstrukcja oka wyprzedziła konstrukcję aparatów fotograficznych. Otóż soczewka oka może zmienić swą odległość ogniskową przez zmianę swego kształtu, a dokładniej - stopnia swej wypukłości. Dzieje się to dzięki ciału rzęskowemu, na wyrostkach którego zawieszona jest soczewka za pomocą obwódki rzęskowej. Jeżeli mięsień rzęskowy nie jest napięty, to włókna tej obwódki napinają torebkę soczewki, dzięki czemu staje się ona bardziej płaska i nastawiona na wyraźne tworzenie obrazów przedmiotów oddalonych. Jeżeli teraz skierujemy wzrok na przedmiot bliski, to odruchowo napina się mięsień rzęskowy, zmniejszając napięcie włókien obwódki rzęskowej, a t y m samym napięcie torebki soczewki. Elastyczna soczewka wraca do swego pierwotnego, bardziej wypukłego kształtu. Soczewka jest teraz nastawiona na przedmioty bliższe i oko rozróżnia drobniejsze szczegóły oglądanego obrazu. Należy więc pamiętać, że widzimy wyraźnie przedmioty bliższe przy skurczu mięśnia rzęskowego, co jest powodem zmęczenia się wzroku. Natomiast rozkurcz tego mięśnia powoduje nastawienie soczewki na przedmioty dalsze. '491
Stąd też skierowanie wzroku w dal daje wypoczynek mięśniowi rzęskowemu, a t y m samym oku. Zjawisko przystosowania się oka do oglądania przedmiotów zarówno dalekich, jaki bliskich nosi nazwę akomodacji, która ma jednak swoje granice nie możemy np. odczytać druku przy nadmiernym przybliżeniu książki do oczu.
Adaptacja W i e m y , że np. rentgenolog, przed oglądaniem obrazu na ekranie, powinien kilka m i n u t (5-10) spędzić w ciemności, aby przygotować wzrok do tej czynności. W t y m czasie czułość jego wzroku na światło wzrasta dziesięciokrotnie i pozwala na dokładniejsze odczytanie obrazu. Zjawisko to nosi nazwę adaptacji wzroku. Po przebyciu w ciemności około 40 min wrażliwość oczu na światło wzrasta około 10 000 razy. W i e m y z własnego doświadczenia, że „przyzwyczajamy się" do ciemności i po p e w n y m czasie zaczynamy za pomocą wzroku orientować się w otoczeniu.
Naczynia i nerwy gałki ocznej Tętnice gałki ocznej są rozgałęzieniami tętnicy ocznej (a. opthalmica), która odchodzi od tętnicy szyjnej wewnętrznej. Są to tętnice rzęskowe przednia i tylna, które odgałęziają się na drobniejsze naczynia, tworzące połączenia wewnątrz gałki. Drugie źródło krwi tętniczej stanowi wspomniana wyżej tętnica środkowa siatkówki, przebiegająca wewnątrz nerwu wzrokowego i rozgałęziająca się w siatkówce. Żyły gałki ocznej biegną również dwiema drogami. Krew żylna z siatkówki płynie naczyniami, tworzącymi po połączeniu się żyłę środkową siatkówki, których przebieg można prześledzić przy badaniu dna oka, podobnie jak gałęzie tętnicze. Żyła ta wstępuje do nerwu wzrokowego, towarzysząc tętnicy środkowej siatkówki. Żyły błony naczyniowej tworzą wspomniane żyły wirowate, które przebijają twardówkę i wychodzą z gałki do tyłu od równika, uchodząc do żyły ocznej. Ciśnienie w żyłach gałki ocznej musi być zestrojone z ciśnieniem śródocznym, gdyż - jak w i e m y - ciecz wodnista odpływa do układu żylnego. W przec i w n y m wypadku doszłoby do poważnych zaburzeń krążenia krwi. Ciśnienie śródoczne jest wyższe od ciśnienia śródczaszkowego, dlatego też przy spadku ciśnienia krwi zaburzenia wzrokowe („ciemno w oczach") występują wcześniej niż utrata świadomości. Przerwanie d o p ł y w u krwi tętniczej do gałki ocznej, trwające do 20 min, nie powoduje jej uszkodzeń, natomiast po 40 min mogą powstać zmiany nieodwracalne. Nerwy gałki ocznej prowadzą włókna czuciowe od nerwu ocznego (gałąź n. '492
trójdzielnego), włókna ruchowe do mięśni gałki - od III pary nerwów czaszkowych (n. okoruchowy). Włókna autonomiczne do mięśni gładkich biegną w składzie n. okoru.chowego ze zwoju rzęskowego (m. rzęskowy i zwieracz źrenicy zaopatrują włókna przywspółczulne, m. rozszerzacz źrenicy - współczulne).
Narządy dodatkowe oka Należą do nich: aparat ruchowy gałki ocznej, aparat ochronny i narząd łzowy. Aparat ruchowy składa się z siedmiu mięśni, obficie unerwionych. Dzięki niemu wzrok może być bardzo szybko i dokładnie kierowany w dowolnym kierunku w jego zasięgu lub ustalany na obserwowanym ruchomym przedmiocie. Aparat ochronny, jak wiemy, chroni gałkę oczną przed urazami mechanicznymi (kurz, dym), termicznymi lub nadmiarem światła. Narząd łzowy chroni gałkę przed wysychaniem, jak również powoduje oczyszczanie rogówki i spojówki. W przypadku uszkodzenia lub zaburzenia czynności któregokolwiek z tych aparatów cierpi cały narząd wzroku.
Aparat ruchowy gałki ocznej Aparat ten stanowią mięśnie poprzecznie prążkowane gałki ocznej, zależne od naszej woli. Są to cztery mięśnie proste i dwa skośne. Mięśnie proste, przyczepione do gałki ku przodowi od równika, zbiegają się ku tyłowi, tworząc dokoła ujścia kanału nerwu wzrokowego na tylnej ścianie oczodołu pierścień ścięgnisty wspólny. Noszą one nazwy związane z ich położeniem - mięsień prosty górny, dolny, boczny i przyśrodkowy. Obracają one gałkę oczną każdy w swoją stronę, a więc boczny odwodzi biegun przedni gałki, przyśrodkowy przywodzi, górny podnosi, dolny obniża, kierując odpowiednio wzrok. Dwa mięśnie skośne przebiegają nieco odmiennie. Mięsień skośny górny, najdłuższy z mięśni oka, biegnie od pierścienia ścięgnistego wspólnego ku przodowi i przyśrodkowo do bloczka chrząstkowego, umocowanego na przyśrodkowej ścianie oczodołu. Jego cienkie ścięgno przechodzi przez bloczek, zagina się ku bokowi i tyłowi i przyczepia się do ściany gałki ocznej do tyłu od równika. Kurcząc się mięsień ten obraca biegun górny gałki w stronę nosa i obniża nieco biegun przedni. Mięsień skośny dolny, najkrótszy, rozpoczyna się na grzebieniu łzowym tylnym u dołu, biegnie prawie poprzecznie pod mięśniem prostym dolnym i przyczepia się do bocznej strony gałki, do tyłu od równika. Przy skurczu obraca biegun górny gałki ku bokowi, zaś unosi i odwodzi biegun przedni. W oczodole leży jeszcze u góry siódmy mięsień poprzecznie prążkowany, zwany dźwigaczem powieki górnej. Rozpoczyna się razem z innymi na pierścieniu ścięgnistym wspólnym i biegnie ponad mięśniem prostym górnym. Płaskim ścięgnem kończy się w skórze powieki górnej i na brzegu tarczki powiekowej. Mięsień ten podnosi powiekę i rozszerza szparę oczną. '493
Mięśnie proste górny i dolny, mięśnie skośne i dźwigacz powieki górnej są unerwione przez III nerw czaszkowy okoruchowy, mięsień prosty przyśrodkow y - przez nerw IV bloczkowy, wreszcie mięsień prosty boczny-przez VI nerw czaszkowy odwodzący. Dzięki specjalnej koordynacji nerwowej ruchy obu gałek ocznych są sprzężone, np. przy skierowaniu wzroku w lewo działają mięśnie prosty boczny lewy, unerwiony przez VI nerw czaszkowy, prosty przyśrodkowy lewy (unerw i o n y z III n. czaszkowego). Przy spojrzeniu w górę działają obustronnie mięśnie proste górne i dźwigacze powieki górnej (unerwione z III n. czaszkowego) itd. Przy porażeniu nerwu VI następuje zez zbieżny (strabismus convergens) na skutek napięcia stałego mięśnia prostego przyśrodkowego antagonisty porażonego mięśnia prostego bocznego. Odwrotnie przy porażeniu mięśnia prostego przyśrodkowego występuje zez rozbieżny( strabismus divergens).
Aparat ochronny gałki ocznej Na aparat ochronny składają się powieki i spojówka. Powieki górna i dolna ( palpebrae superior et interior) stanowią dwa elastyczne fałdy, które mogą się zwierać i zamykać szparę oczną lub się rozszerzać, odsłaniając gałkę oczną. Szkielet ich składa się z tarczek ( tarsus), zbudowanych z tkanki łącznej i chrząstki włóknistej. Dzięki temu powieki utrzymują swój kształt i sprężystość, która pozwala na właściwe im ruchy. Również dzięki tarczkom możemy powieki wywinąć, zwłaszcza górną. Jak wspomniano wyżej, powieki są zaopatrzone w mięśnie - górna ma własny dźwigacz powieki; poza t y m obie tarczki na powierzchni przedniej są przykryte częścią powiekową mięśnia okrężnego oka (m. orbicularis oculi), który zwiera powieki. Powieka dolna opada na parę m i l i m e t r ó w pod w p ł y w e m własnego ciężaru, mogą ją również nieco obniżać włókna mięśni prostego i skośnego dolnego, które dochodzą do brzegu dolnego tarczki. Powierzchnia zewnętrzna powiek jest pokryta cienką skórą. W tarczkach powiekowych leżą gruczoły tarczkowe ( glandulae tarsales), których przewody uchodzą na brzegu rzęskowym powiek. Gruczoły te są ułożone prostopadle do brzegu. Liczba ich w powiece górnej wynosi ok. 35. Są to gruczoły łojowe, których wydzielina natłuszcza brzeg powiek i nie pozwala na wypływanie cieczy łzowej przez ich brzegi. Powieka górna jest większa od dolnej. Obie powieki ograniczają szparę powiek (r/ma palpeprarum), ograniczoną kątami oka ( anguli oculi) bocznym i przyśrodkowym. Na brzegach powiek osadzone są rzęsy (cilia). Są to krótkie sztywne włosy, lekko wygięte na powiece górnej i nieco silniej na dolnej. Liczba ich wynosi na powiece górnej 150-200, na dolnej o połowę mniej. Nad powieką górną wyrastają b r w i (supercilia), różnego kształtu, gęstości i barwy. Kształt ich i układ w ł o s ó w należą do cech dziedzicznych. '494
NJ.
Spojówka Powierzchnia tylna powiek jest pokryta cienką błoną śluzową zwaną spoj ó w k ą ( tunica conjunctiva). Błona ta dochodzi do podstawy powiek, gdzie przechodzi na gałkę oczną, okrywając ją aż do brzegu rogówki. Stosownie do tego odróżniamy spojówkę: a) powiek, b) sklepienia i c) gałki ocznej. Szczelina Włosowata zawarta między powiekami i gałką oczną stanowi w o r e k s p o j ó w k o w y (saccus conjunctive). W kącie przyśrodkowym oka brzegi powiek otaczają zagłębienie zwane jeziorkiem ł z o w y m (lacus lacrimalis), w którym leży różowo zabarwione wzniesienie zwane mięskiem łzowym.
Narząd łzowy Jak już wspomniano, stykająca się z otwartym powietrzem rogówka jest przede wszystkim narażona na wysychanie, a poza t y m na zanieczyszczenie cząsteczkami kurzu oraz na urazy przez drobne ciała obce. Narząd łzowy ma więc za zadanie stałó zwilżanie rogówki i opłukiwanie jej z drobnych zanieczyszczeń. Składa się on głównie z gruczołu łzowego, wytwarzającego płyn łzowy i dróg łzowych, odprowadzających ciecz, która spełniła swe zadanie, z rowka spojówkowego do jamy nosa. Gruczoł ł z o w y Gruczoł ł z o w y ( g l a n d u l a lacrimalis), o wymiarach 2 0 x 1 2 m m , barwy różowoszarej, leży przy stropie oczodołu w jego bocznym przednim kącie, wypełniając dół tej samej nazwy. Ścięgno mięśnia dźwigacza powieki górnej dzieli go na dwie części: większą górną oczodołową, i mniejszą dolną, powiekową. Ciecz łzowa (humor lacrimalis), uchodzi kilkoma przewodzikami do sklepienia górnego spojówki, skąd spływa ku dołowi do worka spojówkowego i do kąta przyśrodkowego oka. W normalnych warunkach ciecz łzowa jest wydzielana jedynie przy otwartych powiekach i w czasie snu gruczoł łzowy jest nieczynny. Ogólna ilość wydzielonej w ciągu doby cieczy w y n o s i około 0,5 cm 3 . Zwiększa się ona przy podrażnieniach, działających na rogówkę, spojówkę czy siatkówkę (ciała obce, wiatr, gazy drażniące, urazy mechaniczne, rażące światło i in.), które wywołują łzawienie, występujące również w pewnych stanach psychicznych, jak np. wzruszenie. W tych wypadkach drogi odprowadzające mogą nie wystarczyć do odpływu cieczy łzowej, która przelewa się przez brzeg powieki dolnej w postaci łez ( l a c r i m a e ) . Ciecz łzowa składa się z w o d y z domieszką około 1% chlorku sodowego, 0,6% białka oraz drobnych ilości innych domieszek. Wydzielahie nadmierne cieczy łzowej występuje pod w p ł y w e m podniet, biegnących drogą nerwu twarzowego (VII para). U n o w o r o d k ó w zaopatrzenie nerwowe nie jest dostatecznie wykształcone i dlatego noworodek właściwie nie płacze, a wydaje jedynie krzyk bez łez. '495
Drogi ł z o w e Jak już w i e m y , płyn łzowy częściowo w y p a r o w u j e z powierzchni rogówki, a reszta jego spływa do tzw. jeziorka łzowego. Na brzegach każdej powieki, w pobliżu kąta przyśrodkowego oka, można dojrzeć drobny punkt ł z o w y (punctum lacrimale). Od punktów łzowych, które leżą na małych wzniesieniach, zwanych brodawkami łzowymi, biegną kanaliki łzowe - górny i dolny. Biegną one początkowo pionowo, a następnie zaginają się w kierunku przyśrodkowym (w stronę nosa) i przechodzą w część poziomą. Długość ich wynosi 8 - 9 m m , po czym uchodzą one do zbiornika, zwanego w o r k i e m ł z o w y m ( saccus lacrimalis), który leży we w ł a s n y m dole na przyśrodkowej ścianie oczodołu między grzebieniami ł z o w y m i przednim i tylnym. Worek łzowy przechodzi ku dołowi w przewód n o s o w o - ł z o w y (ductus nasolacrimalis), który otwiera się w przewodzie nosowym dolnym. Przechodzenie cieczy łzowej przez opisane drogi oparte jest prawdopodobnie na następujących zasadach. Punkty łzowe przy zamkniętych powiekach są zanurzone w cieczy łzowej jeziorka. Płyn wchodzi do nich na zasadzie włoskowatości. Przy otwieraniu powiek następuje wessanie płynu łzowego do części pionowej kanalików łzowych z p o w o d u ich lekkiego rozszerzenia. Przy zamykaniu powiek następuje wyciśnięcie płynu z kanalików do worka łzowego. Za t y m mechanizmem przemawia fakt, że przy występującym łzawieniu szybkie mruganie powiek powoduje opróżnienie rowka spojówkowego. Obrzęk błony śluzowej nosa utrudnia o d p ł y w cieczy łzowej do jamy nosowej. Odczuwamy to przy katarze, kiedy występuje łzawienie, pobudzone również podrażnieniem gruczołu łzowego na drodze odruchowej.
POWŁOKA W S P Ó L N A
Zarodek we wczesnych stadiach rozwoju jest pokryty komórkami zewnętrznego listka zarodkowego, czyli ektodermą, która u organizmów pierwotnych stanowi powłokę zewnętrzną ciała. U człowieka taka powłoka nie wystarcza przede wszystkim ze względu na jej delikatność i małą wytrzymałość na czynniki mechaniczne. Już u płodu do ektodermalnej powłoki dochodzi od wewnątrz silna warstwa łącznotkankowa pochodząca z mezodermy, tworząc ostateczną powłokę ciała - skórę ( cutis ). Jej warstwę zewnętrzną nabłonkową nazywamy naskórkiem (epidermis), a wewnętrzną łącznotkankową - skórą właściwą (dermis albo corium). Pod skórą właściwą leży tkanka podskórna (tela subcutanea) i podściółka tłuszczowa (panniculus adiposus). Zbudowana w ten sposób skóra chroni głębsze narządy przed urazami mechanicznymi, a przesuwając się dzięki podłożu z tkanki łącznej amortyzuje te urazy i pozwala na swobodne ruchy części ciała. Chroni ona również warstwy głębsze przed wysychaniem i nie dopuszcza do wtargnięcia drobnoustrojów czy trucizn do ustroju. Osłania również ustrój przed promieniami ultrafioletowymi i spełnia rolę narządu termoregulacji, o czym będzie m o w a niżej. Poza ochroną głębszych narządów skóra jest również narządem wydalania. Dzięki zakończeniom n e r w o w y m skóra jest też narządem zmysłu dotyku, bólu i temperatury. Powierzchnię skóry człowieka dorosłego ocenia się w zależności od wzrostu i b u d o w y na 1,5-1,8 m 2 . BUDOWA OGÓLNA SKÓRY Cechy skóry poznajemy przy jej oglądaniu i badaniu dotykiem. Najbardziej widoczną jej cechą jest barwa, którą łatwo dostrzegamy przy pierwszych oględzinach osobnika. Barwa skóry jest cechą rasową. Jak w i a d o m o , rozróżniamy rasę białą, czarną i żółtą. Różnice barwy skóry zależą od ilości barwnika - melaniny, wytwarzanego w naskórku. Najmniej zasobna w melaninę jest skóra rasy białej. Nazwa ta jednak nie określa koloru skóry. Barwę białą ma ona jedynie u osobników pozbawionych zdolności wytwarzania barwnika - albinosów. Na ogół skóra ma barwę różowawobiałą lub różową u b l o n d y n ó w czy rudych, natomiast odcienie bardziej lub mniej śniade występują u szatynów i brunetów. Barwa ta w naszym klimacie nie jest stała i zależy od pory roku. W okresach większego nasłonecznienia skóra przybiera odcienie ciemniejsze, również ciemniejsza jest ona u mieszkańców okolic górskich i u ludzi pracujących na w o l n y m powietrzu (zwłaszcza rolników). Wytwarzanie większej ilości barwni32 - Anatomia i fizjologia
497
ka wiąże się z działaniem promieni pozafiołkowych, które przy znaczniejszym natężeniu mogą uszkodzić głębsze warstwy skóry. Znane są ogólnie wypadki oparzeń słonecznych u osób nie opalonych, które nieostrożnie poddają się działaniu słońca w górach czy nad morzem. U osobników słabo pigmentowanych w miesiącach letnich występuje nierównomierne skupienie barwnika w skórze twarzy pod postacią p i e g ó w (ephelides). Zabarwienie skóry nie jest równomierne na całej powierzchni ciała. Skóra okryta ubraniem jest jaśniejsza. Poza t y m silniej zabarwiona jest skóra grzbietu, słabiej brzucha i piersi. Silnie pigmentowane są brodawki sutkowe, ciemniejsze zabarwienie ma skóra pach, okolica narządów płciowych i odbytu. W zabarwieniu skóry występują również różnice płciowe - skóra mężczyzn jest na ogół ciemniejsza. Również jaśniejszą skórę mają dzieci, u których ciemnieje ona z wiekiem. Podobnie jak skóra rasy białej nie jest biała, skóra osobników rasy czarnej nie jest czarna, tylko zabarwiona na brązowo w różnych odcieniach - od jasnego brązu przez kolor czekoladowy do bardzo ciemnegoNpłonie i podeszwy nie mają barwnika.
Naskórek
Przewód potowy -Skóra właściwa
Gruczoł łojowy M. przy włosow
Korzeń włosa
Gruczoł potow\ (•Tkanka podskórna z podściółką tłuszczową
Ryc. 221. Schemat budowy skóry.
'498
U rasy żółtej spotykamy barwy skóry od jasnej, jak u M o n g o ł ó w , do czerwonożółtej, jak u Indian amerykańskich. Zależnie od okolicy ciała skóra ma różną grubość i odporność. Najcieńsza jest na powiekach (około pół milimetra), średniej grubości na przedniej powierzchni tułowia, najgrubsza jest na dłoniach i podeszwach, a na grzbiecie dochodzi do kilku (7-8) milimetrów grubości. Grubość skóry można oceńić w przybliżeniu, ujmując ją w fałd kciukiem i palcem wskazującym. Grubość podściółki tłuszczowej zależy od stopnia odżywienia osobnika, od wieku, płci, okolicy ciała. Tak np. u kobiet grubsza na ogół podściółka odkłada się na kończynach, nadając im okrągłe kształty, u osobników starszych na brzuchu, u osesków mniej więcej równomiernie na całym ciele. Zdrowa skóra ma gładką, matową powierzchnię i jest na ogół jednolita, z wyjątkiem niektórych okolic. Oglądając skórę pod m a ł y m powiększeniem przez lupę widzimy na niej liczne małe pólka, oddzielone d r o b n y m i bruzdkami. Są to pólka skórne ( areae cutaneae) w kształcie r o m b ó w . Na powierzchni tych pólek przebiegają jeszcze drobniejsze bruzdki, dzięki którym powierzchnia skóry ma w y g l ą d matowy. Inaczej wygląda powierzchnia dłoni i podeszew. Widzimy tu wąskie listewki skórne, które przebiegają równolegle do siebie, układając się w określone wzory. Na opuszkach palców rąk tworzą one łuki, pętle i w i r y , złożone z różnej liczby listewek, biegnących w zmiennych kierunkach. Tworzą one niepowtarzalne linie, zwane papilarnymi, odmienne u każdego osobnika. Ta ich właściwość ma duże znaczenie w kryminalistyce, pozwalając identyfikować osoby na podstawie odcisków palców. Nauka o odciskach palców nosi nazwę daktyloskopii. Wynika z niej, że ogólny charakter odcisków daktyloskopowych jest dziedziczny, natomiast ich szczegóły zależą od w p ł y w ó w zewnętrznych. Dlatego też dzieci mają inne odciski palców niż ich rodzice. Jeżeli skóra jest przez dłuższy czas uciskana, to powstają na niej zmiany chorobowe, znane pod nazwą odleżyn ( decubitus ). Dlatego też obłożnie chorzy wymagają szczególnej pielęgnacji w celu zapobieżenia trudno gojącym się odleżynom. BUDOWA SZCZEGÓŁOWA SKÓRY Jak wspomniano, skóra składa się z naskórka i skóry właściwej. Warstwa zewnętrzna, naskórek ( e p i d e r m i s ) , jest zbudowany z nabłonka wielowarstwow e g o płaskiego. Jego warstwa podstawna składa się z wysokich komórek, których liczba zwiększa się przez podział. Nowo powstające komórki są w y s u w a n e ku powierzchni skóry i w miarę przesuwania się stają się coraz niższe i bardziej spłaszczone. Warstwa powierzchowna naskórka składa się z zupełnie płaskich płytek zrogowaciałych, które wreszcie złuszczają się i są zastępowane przez głębsze rogowaciejące komórki. Zrogowaciałe komórki naskórka stanowią jego warstwę ochronną. '499
Grubość naskórka na podeszwach stóp wynosi ok. 2 m m , na dłoniach około 1 mm i jest związana z rodzajem pracy osobnika. Przy ciężkiej pracy fizycznej naskórek tworzy znaczne zgrubienia w postaci odcisków. Na powiekach grubość naskórka jest bardzo mała (ok. 0,1 mm). Warstwa zrogowaciała zawiera substancję rogową, zwaną keratyną, która pod w p ł y w e m w o d y podlega zmacerowaniu, jak to widzimy przy dłuższym moczeniu rąk czy stóp, np. przy praniu lub w kąpieli. Skóra właściwa ( c o r i u m ) jest zbudowana głównie z włókien tkanki łącznej, które nadają jej odporność i elastyczność. W tej warstwie leżą też gruczoły skóry (p. dalej) oraz włosy i drobne mięśnie przywłosowe. Dzięki elastyczności tej warstwy skóra przylega do warstw głębszych, ściśle obejmując części ciała, np. szyję, t u ł ó w , jak to widzimy u dzieci i osobników młodych. W wieku starczym skóra właściwa traci swą sprężystość i zwłaszcza przy schudnięciu nie obejmuje ściśle pokrywanych części ciała, a tworzy fałdy, stanowiące jak gdyby jej nadmiar. Włókna skóry właściwej układają się pasmami w określonych kierunkach, powodując tworzenie tzw. linii napięć. Przebieg ich nie jest przypadkowy, odpowiada on kierunkom najsilniejszego napięcia skóry w stanie spokoju. Przebieg ich ma duże znaczenie praktyczne dla określania kierunku cięć skóry przy zabiegach operacyjnych. Przy przecięciu skóry wzdłuż linii napięć brzegi rany łatwo się łączą, natomiast przy przecięciu w poprzek rana zieje, co też widzi się przy ranach przypadkowych, powstających na skutek urazów. Skóra właściwa jest połączona z narządami głębiej leżącymi, jak kości, powięzie i in., tkanką podskórną ( tela subcutanea), która składa się z tkanki łącznej wiotkiej. Dzięki temu skóra może się przesuwać względem głębszych warstw ciała, nie krępując jego ruchów. Również dzięki ł a t w e m u przesuwaniu się skóry można przeszczepiać jej płaty, nie odcinając jednak ich całkowicie, a pozostawiając ich połączenie z resztą skóry szypułą zabezpieczającą dopływ krwi do przeszczepu. W tkance podskórnej znajdują się większe lub mniejsze skupienia komórek tłuszczowych, tworzących wspomnianą wyżej podściółkę tłuszczową ( panniculus adiposus). Miejsca, w których skupia się tkanka tłuszczowa, zależą od konstytucji, płci, wieku osobnika i działania jego gruczołów dokrewnych. Najmniej tkanki tłuszczowej (lub jej brak) występuje na powiekach, wargach i nosie, na prąciu i mosznie, łechtaczce i wargach sromowych mniejszych oraz na uszach. Obfita podściółka tłuszczowa występuje u kobiet na kończynach, pośladkach i klatce piersiowej. Tkanka tłuszczowa u obu płci łatwo odkłada się na brzuchu. Przy dobrze rozwiniętej podściółce tłuszczowej skóra jest dobrze napięta: Przy zaniku podściółki skóra wiotczeje i fałduje się, gdyż obkurczanie się skóry odbywa się bardzo w o l n o . Podobnie zachowuje się skóra przy szybkim odwodnieniu organizmu, co wybitnie występuje u osesków przy biegunkach. Odwrotnie przy zatrzymaniu w o d y w organizmie w tkance podskórnej występują obrzęki. Przy uciśnięciu palcem miejsca dotkniętego obrzękiem na skórze pozostaje zagłębienie, które powoli się wyrównuje. '500
Skóra ludzi młodych, jędrna i odporna, z biegiem lat zmienia swe właściwości. W wieku starczym następuje jej zanik, atrofia. Skóra wiotczeje, zmniejsza się jej napięcie, wskutek utraty wody. Jednocześnie wszystkie warstwy skóry stają się cieńsze, dzięki czemu nawet drobniejsze żyły przeświecają przez naskórek. Skóra staje się mniej elastyczna, p o m i m o zwiększenia się zarówno ilości, jak i grubości włókien sprężystych. Jak wspomniano, ilość barwnika zwiększa się i skóra przybiera kolor szarawożółty.
TWORY NABŁONKOWE SKÓRY Nabłonek nię tylko pokrywa powierzchnię skóry. Wchodzi on w jej głąb, tworząc szereg narządów pomocniczych, o różnych kształtach i czynnościach. Należą do nich: 1) gruczoły skóry, 2) włosy i 3) paznokcie.
Gruczoły skóry Gruczoły p o t o w e Jak w s p o m n i a n o wyżej, skóra spełnia m.in. zadania wydalania niektórych substancji, głównie w o d y . Są one wydalane z potem, który wytwarzają gruczoły p o t o w e ( g l a n d u l a e sudoriferae). Są to gruczoły cewkowe, przedstawiające cewki nabłonkowe, leżące w głębi skóry właściwej, zwinięte na końcu w kształcie kłębka, stąd też nazwane gruczołami kłębkowatymi. Gruczoły te występują zarówno na uwłosionej, jak i nie uwłosionej części skóry, brak ich jedynie na wargach s r o m o w y c h oraz na napletku i żołędzi prącia. Mniej ich jest na powiekach i na bocznej stronie małżowiny usznej. Liczba ich na centymetrze kwadratowym zależy od okolicy ciała - na karku i pośladkach w y n o s i ona od 50 do 100, na dłoniach i podeszwach od 300 do 400. W innych okolicach mieści się w granicach wymienionych liczb. Wydzieliną omawianych gruczołów jest p o t (sudot), który jest płynem złożonym głównie z w o d y (ok. 98%). Zawiera on ok. 0,8% soli kuchennej oraz pewne ilości mocznika kwasu moczowego i amoniaku. Wydzielanie potu odbywa się stale, jednak w małych ilościach, których zwykle nie dostrzegamy. Wydzielanie to wzmaga się pod w p ł y w e m wysokiej temperatury otoczenia i cięższej pracy fizycznej. Wydzielanie potu ma duże znaczenie dla utrzymania stałej temperatury ciała. Ciepło jest wytwarzane w organizmie w szeregu reakcji chemicznych egzotermicznych, czyli przebiegających z jego wydzielaniem, jak np. przy pracy mięśni. Przy wytężonej pracy fizycznej wytwarza się nadmiar ciepła, które nie zdąży wypromieniować przez skórę i może doprowadzić do przegrzania organizmu, a więc do stanu patologicznego. Jako reakcja obronna wzmaga się wydzielanie potu, który parując pod w p ł y w e m ciepła zabiera duże jego ilości ze skóry, co oziębia przepływającą w naczyniach skóry krew, a przez to cały organizm. '501
Dlatego też należy pamiętać, aby w lecie osesków nie Ubierać ani okrywać zbyt ciepło, jak również zapewnić ludziom ciężko pracującym dostateczną ilość płynów do picia. Hutnicy, pracujący w lecie w przegrzanych pomieszczeniach przy wytopie metali, wypijają dziennie po kilkanaście litrów płynów dla uzupełnienia w o d y , wydzielanej z potem. Poza gruczołami p o t o w y m i organizm traci w o d ę również przez parowanie jej bezpośrednio przez naskórek. Oprócz opisanych gruczołów potowych istnieją w skórze również duże gruczoły kłębkowate (apokrynowe), których wydzielina jest odmienna od potu oraz odznacza się swoistym zapachem, barwą i innymi właściwościami różnymi dla każdego osobnika. Człowiek zatracił zdolność ich odróżniania, natomiast zwięrzęta, jak np. psy, rozpoznają różnych osobników i ich rzeczy, kierując się zmysłem węchu. Gruczoły te występują w dole pachowym, w otoczce brodawki sutkowej, w okolicy odbytu i in., w większej ilości u osobników płci żeńskiej niż męskiej. Niektóre zwierzęta mają dużą ilość gruczołów potowych, np. koń, inne nie pocą się wcale z p o w o d u ich braku, jak np. pies, który w upał chłodzi się przez wywieszony z pyska język, ociekający zwilżającą go śliną.
Gruczoły ł o j o w e Naskórek, jak wspomniano, wysycha i traci miękkość. Jego warstwa zrogowaciała w y m a g a natłuszczania dla utrzymania elastyczności. Rolę tę pełnią gruczoły ł o j o w e (glandulae sebaceae), założone w skórze właściwej, przede wszystkim w najbliższym otoczeniu włosów. Są to gruczoły pęcherzykowe, wyprowadzające swą wydzielinę, zwaną łojem skórnym ( sebum) do lejka u podstawy włosa (p. dalej), skąd w y p ł y w a ona na powierzchnię skóry i ją namaszcza. Ogólna liczba gruczołów łojowych jest mniejsza niż potowych. Najwięcej ich występuje na owłosionej skórze głowy, na twarzy, na grzbiecie i klatce piersiowej, gdzie liczba ich jest zbliżona do liczby gruczołów potowych tych okolic. Na tułowiu i kończynach liczba ich stanowi 1 / 6 - 1 / 8 liczby gruczoł ó w potowych. Na dłoniach i podeszwach gruczoły ł o j o w e nie występują. Jak wspomniano, gruczoły ł o j o w e występują również na nie uwłosionej powierzchni skóry, jak np. na czerwieni wargowej. Wydzielina ich chroni wargi przed wysychaniem i pękaniem. Podobne gruczoły, pod nazwą gruczołów tarczkowych ( glandulae tarsales), są założone w obu powiekach, szczególnie w górnej. Wydzielina ich pokrywa brzeg rzęskowy powiek, zapobiegając w y p ł y w a n i u wydzieliny łzowej na zewnątrz - na skórę powiek i twarzy. Łój skórny zawiera w o l n e kwasy tłuszczowe, cholesterol i związki woskowe. W temperaturze ciała łój jest płynny, a zastyga przy temperaturze poniżej 30 stopni. Powleka on włosy i nadaje im miękkość. Przy jego nadmiarze dochodzi do tzw. ł o j o t o k u (seborrhea), powodującego powstawanie łupieżu.
'502
Gruczoły s u t k o w e Gruczoły sutkowe są największymi gruczołami skóry. Ze względu na ich ścisły związek czynnościowy z narządami płciowymi żeńskimi zostały one o m ó w i o n e w o d p o w i e d n i m rozdziale.
Włosy Włosy należą do t w o r ó w nabłonkowych skóry, podobnie jak sierść ssaków, pióra ptaków, łuski ryb czy tarczki niższych zwierząt. U człowieka utraciły one pierwotną rolę tworzenia płaszcza powietrznego, zatrzymującego ciepło. Owłosienie zachowało się u człowieka głównie na głowie, w dołach pachow y c h i w okolicy łonowej, natomiast w innych okolicach zachowało się jedynie jako owłosienie szczątkowe. W niektórych przypadkach owłosienie nadmierne (hypertrichosis) występuje na tułowiu czy na kończynach i świadczy o zaburzeniach czynności narządów wydzielania wewnętrznego, czyli gruczołów dokrewnych. Włosy pokrywają całe ciało z wyjątkiem dłoni i podeszew, powierzchni grzbietowej paliczków dalszych, żołędzi i prącia, wewnętrznej powierzchni napletka i w a r g sromowych. Ciało noworodka pokrywa meszek ( l a n u g o ) , który następnie zmienia się w drobne włoski. Już u noworodka występują rozmaicie rozwinięte w ł o s y g ł o w y ( c a p i l l i t i u m ) , b r w i ( s u p e r c i l i a ) i rzęsy (cilia), a w okresie pokwitania pojawiają się włosy wąsów i brody (barba). W wieku dorosłym i starczym pojawiają się włosy nozdrzy i przewodu usznego. Na t u ł o w i u w okresie pokwitania pojawiają się włosy pachy i w ł o s y łonowe. Jak widać już z rozwoju w ł o s ó w , ich pojawianie się jest związane z czynnością gruczołów dokrewnych. Na w p ł y w y hormonalne wskazuje również pojawianie się w m ł o d y m wieku łysiny przy dobrym zachowaniu owłosienia innych części ciała, co jest cechą wybitnie męską. Włos (pilus) ma część zewnętrzną, zwaną łodygą włosa, i ukryty w skórze korzeń włosa, którego zgrubiały, widzialny gołym okiem, koniec dolny nosi nazwę opuszki włosa. Opuszka jest osadzona na łącznotkankowej brodawce włosa, do której dochodzą naczynia krwionośne. Z komórek opuszki przez ich podział powstaje rosnący włos. Przy zniszczeniu opuszki włos ginie i już nie odrasta. Włos leży w zagłębieniu skóry, tworzącym kanał, zwany mieszkiem włosa, wysłany nabłonkiem. Kanał na powierzchni skóry tworzy lejkowate zagłębienie, do którego uchodzą przewody gruczołów łojowych. Korzeń włosa jest ustawiony skośnie do powierzchni skóry. Po stronie, w którą jest on pochylony, leży gruczoł ł o j o w y , a za nim pasemko włókien mięśniowych gładkich, tworzących mięsień przywłosowy. Skurcz tego mięśnia powoduje wyprostowanie włosa i wysadzenie go na zewnątrz, co jest przyczyną jeżenia się w ł o s ó w i powstawania „gęsiej skórki", o czym była m o w a przy opisie mięśniówki gładkiej. '503
Barwa w ł o s ó w jest cechą dziedziczną. Zależy ona od ilości ziarnistego barwnika brązowego i rozpuszczonego czerwonego, które w rozmaitych kombinacjach ilościowych dają szereg odcieni barwy w ł o s ó w . Silna koncentracja barwnika brązowego nadaje w ł o s o m kolor czarny, a występowanie jego tylko w warstwie zewnętrznej łodygi powoduje jasny kolor włosa. Przy utracie barwnika w ł o s y przybierają kolor popielaty, natomiast całkowicie bieleją na skutek występowania pęcherzyków powietrza między komórkami. Pod względem kształtu włosy dzielimy na proste, faliste i kędzierzawe. Włosy tworzą na głowie zazwyczaj jeden, a czasem dwa w i r y (vortexpilorum), które bywają prawo- lub lewoskrętne. Cechy te są dziedziczne. Włos żyje 3 - 5 lat. Po jego wypadnięciu wyrasta na jego miejscu n o w y o podobnych cechach. Rosnący włos w ciągu doby zwiększa się o 0,3-0,4 m m . Po okresie wzrostu następuje okres spoczynku. Jeżeli nowe włosy nie wyrastają, występuje łysina ( alopecia ). Włosy zwykle wypadają po ciężkich chorobach zakaźnych (dury). Liczba w ł o s ó w na głowie w y n o s i ok. 120 tysięcy.
Paznokcie Paznokieć ( u n g u i s ) jest w y t w o r e m nabłonkowym skóry, leżącym na dalszych opuszkach palców. Blaszka zrogowaciała paznokcia ochrania opuszkę palca. Paznokcie palców r ą k - podobnie jak i same p a l c e - s ą lepiej rozwinięte niż na stopach, gdzie dobrze rozwinięty paznokieć występuje tylko na palcu pierwszym - paluchu. Rola paznokci u człowieka jest zupełnie inna niż rola pazurów czy odpowiadających im kopyt u zwierząt. Są one związane z dotykiem; opuszka palca pozbawionego paznokcia jest mniej wrażliwa na dotyk. Kształty paznokci są dość urozmaicone. Płytka paznokcia jest wypukła w dwóch płaszczyznach - podłużnej i poprzecznej. Stopień wypukłości może być bardzo różny. Najbardziej widoczną cechą wymierną paznokcia jest jego szerokość w stosunku do długości. M ó w i m y o paznokciach średnio szerokich, jeżeli długość i szerokość płytki (bez wolnego brzegu) są równe. Dawniej przypuszczano, że szerokie paznokcie występują u osób pracujących fizycznie. Nowsze badania dowiodły, że wąskie paznokcie występują częściej u kobiet, szerokie u mężczyzn. Różnice te zaznaczają się u osobników dorosłych, podczas gdy przed okresem pokwitania nie są one uchwytne. Z tego wynika, że na kształt paznokci mają również w p ł y w h o r m o n y płciowe. Grubość paznokci jest różna i wynosi najwyżej niecałe 0,4 m m ; jest ona na ogół większa u mężczyzn niż u kobiet. Osoby pracujące fizycznie mają paznokcie grubsze. Paznokieć ma brzeg w o l n y , występujący przy końcu palca, oraz brzegi boczne i brzeg zakryty, wchodzący do zatoki paznokcia, ograniczony wyniosłością, zwaną w a ł e m paznokciowym, z którego na powierzchnię ciała paznokcia schodzi warstewka nabłonkowa, zwana obrąbkiem naskórkowym paznok'504
cia, której uszkodzenie przy pielęgnacji paznokcia może stać się przyczyną bolesnego zapalenia opuszki palca (zanokcica). Wspomniany brzeg zakryty paznokcia stanowi jego korzeń. Ciało i korzeń leżą na podłożu utworzonym ze skóry właściwej, zwanym macierzą paznokcia, z której też on powstaje. W macierzy biegną liczne naczynia włosowate, które przeświecając przez ciało paznokcia nadają mu barwę różową. Przy niedostatecznym natlenieniu krwi następuje sinica (caynosis) i barwa ta nabiera odcienia fioletowego. Część paznokcia od strony korzenia ma barwę białawą, stanowi obłączek, różnie wykształcony. Leży tu warstwa rozrodcza paznokcia, który w miarę nawarstwiania masy zrogowaciałej przesuwa się w kierunku końca palca. Szybkość wzrostu paznokcia wynosi średnio 1 mm na tydzień. Zależy ona od wieku,stanu zdrowia i innych w p ł y w ó w . Po 30 r. ż. paznokcie rosną wolniej. Po uszkodzeniu czy usunięciu paznokcia odrasta on w czasie około stu dni.
GRUCZOŁY DOKREWNE
Gruczołami d o k r e w n y m i (grandulae sine ductibus) nazywamy takie narządy gruczołowe, które nie mają przewodów wyprowadzających i oddają swą wydzielinę bezpośrednio do krwi. Substancje wydzielane przez te gruczoły nazywamy h o r m o n a m i ( h o r m a o pobudzam). Charakteryzują się one t y m , że działają w obszarach odległych od miejsca swego powstania, dokąd zostają przeniesione z krwią. Zadanie ich polega na regulowaniu i koordynowaniu czynności narządów oraz na utrzym y w a n i u stałości składu środowiska wewnętrznego. Pełnią one zatem rolę podobną do układu nerwowego, z którym zresztą pozostają w ścisłym związku czynnościowym. Różnica polega na t y m przede wszystkim, że regulacja nerwowa odbywa się znacznie szybciej i jest raczej krótkotrwała, natomiast regulacja hormonalna jest wolniejsza i działa dłużej. Jeżeli impulsy nerwowe przyrównamy do informacji lub rozkazów przesyłanych przez kable telefoniczne, to h o r m o n y bsędą podobne do posłańców wędrujących z prądem krwi. Gruczoły dokrewne są rozrzucone w różnych częściach ustroju i różnią się między sobą wielkością, kształtem i budową wewnętrzną. Gruczołami wspólnymi dla obu płci są: przysadka mózgowa, gruczoł tarczowy, gruczoły przytarczyczne, nadnercza i część wewnątrzwydzielnicza trzustki. Gruczoły płciowe, czyli gonady, są swoiste dla każdej płci - u kobiety są to jajniki, u mężczyzny jądra. Gruczoły płciowe pozostają nieczynne w dzieciństwie, tj. do okresu pokwitania. Grasica bywa tradycyjnie opisywana w rozdziale o gruczołach dokrewnych. Nie ma jednak, jak dotąd, d o w o d ó w na to, że jest ona istotnie gruczołem dokrewnym. Hormony są wytwarzane i działają w ilościach niezmiernie małych. Działanie ich jest swoiste, tj. poszczególne h o r m o n y wywierają swój w p ł y w tylko na pewne narządy, tkanki lub na pewne procesy zachodzące w ustroju. Pod względem chemicznym h o r m o n y można podzielić na: 1) pochodne fenolu (adrenalina, tyroksyna), 2) h o r m o n y białkowe (hormony przysadki, Insulina, parathormon) i 3) h o r m o n y sterydowe (hormony kory nadnerczy, h o r m o n y płciowe). ( Obecnie budowa chemiczna i właściwości fizjologiczne i farmakologiczne wiełu h o r m o n ó w są dokładnie poznane. Wyjaśnienie budowy chemicznej poszczególnych h o r m o n ó w pozwoliło nie tylko na ich syntezę, ale i na otrzymanie substancji podobnych o pożądanych właściwościach farmakologicznych. Badaniem właściwości i działania h o r m o n ó w zajmuje się specjalna gałąź nauki, zwana endokrynologią (endon - wewnątrz, crino- wydzielam). Postępy w tej dziedzinie należą do największych zdobyczy medycyny i biologii bieżącego stulecia. '506
Regulacja hormonalna
Regulacja nerwowa
Narząd Ryc. 222. Różnice między regulacją nerwową (szybką) i hormonalną (wolną).
PRZYSADKA Przysadka (hypophysis) jest nieparzystym gruczołem, ważącym ok. 0,5 g, kształtu owalnego, średnicy ok. 1 cm, położonym u podstawy mózgu w zagłębieniu siodła tureckiego. Przysadka łączy się z podstawą mózgu za pośrednict w e m lejka (infundibulum). Rozróżniamy w przysadce płat przedni (lobus anterior), zwany też adenohypophysis i płat t y l n y (lobus posterior), czyli neurohypophysis. Płat przedni, bardzo obficie unaczyniony, zbudowany jest ze zrębu łącznotkankowego, na którym spoczywają pasma komórek nabłonkowych. Rozróżniamy tu dobrze barwiące się komórki barwnikochłonne (chromofilne) oraz słabo barwiące się - komórki barwnikooporne (chromofobne), czyli podstawowe. Wśród komórek barwnikochłonnych jedne barwią się barwnikami zasadow y m i i noszą nazwę komórek zasadochłonnych, inne barwiące się barwnikami kwaśnymi nazywane są komórkami kwasochłonnymi. Płat t y l n y przysadki zbudowany jest z elementów neuroglejowych. U licznych zwierząt występuje między przednim i t y l n y m płatem wąska warstwa prawie zupełnie pozbawiona naczyń, zwana częścią pośrednią. U człowieka część ta jest bardzo słabo rozwinięta. '507
Bezpośrednie sąsiedztwo i łączność anatomiczna z mózgowiem ma swoje konsekwencje czynnościowe - przysadka stanowi o g n i w o łączące układ dokrewny z układem n e r w o w y m . Czynność przysadki pozostaje pod kontrolą sygnałów przekazywanych z podwzgórza międzymózgowia.
Mózgowie
Hormon wzrostowy
Hormony sterydowe nadnerczy
Tyroksyna
W
Estrogeny Progesteron
Ryc. 223. S c h e m a t i l u s t r u j ą c y c z y n n o ś c i w y d z i e l n i c z e p r z y s a d k i .
'508
Tylny płat przysadki wydziela dwa hormony: 1) hormon antydiuretyczny (ADH), regulujący gospodarkę wodną ustroju, i 2) oksytocynę, która bierze udział w wydzielaniu mleka podczas laktacji, wspomaga zapłodnienie komórki jajowej oraz odgrywa istotną rolę podczas porodu. Sekrecja tylnego płata pozostaje pod kontrolą podwzgórza za pośrednictwem włókien nerwowych łączących je z tą częścią mózgowia. Płat przedni przysadki produkuje co najmniej sześć hormonów, które pełnią ważną rolę w regulacji procesów metabolicznych ustroju (ryc. 223). Są to: 1) hormon wzrostowy, zwany także somatotropiną, powodujący wzrost wszystkich tkanek, które mają możność wzrastania, 2) ACTH, czyli hormon kortykot r o p o w y , 3) TSH, czyli hormontyreotropowy,4)folikulostymulina, 5) hormon luteinizujący i 6) hormon luteotropowy, prawdopodobnie identyczny z prolaktynę. Hormony przysadki regulujące czynność innych gruczołów dokrewnych noszą nazwę hormonów tropowych. Wydzielanie hormonów tylnego płata przysadki pozostaje pod kontrolą podwzgórza, która odbywa się jednak nie na drodze nerwowej, lecz za pośrednictwem pewnych substancji o charakterze polipeptydów, które są wytwarzane w podwzgórzu i przekazywane do przysadki drogą drobnych naczyń krwionośnych. Podwzgórze jest, jak wiemy, centralną stacją odbioru wszelkiego rodzaju bodźców czuciowych. Na poziomie podwzgórza następuje integracja wszystkich informacji napływających z receptorów (narządów zmysłów i interoreceptorów) i w zależności od aktualnych potrzeb odbywa się wydzielanie odpowiednich substancji, nazwanych czynnikami (hormonami) podwzgórza. Odróżnia się wśród nich.czynniki uwalniające hormony przysadkowe oraz czynniki hamujące uwalnianie, a prawdopodobnie także i syntezę hormonów przysadki. Hormon wzrostowy. Obecność w przysadce hormonu pobudzającego wzrost sugerowały od dawna obserwacje kliniczne: karłowatość w przypadkach zniszczenia przysadki przez proces chorobowy i gigantyzm - w przypadkach nowotworów wychodzących z jej aktywnych komórek. Wstrzykiwanie świeżych wyciągów przysadki młodym zdrowym zwierzętom przyspiesza ich wzrost. Drogą analiz chemicznych wyizolowano z przysadki czysty hormon wzrostowy, który jest białkiem. Działanie hormonu wzrostowego polega na tym, że powoduje on wykorzystanie znacznej części pobieranego pokarmu na budowę nowych tkanek. Wzrost ustroju, jak wiadomo, ulega zahamowaniu z chwilą osiągnięcia dojrzałości płciowej, kiedy to następuje zrośnięcie nasad i trzonów kości długich. Proces ten przyspieszają hormony płciowe, które z kolei pozostają pod kontrolą przysadki. Hormon wzrostowy wywołuje proporcjonalny wzrost wszystkich narządów ustroju. Nadmierne wydzielanie tego hormonu w ustroju dorosłym nie może wpłynąć na wzrost kości długich, powoduje jednak przerost i zniekształcenie kości na obwodzie - zespół chorobowy zwany akromegalią. '509
Za dużo
H
Karłowatość przysadkowa
Wzrost prawidłowy
Gigantyzm
Akromegalia
Ryc. 224. Schemat działania hormonu wzrostowego.
Przysadka reguluje wzrost młodego ustroju nie tylko poprzez swoisty hormon wzrostowy, lecz również pośrednio przez hormon tarczycy i gonad. Hormon tyreotropowy pobudza wzrost i czynność komórek w pęcherzykach ta rczycy. Brak tego hormonu może doprowadzić do niedoczynności tarczycy, podobnie jak nadmierne jego wydzielanie jest przyczyną nadczynności tego gruczołu. Wydzielanie hormonu tyreotropowego można zahamować podawaniem jodu. Na tym właśnie polega leczenie jodem w chorobie Basedowa, stosowane jako przygotowanie do zabiegu operacyjnego. ( Działanie hormonów gonadotropowych zostało dokładniej omówione przy opicie gruczołów płciowych. Hormon adrenokortykotropowy (ACTH) pobudza wydzielanie hormonów kory nadnerczy, o czym będzie mowa w rozdziale poświęconym nadnerczom. Płat tylny przysadki wydziela dwa hormony o podobnej budowie chemicznej, lecz odmiennym działaniu: 1) hormon antydiuretyczny (ADH), regulujący wydalanie wody z moczem oraz 2) oksytocynę, która odgrywa istotną rolę przy porodzie. Są to oktapeptydy, tj. związki złożone z ośmiu aminokwasów, przy tym jak to zaznaczono na ryc. 225, różnią się one między sobą tylko dwoma aminokwasami. Hormony te są obecnie wytwarzane syntetycznie. Wazopresyna, która bywa wymieniana jako hormon tylnego płata przysadki, jest substancją identyczną z hormonem antydiuretycznym. Nazwa pocho'510
a* Ryc. 225. Cząsteczka oksytocyny i ADH - oktapeptydy różniące się między sooą tylko dwoma aminokwasami (zaznaczonymi strzałką).
dzi stąd, że ADH w dużych dawkach powoduje skurcz naczyń włosowatych i wzrost ciśnienia krwi. H o r m o n antydiuretyczny (ADH) działa na nerki, powodując zatrzymanie w o d y i wydzielanie zagęszczonego moczu. ADH jest ważnym regulatorem bilansu w o d n e g o ustroju. Jeżeli dowóz w o d y ulegnie zmniejszeniu lub dojdzie do dużych strat w o d y , np. wskutek nadmiernego pocenia, wówczas następuje wzrost wydzielania ADH, który hamuje diureżę. Zwiększony dowóz w o d y powoduje zahamowanie wydzielania ADH i szybki wzrost diurezy. Brak h o r m o n u antydiuretycznego spowodowany zniszczeniem komórek tylnego płata przysadki, np. wskutek urazu, przez guz, a czasem z przyczyn nie znanych, jest przyczyną choroby zwanej moczówką prostą (diabetes insipidus), która charakteryzuje się wzmożonym pragnieniem i wydalaniem dużej ilości moczu (do 121 na dobę) o niskim ciężarze właściwym. Wyciągi z tylnego płata przysadki oraz syntetyczny ADH działają w tej chorobie leczniczo. Oksytocyna działa obkurczająco na ciężarną macicę. Obkurcza ona również pęcherzyki gruczołów m l e k o w y c h w okresie laktacji powodując wydzielanie mleka. Te właściwości oksytocyny wykorzystuje się w lecznictwie. '511
Mocz
Duzo
Normalnie
Mało
Ryc. 226. Produkcja i wydzielanie moczu w zależności od ilości wytwarzanego hormonu antydiuretycznego (ADH) w przysadce. A - Alkohol hamuje produkcję ADH działając moczopędnie, C's-t i praca fizyczna zwiększa wydzielanie ADH i hamuje diurezę.
GRUCZOŁ TARCZOWY Gruczoł tarczowy (glandula thyroidea), zwany pospolicie tarczycą, jest narządem nieparzystym, położonym w przedniej okolicy szyi przed tchawicą i krtanią. Dzięki temu położeniu jest on j e d y n y m gruczołem dokrewnym, którego powiększenie jest dostrzegalne z zewnątrz. Ciężar gruczołu wynosi 30-60 g. Składa się on z dwóch p ł a t ó w bocznych (lobi laterales) i z niepa rzystej części środkowej, zwanej węziną (isthmus). Czasami od węziny odchodzi ku górze tzw. płat p i r a m i d o w y (lobus pyramidalis). Węzina leży na tchawicy w obrębie II—IV pierścienia, płaty boczne d o l n y m brzegiem dochodzą do V pierścienia tchawicy, g ó r n y m - sięgają krtani. Powierzchnie gruczołu tarczowego pokrywa torebka łącznotkankowa, od '512
której odchodzą w głąb gruczołu wyrostki rozdzielające gruczoł na zraziki. Miąższ gruczołu zbudowany jest z pęcherzyków gruczołowych o średnicy 0,05-0,12 mm. Pęcherzyki te są wysłane od wewnątrz jedną warstwą komórek nabłonka sześciennego. Pęcherzyki mają różny kształt: mogą być okrągłe, owalne, wielokątne. Wnętrze ich wypełnia wydzielina komórek nabłonka, zwana koloidem. Tarczyca różni się od innych gruczołów dokrewnych tym, że funkcja jej w dużej mierze jest zależna od czynnika zewnętrznego, którym jest jod. Głównym hormonem wytwarzanym przez tarczycę jest tyroksyna. Ponadto powstają tu jeszcze w niewielkich ilościach dwu- i trójjodotyronina. Ta ostatnia wywiera działanie pięciokrotnie silniejsze niż tyroksyna. Dla prawidłowej czynności tarczycy konieczny jest dowóz jodu w ilości 35-50 mg na rok, czyli ok. 1 mg tygodniowo. Przeciętna dieta zawiera około 100-200 (i,g jodu dziennie. Jest to ilość wystarczająca na pokrycie zapotrzebowania tarczycy dla produkcji hormonu. Zawartość jodu w całym ustroju wynosi ok. 40 mg, z czego jedna piąta znajduje się w tarczycy. Przy braku jodu w wodzie i pożywieniu .tarczyca ulega powiększeniu, co uwidocznia się na zewnątrz w postaci tzw. wola. Wchłonięty z przewodu pokarmowego jod gromadzi się w tarczycy dzięki temu, że nabłonek pęcherzyków tarczycowych ma zdolność wychwytywania soli jodowych z krwi i około 25-krotnego zagęszczania ich w porównaniu ze stężeniem we krwi. Przy maksymalnej aktywności tarczycy zagęszczenie jodu w gruczole może sięgać 350-krotnej wartości jego stężenia we krwi. Równocześnie do światła pęcherzyków wydziela się swoiste białko zwane tyreoglobuliną. Odznacza się ono tym, iż jeden z aminokwasów wchodzących w jego skład - mianowicie tyrozyna, może reagować z jodem tworząc hormony tarczycy. Powstające w pęcherzykach hormony (głównie tyroksyna) są przechowywane w gruczole w postaci tzw. koloidu. Tyreoglobuliną w warunkach prawidłowych nie wydostaje się z gruczołu do krwi. Hormony tarczycy, tyroksyna i trójjodotyronina, aby wydostać się do krwiobiegu muszą zostać enzymatycznie odszczepione od tyreoglobuliny i dopiero w tej postaci dyfundują do naczyń krwionośnych. Aktywność enzymu odszczepiającego te hormony od tyreoglobuliny pozostaje pod kontrolą hormonu tyreotropowego przysadki. Działanie tyroksyny na ustrój polega przede wszystkim na zwiększaniu aktywności metabolicznej większości tkanek ustrojowych. Nadmiartyroksyny może zwiększyć podstawową przemianę materii od 60 do 100% ponad normę. Wzrost przemiany materii obciąża serce i układ krążenia, zwiększa produkcję ciepła; przyspieszone spalanie, prowadzi do utraty ciężaru ciała i sił przy zachowanym lub Wzmożonym łaknieniu. Tyroksyna wzmaga aktywność procesów psychicznych wywołując stan wzmożonego pobudzenia i niepokoju. Na poziomie komórkowym tyroksyna powoduje wzrost aktywności wielu enzymów, głównie w mitochondriach, co pociąga za sobą nasilenie procesów ogólnej przemiany komórki wyrażające się zwiększonym zużyciem tlenu, glukozy, tłuszczów i białek. 33 - Anatomia i fizjologia
513
A
B
C
Ryc. 227. Wygląd kobiety: A-z nadczynnością tarczycy, B - zdrowej, C- z niedoczynnością tarczycy.
Nadmierna produkcja tyroksyny prowadzi do powstania zespołu o b j a w ó w chorobowych charakterystycznych dla nadczynności tarczycy. Pierwotna postać nadczynności tarczycy nazywa się chorobą Gravesa-Basedowa. Głównymi jej objawami są: powiększenie tarczycy (wole), przyspieszenie czynności serca, wytrzeszcz gałek ocznych. Chorzy tacy są niespokojni, ruchliwi, pobudliw i , w y b u c h o w i , łatwo tracą panowanie nad sobą, często podejmują zbędną aktywność, łatwo się męczą. Niedoczynność tarczycy jest spowodowana niedoborem lub brakiem horm o n ó w tarczycy. Niedoczynność istniejąca od urodzenia charakteryzuje się zahamowaniem wzrostu (karłowatość) i niedorozwojem u m y s ł o w y m (matołectwo, kretynizm). Njedoczynność rozwijająca się u dorosłych powoduje powstanie tzw. obrzęku sluzakowatego. Następuje przy t y m zwolnienie wszystkich czynności ustroju, osłabienie, senność, spowolnienie m o w y , głos grubieje i staje się ochrypły, pojawia się obrzęk powiek i innych tkanek. Skóra staje się sucha, blada i chłodna. Występuje skłonność do wypadania włosów. Następuje zobojętnienie i przytępienie umysłowe. Wyżej przytoczone objawy nadczynności i niedoczynności tarczycy dają najlepsze pojęcie o jej roli fizjologicznej w ustroju.
GRUCZOŁY PRZYTARCZYCZNE Gruczoły przytarczyczne (glandulae parathyroideae) w liczbie dwóch par leżą na tylnej powierzchni tarczycy, a często są ukryte w jej utkaniu. Mają one kształt okrągławych lub owalnych nieco spłaszczonych t w o r ó w długości 6 - 8 m m , szerokości 2 - 4 m m . Mikroskopowo gruczoł składa się z pasm komórek nabłonkowych poprzegradzanych warstwami tkanki łącznej, która wnika do torebki pokrywającej gruczoł. '514
W odróżnieniu od innych gruczołów dokrewnych czynność gruczołów przytarczycznych nie jest regulowana przez przysadkę mózgową ani przez układ nerwowy. Hormon gruczołów przytarczycznych reguluje gospodarkę wapniowo-fosforanową w ustroju. Stężenie wapnia w osoczu wynosi ok. 10 mg/100 m ! , w a h a j ą c s i ę w w ą s k i c h granicach od 9 do 11 mg/100 ml. Poziom ten jest bardzo dokładnie regulowany - głównie przez hormon przytarczyc. Zwiększona aktywność gruczołów przytarczycznych powoduje szybką mobilizację soli w a p n i o w y c h z kości i wzrost poziomu wapnia w płynach ustrojowych (hiperkalcemia), zaś niedoczynność tych gruczołów pociąga za sobą spadek poziomu wapnia we krwi (hipokalcemia). Hormon przytarczyc zmniejsza wchłanianie zwrotne fosforanów w kanalikach nerkowych, wskutek czego następuje ucieczka fosforanów z moczem i w następstwie tego spadek ich stężenia w płynach ustrojowych. Odwrotnie - nawet bardzo niewielki spadek poziomu wapnia we krwi powoduje wzrost wydzielania hormonu przytarczyc, a jeśli trwa dłużej, jak np. w krzywicy, doprowadza do przerostu gruczołów. Przytarczyce powiększają się także podczas ciąży i laktacji. Z drugiej strony wszelkie sytuacje, w których dochodzi do wzrostu poziomu wapnia w płynie pozakomórkowym, jak np. przy nadmiernej podaży wapnia w diecie, przy zwiększonym dowozie witaminy D lub zwiększonej mobilizacji wapnia z kości z innych przyczyn gruczoły przytarczyczne ulegają zmniejszeniu. Niedoczynność przytarczyc prowadzi do spadku poziomu wapnia we krwi. Usunięcie gruczołów przytarczycznych powoduje po 3 - 4 dniach wystąpienie tonicznych nieskoordynowanych uogólnionych skurczów mięśniowych zwanych tężyczką. Objawy te są związane ze zbyt niskim stężeniem wapnia we krwi. Dożylne wstrzyknięcie soli wapnia przerywa napad tężyczki. Nadmierne wydzielanie lub wstrzyknięcie zbyt dużej ilości h o r m o n u powoduje niebezpiecznie duży wzrost zawartości wapnia we krwi. Nadczynność przytarczyc wiąże się ze wzrostem poziomu wapnia, który może dochodzić do 15-20 mg/100 ml. Następstwem tak wysokich wartości wapnia w płynie pozakomórkowym jest depresja ośrodkowego i obwodowego układu nerwowego, osłabienie mięśniowe, zaparcie stolca, bóle brzucha, powstawanie w r z o d ó w trawiennych, zaburzenia czynności serca. Nadmiar wapnia uruchamianego z kości w nadczynności przytarczyc jest wydalany przez nerki, co sprzyja powstawaniu kamicy nerkowej. GRASICA Grasica (glartdula thymus) jest stosunkowo dużym narządem, położonym w klatce piersiowej za mostkiem w śródpiersiu przednim górnym. U dzieci waży ona 10-18 g, u młodocianych 25-30 g. U osób dorosłych znajduje się w stanie zaniku. '515
Mikroskopowo grasica zbudowana jest ze zrazików porozdzielanych tkanką łączną. Każdy zrazik składa się z warstwy korowej i rdzeniowej. W warstwie korowej występują liczne komórki okrągłe, bardzo podobne do limfocytów. Hormon grasicy nie został dotąd odkryty i niektórzy sądzą, że grasica w ogóle nie jest gruczołem dokrewnym. Jedyną znaną czynnością grasicy jest produkcja limfocytów. GRUCZOŁY NADNERCZOWE Gruczoły nadnerczowe albo nadnercza ( g l a n d u l a e suprarenales) są to dwa niewielkie, płaskie twory, położone na górnych biegunach każdej nerki, otoczone cienką torebką łącznotkankową, która oddziela je od nerek. Prawe nadnercze ma kształt trójkątny, lewe - półksiężycowaty. Nadnercza są obficie ukrwione: w ciągu minuty przepływa przez nie pięć razy więcej krwi niż w y n o s i ich własna objętość. Na przekroju poprzecznym widać, że tkanka nadnerczy składa się z części zewnętrznej o zabarwieniu żółtawym, zwanej korą, i części wewnętrznej barwy ciemnobrunatnej - zwanej rdzeniem nadnerczy. Obie te części różnią się pomiędzy sobą pod względem pochodzenia, b u d o w y i czynności. Kora nadnerczy rozwija się z mezodermy, rdzeń pochodzi z tego samego źródła co komórki układu współczulnego. W badaniu mikroskopowym kora składa się z pasm komórek nabłonkowych, w których rozróżniamy trzy warstwy, zależnie od ułożenia tych pasm. Rdzeń jest złożony z komórek układających się w pasma lub wysepki w o k ó ł szerokich zatok żylnych, które drążą środkową część narządu. Komórki rdzenia nadnerczy barwią się intensywnie solami c h r o m o w y m i i dlatego noszą nazwę komórek chromochłonnych. j Nadnercza należą do narządów, bez których życie ustroju jest niemożliwe. Obustronne usunięcie gruczołów prowadzi nieuchronnie do śmierci w ciągu kilku dni. Zniszczenie samego rdzenia nie jest groźne dla życia. Stąd wniosek, że dla życia człowieka szczególne znaczenie ma kora nadnerczy. Część rdzeniowa nadnerczy jest w istocie dokrewnym odpowiednikiem układu nerwowego współczulnego - wytwarzane są tu i wydzielane do krwi dwa hormony: adrenalina i noradrenalina. Wywierają one na narządy wewnętrzne taki sam w p ł y w , jak pobudzenie n e r w ó w współczulnych, gdyż, jak już wspominaliśmy, noradrenalina jest mediatorem powstającym w zakończeniach włókien współczulnych. Różnica w budowie chemicznej adrenaliny i noradrenaliny jest niewielka (adrenalina jest metylowaną noradrenaliną). Obydwie substancje działają na o b w o d o w e naczynia krwionośne, podnosząc ciśnienie krwi; różnica w ich działaniu polega na t y m , że adrenalina działa silniej na serce, wzmagając jego czynność oraz zwiększa przemianę materii. Adrenalina i noradrenalina są stale wydzielane do krwi w niewielkiej ilości; wszelkie stany emocjonalne, zwłaszcza takie, jak strach lub gniew, powodują nagłe wyrzucenie większej ilości tych h o r m o n ó w , wywołując przyspieszenie czynności serca, zblednięcie skóry, skurcz mięśni przywłosowych. Jest to '516
reakcja przystosowująca ustrój do niebezpiecznej sytuacji, na którą prymitywną odpowiedzią jest ucieczka lub walka. Adrenalina zwiększa dopływ krwi do mięśni, przyspieszając czynność serca i zwiększając jego pojemność wyrzutową wskutek przesunięcia krwi z obszaru skórnego do jelitowego. Równocześnie adrenalina powoduje uwalnianie glukozy z glikogenu w wątrobie, dostarczając mięśniom dodatkowego źródła energii. Wyrazem tego jest wzrost stężenia cukru we krwi. W ten sposób organizm w obliczu grożącego niebezpieczeństwa mobilizuje wszystkie swe siły dla wykonania nadzwyczajnego wysiłku fizycznego. Dzięki adrenalinie człowiek w obliczu niebezpieczeństwa jest zdolny do „nadludzkieg o " wysiłku. Również wszystkie objawy tzw. tremy przed egzaminem, publicznym wystąpieniem itp. są związane z działaniem adrenaliny. Istota rdzeniowa nadnerczy przygotowuje zatem ustrój do maksymalnego krótkotrwałego ofensywnego wysiłku, natomiast kora nadnerczy, jak zobaczymy, mobilizuje Kora nadnercza
Zatrzymanie !>Oil
Ryc. 228. Hormony kory nadnerczy i ich działanie.
siły obronne ustroju o działaniu przedłużonym w odpowiedzi na różne, szkodliwe wpływy. Pod względem fizjologicznym kora nadnerczy jest najbardziej skomplikowanym gruczołem dokrewnym. Wyizolowano z niej i scharakteryzowano ponad 30 substancji sterydowych. Większość tych substancji stanowi etapy pośrednie na drodze syntezy kilku ostatecznych hormonów ze związku wyjściowego, którym jest cholesterol. Do głównych aktywnych sterydów kory nadnerczy należą hydrokortyzon, kortykosteron i aldosteron. Stosunkowo niewielkie różnice w ich budowie chemicznej powodują jednak zdecydowane różnice '517
w ich właściwościach fizjologicznych. Ujmując sprawę najogólniej, można przyjąć, że hydrokortyzon wywiera w p ł y w na przemianę węglowodanową i chroni ustrój przed stresem, aldosteron - reguluje gospodarkę elektrolitową, zaś kortykosteron zajmuje miejsce pośrednie wykazując cechy obu tych hormonów. Pojęcie stresu zostało wprowadzone przez fizjologa kanadyjskiego-Selyego - i chociaż zyskało sobie prawo obywatelstwa w fizjologii, medycynie i weszło niemal do wszystkich języków, podanie dokładnej definicji stresu nie jest łatwe. Dla rozważań związanych z fizjologią kory nadnerczy stres możemy określić jako zmiany w ustroju zachodzące pod wpływem czynników szkodliwych, powodujące zaburzenie jego fizjologicznej i chemicznej równowagi. Stresem są np. zmiany zachodzące pod wpływem napięcia nerwowego, zakażenia, urazu, promieni Roentgena itp. Zauważono, że stres wywołuje w ustroju szereg zmian identycznych z tymi, jakie występują po wstrzyknięciu hydrokortyzonu bez udziału czynników stresowych, a więc następuje wzmożony rozpad białka i produkcja glukozy z aminokwasów bez naruszania wątrobowych zapasów glikogenu, zatrzymywanie sodu i wzrost wydzielania potasu. Mamy tu zjawiska analogiczne do wpływu adrenaliny, która przygotowuje ustrój do nagłego wysiłku mobilizując z wątroby szybko dostępny materiał energetyczny - glukozę. Zapasy glikogenu w wątrobie są jednak niewielkiej w następnym etapie włącza się kora nadnerczy dostarczając energii z materiału białkowego. Hydrokortyzon hamuje syntezę białek i kieruje na drogę rozpadu dla produkcji energii zapasy białkowe pochodzące głównie z mięśni. Podawanie hydrokortyzonu u dzieci hamuje wzrost. Wszelki stres powoduje wzmożone wydzielanie hydrokortyzonu. Stres o bardzp ckiżej sile lub długim czasie trwania (jak np. uraz, krwawienia, wyczerpujące choroby, względnie nadmierny wysiłek fizyczny) może doprowądzić do wyczerpania kory nadnerczy. Organizm pozbawiony nadnerczy ginie pod wpływem nawet niewielkiego stresu. Hydrokortyzon zapobiega temu bardzo skutecznie i podawanie tego hormonu w takich sytuacjach niejednokrotnie ratuje życie. Aldosteron jest najpotężniejszym mineralokortykoidem nadnerczy. Działa on na nerki powodując zatrzymanie sodu w ustroju, z równoczesnym wydalaniem potasu. Podobne działanie ma kortykosteron, który jest znacznie słabszym mineralokortykoidem, jednakże jest wytwarzany w znacznie większych ilościach. W warunkach fizjologicznych w p ł y w tych hormonów na wydalanie potasu jest niewielki; wycięcie nadnerczy lub zniszczenie ich przez proces chorobowy (choroba Addisona) prowadzi do zatrzymywania potasu i strat sodu. Nadmiar mineralokortykoidów prowadzi nie tylko do zatrzymania sodu, lecz również do bardziej niebezpiecznych strat potasu. Oprócz wymienionych trzech głównych hormonów w korze nadnerczy występuje szereg substancji sterydowych będących członami pośrednimi na drodze syntezy poszczególnych hormonów. Niektóre z nich, tzw. androgeny, mają właściwości hormonów płciowych męskich. Syntezy sterydów nadner'518
czowych dokonuje układ enzymów i od sprawności tych enzymów zależy jakość ostatecznego produktu. Znane są dziedzicznie przekazywane zaburzenia w układzie tych enzymów, w wyniku których nadnercza produkują „nieprawidłowy zestaw" hormonów odpowiedzialny za krępą budowę, mocno rozwinięty układ mięśniowy i nadmierne owłosienie kończyn u młodych dziewcząt. Sekrecja hormonów kory nadnerczy pozostaje pod kontrolą przedniego płata przysadki, który wytwarza hormon adrenokortykotropowy, zwany w skrócie ACTH. Pierwotną odpowiedzią na stres jest wzmożona produkcja ACTH, która zwiększa z kolei produkcję hydro ko rtyzonu. Zależność pomiędzy ACTH i hydrokortyzonem ma charakter sprzężenia zwrotnego - z chwilą, gdy stężenie hydro ko rtyzonu we krwi wzrasta, przysadka przestaje produkować ACTH. Dlatego podawanie hydrokortyzonu pacjentom powoduje zahamowanie produkcji ACTH, co pociąga za sobą również zahamowanie czynności k^ry nadnerczy. Wydzielanie ACTH pozostaje pod kontrolą podwzgórza, które jest głównym ośrodkiem czuciowym odbierającym bodźce nerwowe i chemiczne nadchodzące z obszaru uszkodzonego w ustroju. Aldosteron nie ma właściwości antystresowych. Jego wydzielanie jest niezależne od ACTH. Istnieją choroby polegające na pierwotnej lub wtórnej nadczynności kory nadnerczy. Jest to tzw. zespół Cushinga, występujący wskutek nadmiernego wydzielania ACTH z przysadki (guzy) lub w następstwie przerostu lub guzów kory nadnerczy. Hormony sterydowe nadnerczy, przede wszystkim kortyzon i hydrokortyzon, okazały się bardzo skutecznymi środkami w leczeniu szeregu chorób, głównie ostrej choroby gośćcowej, ciężkich stanów alergicznych i kolagenoz. W związku z tym chemicy opracowali i wypróbowali szereg związków sterydowych o budowie zbliżonej do znanych hormonów. Uzyskano w ten sposób wiele cennych substancji o spotęgowanym działaniu leczniczym i zmniejszonym niepożądanym działaniu ubocznym, jak prednison, prednisolon, milikorten i in.
APARAT WYSEPKOWY TRZUSTKI Aparat wysepkowy trzustki składa się z tzw. wysp trzustki (Langerhansa). Są to okrągłe skupienia komórek nabłonkowych rozrzucone nieregularnie w obrębie tkanki trzustkowej. Liczba i wielkość wysp jest zmienna: średnica ich waha się od 0,1 do 0,5 mm. Komórki wysp są ułożone w pasma pozostające w ścisłym związku z naczyniami krwionośnymi. W wyspach trzustki rozróżniamy trzy rodzaje komórek w zależności od sposobu barwienia się różnymi barwnikami oznaczone greckimi literami a, (3 i y. Komórki (3 wytwarzają insulinę, która jest najważniejszym hormonem regulującym przemianę węglowodanową. '519
Insulina została odkryta w r. 1921 przez Bantinga i Besta. Było to epokowe odkrycie w historii medycyny, gdyż otrzymanie tego h o r m o n u umożliwiło leczenie cukrzycy - powszechnej choroby znanej od czasów starożytnych, która w dobie przedinsulinowej miała bardzo dużą śmiertelność. Wtrzydzieści siedem lat później, w 1958 r. w wyniku dziesięcioletniej żmudnej pracy, cherfiik Komórka
-Naczynie krwionośne
Ziarnistości zawierające hormon
Ryc. 229. Schemat ilustrujący proces wydzielania insuliny z komórki.
angielski Sanger ustalił budowę i wzór chemiczny insuliny, która jest białkiem o niewielkiej cząsteczce. Było to równocześnie pierwsze ustalenie składu cząsteczki białkowej, co stanowi pierwszy krok na drodze do jej syntezy. Na razie insulinę otrzymuje się z wyciągów trzustkowych bydła. Odkrycie insuliny otworzyło nowy rozdział w fizjologii, gdyż odgrywa ona olbrzymią rolę w procesach przemiany, przede wszystkim węglowodanowej. Węglowodany są głównym źródłem energii w ustroju potrzebnym dla podtrzymania przejawów życiowych. Czynność mięśni, mózgu i szeregu innych narządów wymaga stałego dowozu glukozy, której dosta rczaim krew zawierająca ten cukier w stałym stężeniu, wynoszącym ok. 100 mg/100 ml. Spadek poziomu cukru we krwi nazywamy hipoglikemią. Najbardziej czuły na hipoglikemię jest mózg. W razie braku węglowodanów w pokarmie ustrój potrafi '520
wytwarzać glukozę z białka. Nadmiar spożywanych węglowodanów ulega przemianie na tłuszcz i w tej postaci jest on magazynowany głównie w tkance podskórnej. Wszystkie te bardzo skomplikowane procesy są regulowane przez hormony, wśród których naczelną rolę odgrywa insulina. Pierwotna rola insuliny polega na tym, że umożliwia ona wejście glukozy do wnętrza komórki, gdzie cukier ten może ulec dalszym procesom: spaleniu.
Zapas glukozy we krwi
Zapas glukozy we krwi
Huszcze
Białka
Cykl kwasów trójkarboksylowych
Glikogen
Tłuszcze
Białka
trójkarboksylowych
Energia
Ryc. 230. Schemat ilustrujący przebieg procesów metabolicznych dostarczających energii ustrojowi. A - U osoby zdrowej (w obecności insuliny) i B - u chorego na cukrzycą (przy braku lub niedoborze insuliny). W tym drugim przypadku komórka (przedstawiona w postaci turbiny) czerpie energię ze źródeł zapasowych, głównie tłuszczu (wg Ganonga).
przemianie na glicerol lub tłuszcze. Brak insuliny lub nadmiar hormonów działających antagonistycznie utrudnia lub uniemożliwia wprowadzenie glukozy do komórki i co za tym idzie - wykorzystanie jej przez ustrój. Wchłaniany cukier gromadzi się wówczas we krwi, gdzie stężenie jego wzrasta nieraz do bardzo wysokich wartości. Wzrost stężenia cukru we krwi powyżej 120 mg% nazywamy hiperglikemią. Jeżeli stężenie cukru we krwi przekroczy 160-180 mg%, zaczyna on pojawiać się w moczu. Zjawisko to określa się mianem progu nerkowego, a wartość 160-180 mg% cukru we krwi jest wartością progową. Przechodzenie cukru do moczu nazywamy cukromoczem lub glikozurią. Wzrost stężenia cukru we krwi zwiększa ciśnienie osmotyczne krwi, którego następstwem jest zwiększone wydalanie moczu i zwiększone pragnienie. Przedstawione w wielkim skrócie '521
i uproszczeniu zaburzenia - wzrost poziomu cukru we krwi ( h y p e r g l i c a e m i a ) , cukromocz ( g l u k o s u r i a ) , wielomocz (polyuria) i wzmożone pragnienie (polydipsia) - są najistotniejszymi objawami cukrzycy i stanowią następstwo bezwzględnego lub względnego niedoboru insuliny. W cukrzycy organizm p o m i m o nadmiaru glukozy we krwi nie może jej wykorzystać i musi czerpać energię z innego materiału, głównie z tłuszczu. Uruchamianie i spalanie tłuszczu prowadzi do bardzo groźnych powikłań cukrzycy w postaci tzw. kwasicy cukrzycowej i śpiączki. W epoce przedinsulinowej powikłania te nieuchronnie prowadziły do zgonu. Odkrycie i wprowadzenie do lecznictwa insuliny zmieniło radykalnie rokowanie w tej bardzo rozpowszechnionej chorobie. Chorzy tacy muszą jednakże przez całe życie wstrzykiwać sobie insulinę; J e d n y m z problemów w leczeniu cukrzycy jest dobranie odpowiedniej dawki insuliny, odpowiadającej zmieniającym się potrzebom ustroju. Wprowadzenie do ustroju nadmiernej ilości insuliny powoduje duży spadek stężenia glukozy we krwi, czyli hipoglikemię. Jak już wspomnieliśmy, najbardziej czuły na hipoglikemię jest ośrodkowy układ nerwowy. Gdy poziom cukru we krwi spadnie poniżej 50-30 mg/100 ml,
Ryc. 231. Efekt obniżania się poziomu cukru we krwi (hipoglikemii).
'522
pojawiają się zaburzenia czynności ośrodkowego układu nerwowego szybko prowadzące do utraty przytomności (śpiączka hipoglikemiczna), drgawek i zgonu. Objawy hipoglikemiczne spotyka się najczęściej wskutek przedawkowania insuliny. Istnieją jednakże rzadkie choroby, w których dochodzi do samoistnego wzrostu wytwarzania insuliny w wyspach trzustki. Objawy w tych chorobach są takie same, jak przy przedawkowaniu insuliny. Insulina jest j e d y n y m h o r m o n e m obniżającym poziom cukru we krwi. Do h o r m o n ó w wywierających w p ł y w regulacyjny na przemianę węglowodanową należą h o r m o n y sterydowe nadnerczy, h o r m o n wzrostowy przysadki i adrenalina. Działają one w pewnym sensie antagonistycznie w stosunku do insuliny, podnosząc poziom cukru we krwi. Cukrzyca może być wywołana nie tylko bezwzględnym niedoborem insuliny, lecz także nadmiernym wydzielaniem takich h o r m o n ó w , jak sterydy nadnerczy lub h o r m o n wzrostowy.
GRUCZOŁY PŁCIOWE JAKO GRUCZOŁY DOKREWNE
Jajnik Budowa i czynności jajnika zostały o m ó w i o n e na str. 298. Jak w i a d o m o , jajnik obok wytwarzania gamet żeńskich pełni również rolę gruczołu dokrewnego. Te dwie czynności pozostają ze sobą w ścisłym związku - aktywność hormonalna jajnika jest zależna od jego zdolności wytwarzania komórek jajowych. W obu jajnikach znajdują się zawiązki ok. 400 000 komórek jajowych; z tego zaledwie jedna na tysiąc dojrzewa, uzyskując potencjalną szansę zapłodnienia. Każde jajo rozwija się w tzw. pęcherzyku jajnikowym (pęcherzyku Graafa), który również wytwarza hormonyżeńskie. Hormony te wstrzyknięte kastrowanym samicom zwierzęcym wywołują ruję (po łacinie oestrus) i dlatego nazywane są estrogenami. Pod względem chemicznym h o r m o n y te są sterydami o budowie zbliżonej do budowy h o r m o n ó w sterydowych nadnerczy i jąder. W pęcherzyku powstają głównie trzy estrogeny - estradiol, estriol i estron. Największą aktywnością odznacza się estradiol. W miarę wzrostu komórek pęcherzykowych zwiększa się wydzielanie estrogenów, które powodują wzrost i rozpulchnienie błony śluzowej macicy. Z chwilą pęknięcia pęcherzyka jajnikowego wydzielanie estrogenów nagle zmniejsza się. Dalszą kontrolę nad wzrostem błony śluzowej przejmuje teraz ciałko żółte ( corpus luteum), które powstaje w miejsce pękniętego pęcherzyka jajnikowego. Ciałko żółte wytwarza hormon, zwany progesteronem. Również i progesteron jest sterydem. Pod w p ł y w e m progesteronu błona śluzowa macicy grubieje, zachodzi w niej szereg złożonych zmian, które przygotowują ją do ciąży, stąd nazwa tego hormonu (gestatio- ciąża). Rozwijające się ciałko żółte wydziela coraz to większą ilość progesteronu. Przetrwałe komórki pęcherzykowe podejmują produkcję estrogenów, jednakże już w mniejszej ilości. Progesteron nie działa na wyściółkę macicy, dopóki nie zajdą w niej zmiany wywołane przez estrogeny. Hormon ten jest jednak '523
niezbędny do utrzymania ciąży. Progesteron jest również prekursorem horm o n ó w sterydowych nadnerczy. Jeżeli jajo nie zostanie zapłodnione, to po upływie dwóch tygodni od pęknięcia pęcherzyka jajnikowego ciałko żółte zanika, ustaje produkcja progesteronu i estrogenów, a pozbawiona w p ł y w u tych h o r m o n ó w błona śluzowa macicy ulega rozpadowi i wydaleniu na zewnątrz w r a z * krwią, powodując k r w a w i e n i e miesiączkowe, czyli miesiączkę. W ten sposób kończy się nie uwieńczony powodzeniem miesięczny cykl intensywnych przygotowań macicy do przyjęcia zapłodnionego jaja. Natychmiast rozpoczyna się cyki rozwoju nowego pęcherzyka. Wprawdzie cykl miesiączkowy odbywa się na osi jajnik - macica, jednakże czynnikiem regulującym jego rytm jest podwzgórze i przysadka. Przysadka jest o g n i w e m pośrednim pomiędzy układem n e r w o w y m i dokrewnym, centralnie regulującym cykl miesiączkowy kobiety. Powszechnie w i a d o m o , że znaczniejsze w p ł y w y e m o c j o n a l n e - z m i a n a pracy, wyjazd itp., mogą w y w o ł a ć nagłe zmiany w cyklu miesiączkowym kobiety. Podczas pierwszej p o ł o w y cyklu g ł ó w n y m składnikiem gonadotropin wytwarzanym przez przedni płat przysadki jest folikulostymulina; w czasie owulacji wzrasta produkcja hormonu luteinizującego i spada wytwarzanie folikulostymuliny. Współdziałanie dokrewne przysadki mózgowej i jajnika oparte jest na zasadzie tzw. sprzężenia zwrotnego. Jest to pojęcie zapożyczone z cybernetyki, coraz częściej używane w fizjologii, gdyż wiele mechanizmów fizjologicznych w ustroju ż y w y m działa na tej właśnie zasadzie. Sprzężenie zwrotne przysadki z jajnikiem polega na t y m , że folikulostymulina pobudza wydzielanie estrogenów, natomiast wysokie stężenie estrogenów hamuje wydzielanie folikulostymuliny. W podobny sposób duże stężenie progesteronu hamuje produkcję h o r m o n u luteinizującego. Cały opisany cykl przemian miesiączkowych zostaje zatrzymany z chwilą, gdy jajo zostaje, zapłodnione. Przez pierwsze dwa tygodnie ciąży zapłodnione jajo może utrzymać się w macicy dzięki obecności progesteronu wytwarzanego w ciałku żółtym. Po t y m okresie, jak powiedzieliśmy wyżej, zmniejsza się wydzielanie hormonu luteinizującego przysadki, co zapowiada zanik ciałka żółtego i wystąpienie krwawienia miesiączkowego grożąc oderwaniem się zapłodnionego jaja. W t y m wypadku nie dochodzi jednak do krwawienia, gdyż po d w ó c h tygodniach warstwa komórek otaczających zarodek, z której wytworzy się łożysko, zaczyna produkować h o r m o n - gonadotropinę kosmówkową, o dzia1łaniu bardzo p o d o b n y m do h o r m o n u luteinizującego przysadki. Hormon ten podtrzymuje aktywność ciałka żółtego, które pod jego w p ł y w e m przekształca się teraz w tzw. ciałko żółte ciążowe. Łożysko produkuje tak dużą ilość tego hormonu, że przechodzi on w wielkiej ilości do moczu kobiety ciężarnej. Na stwierdzeniu obecności tego hormonu w moczu oparte są próby diagnostyczne wczesnego okresu ciąży. Gonadotropina łożyskowa podtrzymuje ciałko żółte ciążowe w ciągu pierw'524
szych kilku tygodni. Od trzeciego - c z w a r t e g o miesiąca ciąży łożysko produkuje dostateczną ilość progesteronu i estrogenów; wycięcie jajników u ciężarnej kobiety w t y m okresie nie powoduje już przerwania ciąży.
Jądro W odróżnieniu od skomplikowanej cykliczności hormonalnej jajników jądra produkują tylko jeden h o r m o n w mniej więcej stałej ilości od chwili osiągnięcia dojrzałości płciowej do późnej starości. Hormon wytwarzany w jądrach nosi nazwę testosteronu. W nazwie zawarte jest określenie jego pochodzenia (testes) i b u d o w y chemicznej (sterydowej). Niektóre h o r m o n y sterydowe kory nadnerczy o budowie i właściwościach podobnych do testosteronu noszą nazwę androgenów. Testosteron jest wytwarzany przez komórki śródmiąższowe jąder, które stanowią bardzo niewielką część jądra. Produkcja h o r m o n u pozostaje pod kontrolą h o r m o n u luteinizującego przysadki, który bywa również określany przydługą nazwą „ h o r m o n u pobudzającego komórki śródmiąższowe", a który jest identyczny z h o r m o n e m luteinizującym kobiety. Wzajemny stosunek pomiędzy przysadką a jądrem działa na zasadzie „sprzężenia zwrotnego". Mechanizm ten nie jest jednak tak czuły, j a k u kobiety. Należy zaznaczyć, że przysadka wytwarza dwa h o r m o n y gonadotropowe, pobudzające jądra; te zaś produkują tylko jeden rodzaj h o r m o n u - t e s t o s t e r o n . Zasadniczo produkcja testosteronu jest sprzężona z h o r m o n e m luteinizującym. Natomiast wydzielanie folikulostymuliny wywierające u mężczyzny w p ł y w na produkcję spermatozoidów jest hamowane tylko przez bardzo duże ilości testosteronu. Testosteron pobudza wzrost kości i mięśni w kierunku kształtów męskich. Powoduje również wzrost narządów płciowych męskich oraz rozwój w t ó r n y c h cech płciowych męskich. Jedną z charakterystycznych cech w y w o ł a n y c h przez testosteron jest zgrubienie głosu, wskutek wzrostu krtani i więzadeł głosowych. Dorośli kastraci mają cienki, dziecinny głos.
HORMONY LOKALNE Hormonami lokalnymi nazywamy substancje, które występują w tkankach w postaci fizjologicznie nieczynnej, lecz mogą się aktywować w pewnych okolicznościach i w y w o ł y w a ć głębokie zmiany w bezpośrednim sąsiedztwie. Do substancji takich należą: 1) acetylocholina, 2) histamina, 3) heparyna, 4) serotonina. Acetylocholina, poza rolą pośrednika w przekazywaniu pobudzenia nerwowego, jest niezbędna do powstania czynności rytmicznych, np. skurczów serca. 34 - Anatomia i fizjologia
525
Histamina i heparyna są wytwarzane przez komórki tuczne. Fizjologiczna rola histaminy jest nie znana. Jest ona odpowiedzialna za miejscowe i ogólne odczyny alergiczne. Serotonina występuje w błonie śluzowej przewodu pokarmowego, w mózgu i w płytkach krwi. Jej rola fizjologiczna jest nie znana. Serotonina wywiera duży wpływ na krążenie, oddychanie, mięśnie gładkie i układ nerwowy.